
硅锌互作对水稻幼苗镉吸收转运特性的影响
2018-12-07 08:41焦欣田薛卫杰赵艳玲张长波刘仲齐
农业环境科学学报 2018年11期
焦欣田,薛卫杰,赵艳玲,张长波,闫 雷,刘仲齐*
(1.东北农业大学资源与环境学院,哈尔滨 150030;2.农业农村部环境保护科研监测所农业环境污染修复研究中心,天津300191)
重金属镉(Cd)因其污染性强、污染范围广而受到广泛关注。Cd在植物体内积累到一定程度后会抑制光合作用和呼吸作用,甚至导致生物量下降或死亡[1]。水稻是Cd积累能力最强的粮食作物,土壤中的Cd可以通过根系吸收转运到叶片和穗轴中,大气沉降中的Cd也可以通过叶片进入植物体内,在籽粒灌浆期间转运到稻米中[2-3]。稻米中的Cd通过食物链进入人体,危害人体健康[4]。
锌(Zn)是许多蛋白质和酶的重要组成部分,对植物的生长发育起着至关重要的作用[5-6]。研究表明,Zn与Cd会竞争相同载体蛋白进行跨膜运输[7]。加Zn可使水稻、烟草、小麦对Cd的吸收减弱,说明Zn对植物吸收Cd可能存在拮抗作用[8]。施Zn能促进小麦的光合作用,使POD酶活性提高,质膜稳定性增强,还降低了小麦体内脯氨酸的含量,进而加强了小麦对Cd胁迫的抵御能力[9]。此外,Zn可以通过缓解DNA损伤、调控脯氨酸及多胺代谢来降低Cd对植物的毒害[10]。土壤中Zn和Cd之间也存在交互作用。土壤中添加Cd会使Zn的吸附量下降、吸附速率减慢[11]。Zn能与吸附在土壤胶体上的Zn进行离子交换,降低土壤中Zn的有效性,增加土壤中可溶性Zn的含量。随着土壤中Zn浓度的增加,Zn与Cd在配合物上发生显著的竞争吸附,使得土壤中有效态Cd显著增加[12]。
硅(Si)是土壤中第二大元素,植物可以在茎叶中积累丰富的Si。Si能提高植物生长发育过程中抵抗各种生物和非生物胁迫的能力[13]。水稻是一种高积累Si的植物[14]。Si在质外体与Cd形成沉淀是缓解植物体内重金属毒害的主要原因[15]。Si和Cd在一些代谢不活跃的组织尤其是中柱鞘、木质部和韧皮部细胞壁内皮层形成的Si-Cd沉淀也可以抑制Cd的吸收和转运[16]。Si还能将更多的Cd分隔在液泡中或固定在节间[17-18],从而减少Cd在水稻中的移动。这些结果表明,施加Si能通过影响Cd在植物体内的吸收转运来减缓Cd的毒害作用。
虽然一定浓度的Si和Zn都能抑制水稻对Cd的吸收转运,但未见Si-Zn互作方式与水稻Cd吸收转运关系的研究报道。本文以水稻幼苗为材料,通过在根际环境中施加不同浓度的巯基硅和Zn,对水稻幼苗Cd的亚细胞分布特征、必需元素、可溶性蛋白进行研究,探讨了根际环境中添加Si和Zn离子抑制水稻Cd吸收转运的互作机理,从而为降Cd复合肥料的研发提供参考依据。
1 材料与方法
以湘早籼24(X24)为供试材料,选用饱满均一的水稻种子,在300 mL 5%的NaClO中浸泡30 min,用去离子水反复冲洗以去除残余的NaClO。水稻种子冲洗干净后均匀放入装有少量水的育苗盘内,放入28℃恒温培养箱黑暗培养72 h后移入人工气候室。待水稻长到两叶一心时,选取均一的水稻幼苗移到装有8 L 1/10Hoagland营养液的塑料盆(35 cm×25 cm×12 cm)中进行培养。塑料盆上盖有带小圆孔的硬塑料板,将幼苗用海绵固定在小圆孔内。待水稻长到三叶一心时挑选长势一致的幼苗,在去离子水中饥饿1 d,然后放置于以1/10 Hoagland营养液为基础的不同处理液中处理7 d。水培试验整个过程在人工气候室中进行,昼夜时间为16 h/8 h,昼夜温度为25℃/20 ℃,白天光照为 105 μmol·m-2·s-1、相对湿度为60%[19]。
1.1 培养与处理方法
设置处理液镉(CdCl2)浓度2.7 μmol·L-1,巯基硅浓度为0.05(Si1)、0.1(Si2)mmol·L-1,Zn浓度(ZnSO4·7H2O)分别为0、0.2 mmol·L-1。巯基硅来自深圳百乐宝生物农业科技有限公司,硅含量≥30%,溶解度为75%,明显优于硅酸钠(NaSiO3·H2O)的溶解度36.1%。处理液用HCl和NaOH调节pH至5.5~6。每个处理3次重复。
1.2 样品采集与处理
将处理好的水稻幼苗根系用5 mmol·L-1的CaCl2浸泡10 min[20],以吸附去除根系表面的Cd2+,之后用去离子水冲洗干净,用吸水纸吸干表面水分,用剪刀将地上部与根系分开。一部分放入信封,75℃恒温烘干,用于全Cd测定;另一部分,分别称取根系和地上部鲜样1 g,用锡纸包裹后放入液氮中冷冻,之后置于-80℃储存,用于亚细胞测定。
1.3 Cd、Si含量的测定
用剪刀将恒重的样品剪碎,称重后置于消煮管中,加入7 mL HNO3过夜,用消解仪ED54于110℃加热2.5 h,冷却后加入1 mL的H2O2,盖盖加热1.5 h,170℃开盖赶酸至0.5 mL左右,定容至25 mL,用原子吸收仪(AAS,ZEEnit70d0,Analytikjena,德国)测定样品中Cd含量[21]。
分别吸取 Si标液 0、0.125、0.25、0.5、1.25、2.5、5 mL 定容至 50 mL,Si含量分别为 0、2.5、5.0、10、25、40、50 μg·mL-1。向10 mL离心管中依次加入2.7 mL去离子水、200 μL Si标液、1.5 mL 0.26 mol·L-1HCl、200 μL 10%钼酸铵,静止5 min后再加入200 μL 20%酒石酸、200 μL 2%抗坏血酸摇匀静止25 min。样品采用高压灭菌法处理后依次加入药品,600 nm处比色[22]。
1.4 亚细胞组分的测定
将冷冻的水稻幼苗放在含有50 mol·L-1Hanks平衡盐溶液(HEPES)、500 mol·L-1蔗糖溶液、1 mol·L-1二流苏糖醇(DDT)、5 mmol·L-1抗坏血酸和1%(W/V)聚乙烯吡咯烷酮(PVPP)的提取缓冲液中匀浆,并用NaOH调pH至7.5。匀浆液通过尼龙布(100目)过滤,用缓冲液冲洗后100 g离心5 min,沉淀为细胞壁组分(F1)。上清液以10 000 g离心30 min,沉淀为细胞器组分(F2),上清液为胞液组分(F3)。所有步骤均在4℃下进行[23]。各组分烘干后按照1.3的步骤测定Cd含量。
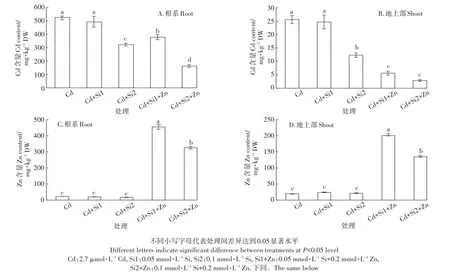
图1 Si、Zn处理对Cd胁迫中根系和地上部Cd及Zn含量的影响Figure1 Effects of Si and Zn treatments on the concentrations of root Cd,shoot Cd,root Zn and shoot Zn under Cd stress
1.5 可溶性蛋白的测定
称取45.6 g K2HPO4·3H2O和27.2 g KH2PO4放入1 L容量瓶中摇匀,标为A液和B液。将153.8 mL A液和96.2 mL B液加去离子水定容至1 L,配制成pH为7的50 mmol·L-1磷酸缓冲液。将227 mL A液和23 mL B液用去离子水定容至1 L,配制成pH为7.8的0.05 mmol·L-1磷酸缓冲液。称取0.3 g EDTA-Na2和5 g聚乙烯吡咯-K30溶于1 L pH为7的磷酸缓冲液,配制成粗酶提取液。
将新鲜水稻幼苗用去离子水清洗干净,然后各称取0.5 g左右。用液氮研磨,加入8 mL提取液,4℃10 000 r·min-1离心20 min,取上清液待测。
1.6 数据处理与计算
元素转移因子(%)=地上部元素含量/根系元素含量×100%[24]
Cd在水稻根系和地上部亚细胞组分中的分配比率(%)为各器官亚细胞组分中的Cd含量占各器官Cd总量的百分比。
采用Excel 2007、SPSS 11.5进行数据统计分析和作图。
2 结果与分析
2.1 Si-Zn互作对水稻幼苗Cd和Zn含量的影响
由图1所示,施加Si和Zn对水稻根系和地上部Cd含量有显著影响。施加0.05 mmol·L-1和0.1 mmol·L-1巯基硅可使根系Cd积累量下降5.4%~34.6%(图1A)。地上部Cd积累量分别下降3.5%和51.8%(图1B)。同时添加0.2 mmol·L-1Zn和0.05 mmol·L-1Si或0.1 mmol·L-1Si使水稻幼苗根系中的Cd含量分别下降27.4%和68.9%,使地上部的Cd含量分别下降78.0%和88.0%。Si和Zn共存时对Cd的抑制效应明显大于单独施加巯基硅的抑制效应,对地上部Cd积累量的抑制效应大于对根系Cd的抑制效应。
单独添加巯基硅对水稻幼苗中的Zn含量没有显著影响。施加0.2 mmol·L-1Zn后,水稻幼苗根的Zn含量明显高于地上部,锌含量随着添加Zn浓度的增加而大幅升高。和对照相比,Si1+Zn和Si2+Zn处理使水稻根系中的Zn含量分别提高了19倍和14倍(图1C),使地上部中的Zn含量分别提高了10倍和6倍(图1D)。
2.2 Si-Zn互作对水稻幼苗亚细胞组分中Cd含量及分配比例的影响
水稻根系细胞壁、细胞器和胞液组分中的Cd含量有显著差异。Cd在幼苗根系的亚细胞分布表现为细胞壁>胞液>细胞器(图2A),在地上部的亚细胞分布表现分别为细胞壁≈胞液>>细胞器(图2B)。施加Si和Zn显著降低了水稻幼苗亚细胞各组分的Cd含量。其中细胞壁和胞液中的Cd含量下降幅度较大。
如图2A所示,和Cd处理相比,Cd+Si1处理对根系细胞壁组分中的Cd含量无显著影响,Cd+Si2处理使根系细胞壁组分中的Cd下降了43.8%,但这2种处理使胞液组分中的Cd含量分别下降了38.7%和53.0%。Zn-Si互作显著抑制Cd进入胞液组分的过程。Cd+Si1+Zn和Cd+Si2+Zn处理使细胞壁组分中的Cd含量分别下降了39.2%和79.8%,使胞液组分中的Cd含量分别下降了43.9%和63.7%。Zn-Si互作显著抑制Cd进入细胞壁和胞液组分的过程。如图2B所示,Cd+Si1和Cd+Si2处理使地上部细胞壁组分中的Cd含量分别降低了18.3%和56.4%,胞液组分中的Cd含量分别下降了14.8%和61.7%。Cd+Si1+Zn和Cd+Si2+Zn处理使细胞壁中的Cd分别下降了87.2%和91.4%,使胞液中的Cd含量分别下降了95.3%和95.4%。Si-Zn互作对根系细胞壁中Cd积累的抑制作用较弱,对胞液中Cd的积累抑制作用较强。
在根际环境中添加Si和Zn对根系和地上部亚细胞组分中的Cd分配也产生了显著影响。随着Si浓度的增加,根系细胞壁中的Cd分配比例显著下降,和无Si的Cd处理相比,Cd+Si2和Cd+Si2+Zn处理使根系细胞壁中的Cd分配比例分别下降了10%和20.3%(图3A)。而地上部Cd分配比例的下降主要表现在细胞液组分中,Cd+Si2、Cd+Si1+Zn和Cd+Si2+Zn处理使地上部细胞液中的Cd分配比例分别下降了24.4%、53.4%和51.9%(图3B)。

图2 Si、Zn处理对Cd胁迫下幼苗根系和地上部亚细胞中Cd含量的影响Figure2 Effects of Si and Zn supply on Cd concentrations in subcellular fraction of roots and shoots in rice seedlings under Cd stress
2.3 Si-Zn互作对水稻幼苗必需元素含量的影响
在 Cd 胁迫环境中,增加 0.05~0.1 mmol·L-1的巯基硅,根系中的Ca、Fe含量都显著下降,Mn的含量显著增加(表1)。和单施Si相比,同时添加Si和Zn时,根系中的K和Zn含量显著增加,而Ca和Mn含量则显著下降。Si-Zn共存时对水稻地上部的K、Fe和Zn含量有明显的促进作用。从转移因子来看,与单施Si相比,Si-Zn互作促进根系中Ca、Fe向地上部的转运,对K、Mn的转运有一定的抑制作用(表2)。
2.4 Si和Zn对水稻根系和地上部可溶性蛋白的影响
与单独的Cd胁迫处理相比,添加0.05 mmol·L-1和0.1 mmol·L-1的巯基硅使根系中的可溶性蛋白含量分别上升14%和44%,Si和Zn共存时,根系中的可溶性蛋白含量可提高20%~45%(图4A)。但地上部的可溶性蛋白含量在处理间没有显著差异(图4B)。
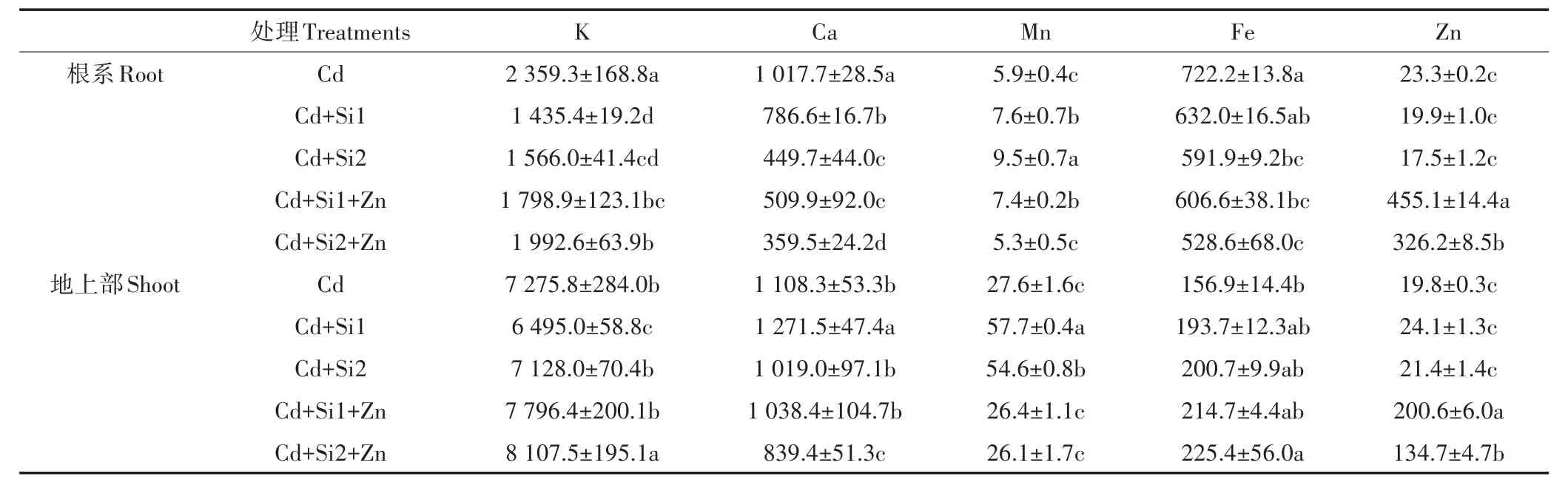
表1 Si和Zn对水稻幼苗根系和地上部必需元素含量的影响(mg·kg-1DW)Table1 Effects of Si and Zn on essential elements in rice seedlings under Cd stress(mg·kg-1DW)

图3 Si、Zn处理对幼苗根系和地上部亚细胞组分中Cd分配比例的影响Figure3 Effects of Si and Zn on Cd distribution in subcellular fractions of roots and shoots in rice seedlings under Cdstress
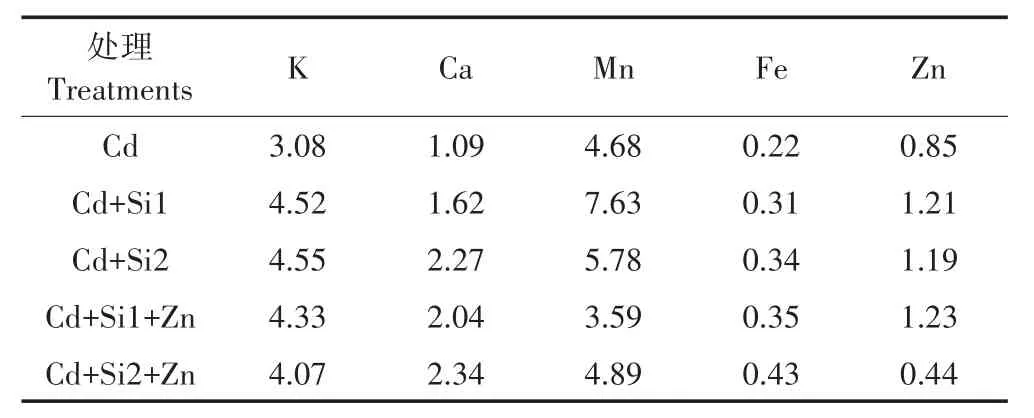
表2 Si和Zn对水稻幼苗必需元素转移因子的影响Table2 Effects of Si and Zn on the translocation factors of essential elements in rice seedlings under Cd stress
3 讨论
Si是促进水稻生长发育、提高水稻抗逆能力的有益元素,Zn则是水稻生长发育所必需的微量元素,参与多种生命活动过程。增加根际环境中的Si或Zn离子浓度能有效缓解Cd对根系的生理毒害,促进根系和地上部的生长发育[8,25-28]。本研究发现,在含2.7 μmol·L-1Cd的营养液中添加低浓度的Si(0.05 mmol·L-1)对水稻根系中的Cd积累量几乎没有影响,当巯基硅的浓度增加到0.1 mmol·L-1时,水稻根系和地上部的Cd含量才会显著下降。但当Si和0.2 mmol·L-1Zn共存时,低浓度的巯基硅也能显著抑制水稻对Cd的吸收和转运。
植物在长期的进化过程中形成了多元的解毒机制,在重金属污染环境中,水稻能将大量的Cd固定在根系中[29],只有少部分Cd通过维管束组织向地上部转运。Si既能促进细胞壁的发育以及Cd与细胞壁中半纤维素的结合[30],又能和Cd形成难移动的化合物沉淀在细胞壁或液泡中[31],还能通过降低蒸腾作用抑制Cd从根系向地上部的转运[32]。Zn是多种蛋白质的主要辅酶,对重金属的跨膜运输和进出韧皮部的过程都有重要调控作用[33-34]。在本试验中,Si和Zn共存时对地上部的K、Fe和Zn含量都有显著的促进作用,必需元素吸收转运效率的提高必然会抑制相关通道对Cd的转运,因而能使根系胞液组分中的Cd含量下降43.9%以上。随着根系中可移动的Cd数量大幅度减少,地上部的Cd总量也显著下降。Si和Zn互补作用使得低浓度的Si也能对Cd在水稻体内的转运产生显著的抑制效果。
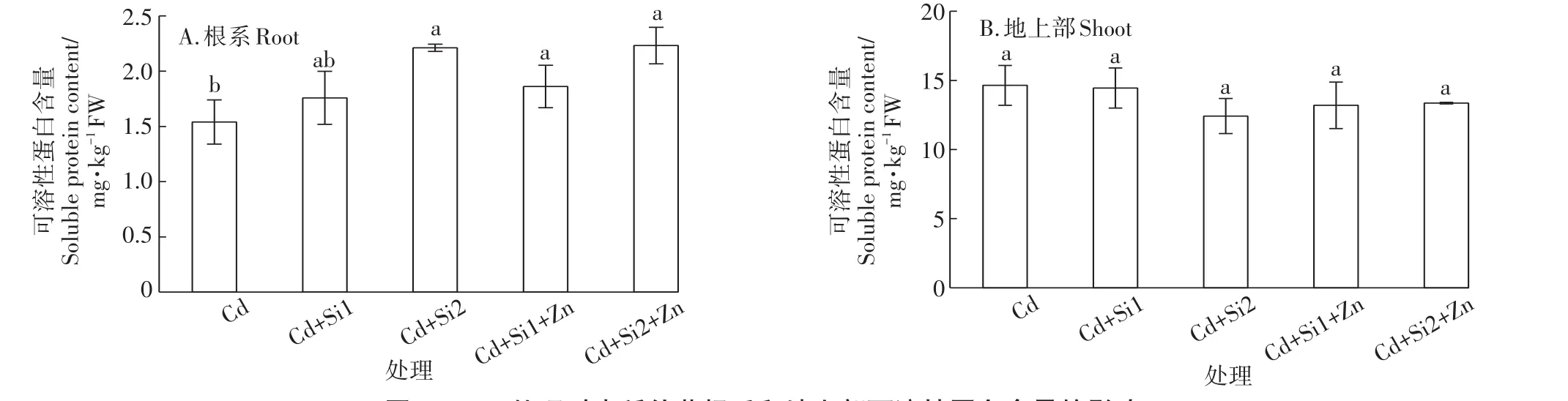
图4 Si、Zn处理对水稻幼苗根系和地上部可溶性蛋白含量的影响Figure4 Effects of Si and Zn on soluble protein of roots and shoots in rice seedlings under Cd stress
植物体内的可溶性蛋白质和可溶性糖作为渗透保护物质,有利于植物在逆境胁迫下维持细胞的结构和功能[35]。Cd胁迫通常会抑制可溶性蛋白的合成,降低酶的生物活性[11]。而Si和Zn对维持细胞内大量元素(如P、K、S)和微量元素(如Fe、Cu、Mn、Zn等)的离子平衡发挥着重要作用[36-38]。这种离子动态平衡的保持与可溶性蛋白的代谢有着密切的关系。环境中增加Si浓度不仅能显著提高Lsi1基因的表达水平,同时还调控OsHMA3、Nramp5等许多与离子转运、金属硫蛋白合成等有关的基因[30,32],因而对蛋白质代谢产生重要影响。当Si和Zn共存时,水稻幼苗根部的可溶性蛋白含量显著提升,说明Cd对根系的生理伤害因Si和Zn的存在而修复。由于地上部胞液组分中的Cd含量仅相当于根系胞液中的十分之一,对细胞产生的生理毒害甚微,所以地上部的可溶性蛋白含量在处理间没有表现出显著差异。
4 结论
(1)在含 Cd 2.7 μmol·L-1的营养液中添加 0.1 mmol·L-1的巯基硅能使水稻幼苗根系和地上部的Cd含量分别下降34.7%和51.8%。
(2)Si和Zn共存时对Cd的抑制作用更加明显,同时添加0.2 mmol·L-1Zn和0.05 mmol·L-1的巯基硅,能使水稻幼苗根系和地上部的Cd含量分别下降27.4%和78.0%。
(3)同时添加Si和Zn能促进根系对K、Ca、Fe、Zn等元素的吸收以及向地上部的转运,并能显著提高根系中可溶性蛋白的含量。
猜你喜欢
广州化学(2022年4期)2022-09-01
中国金属通报(2021年20期)2021-03-11
三农资讯半月报(2020年18期)2020-10-14
商品与质量(2019年31期)2019-11-28
天然产物研究与开发(2019年1期)2019-03-01
数码世界(2018年1期)2018-12-23
浙江工业大学学报(2017年5期)2018-01-22
饮食科学(2016年9期)2016-11-18
分析科学学报(2016年2期)2016-10-15
中南民族大学学报(自然科学版)(2015年2期)2015-12-16
