
青藏高原川西北高寒牧区黑麦青贮饲料中耐低温乳酸菌的分离与鉴定
2019-01-07 01:57李世丹陈仕勇
草原与草坪 2018年6期
李世丹,陈仕勇,马 莉,王 泰
(1.西南民族大学 生命科学与技术学院,四川 成都 610041; 2.四川省阿坝州畜牧科学技术研究所,四川 汶川 623000)
川西北高寒草地位于青藏高原东缘,草地畜牧业是该地区经济发展的支柱产业。但由于川西北高寒草地海拔高,气候寒冷,牧草生长期短,冷季长达8个月,一直以来天然草地牧草供给的季节性不平衡严重制约着草地畜牧业的高效发展[1]。因此,在青藏高原川西北牧区通过人工种草和天然草地打草调制青贮饲料,不仅是解决冬春季节饲草料严重不足的有效措施,同时也是提高畜牧业生产水平的有效手段[2-3]。
牧草青贮料比青干草较好的保持了牧草的营养价值,但受青藏高原气候条件和调制技术的制约,使青贮料在草地牧业生产中的应用远不及青干草广泛[4]。牧草青贮发酵进程最终应是以乳酸菌为优势的微生物群落演替过程,乳酸菌的种类及其生长特性影响着青贮饲料的发酵品质,因而通过对川西北高寒草地牧草中天然附着的乳酸菌进行分离和鉴定,筛选出适合当地气候的优良耐低温乳酸菌株,开发出乳酸菌青贮添加剂,对提升川西北高寒牧区青贮料调制技术和促进青贮料的生产应用具有重要意义[5]。
目前,青藏高原青贮乳酸菌的多样性与种质资源的研究已备受关注,其中包括从高原传统发酵乳品分离与鉴定乳酸菌的工作[6],对高寒地区野生牧草天然附着乳酸菌进行分离鉴定[7]。许冬梅等[8]从青藏高原地区垂穗披碱草、燕麦等分离出148株乳酸菌,进一步分析表明植物乳杆菌、干酪乳杆菌等4种乳酸菌可作为低温发酵青贮饲料的备选菌株。崔棹茗等[9]对青藏高原地区青稞秸秆青贮饲料中的乳酸菌进行了分离,结果共分离出植物乳杆菌、乳酸片球菌、戊糖片球菌等4种耐低温、生产快,产酸能力强的菌株。在青贮的制作过程中,不同区域及不同原料青贮中的乳酸菌种类存在差异。目前,对川西北高寒牧区青贮乳酸菌的研究较少,也没有成功研制出一种应用于生产的商品乳酸菌青贮添加剂[10-11]。因此,以川西北高寒牧区饲用黑麦青贮料为材料,从中分离、筛选适合当地青贮用添加剂的乳酸菌菌株,进一步鉴定乳酸菌种类,并对其生物学特性进行研究,为川西北高寒牧区青贮用乳酸菌添加剂的研制和应用提供理论依据。
1 材料和方法
1.1 供试材料
供试黑麦(Secalecereale)为课题组引种筛选的新品系M1306。供试材料2017年5月6日种植于四川省阿坝州红原刷金寺镇阿坝州畜牧科学技术研究所试验田。地理座标为N 32°01′、E 102°62′,海拔3 351 m,属大陆性高原寒温带季风气候,年均温1.4℃;年均日照时数2 158.7 h;年均降水量749.1 mm,无绝对无霜期。黑麦生长至初花期,于2017年7月26日刈割鲜草450 kg,晾晒萎蔫,切短至4~5 cm,分层装填并压实于长95 cm、宽58 cm的3个塑料袋中,用胶带密封,外套编织袋防护,存储在室内。青贮60 d后对3袋青贮料分别取样0.5 kg,混合真空包装,带回实验室进行乳酸菌的分离与鉴定。
1.2 试验方法
1.2.1 乳酸菌的分离和纯化 在无菌操作下取10 g待测青贮料样品,剪碎放入装有100 mL无菌生理盐水的三角瓶,用震荡仪器充分振荡,使样品附着的乳酸菌分散在液体中,静置后取上清液配置成7个梯度的乳酸菌稀释液。然后取各稀释液分别涂布在MRS平板培养基上,静置约30 min,倒置于20℃恒温培养箱中低温厌氧培养48 h,选择菌落密度适中、生长良好、溶钙圈明显的菌落进行革兰氏染色和过氧化氢酶实验。确定的乳酸菌菌落采用平板划线法再进行纯化培养,最后将分离纯化后的乳酸菌按1∶1比例保藏在30%的甘油中,-80℃低温冰箱保存。
1.2.2 乳酸菌菌株16S rDNA测定 按照Ezup柱式细菌基因组DNA抽提试剂盒(上海生工)产品使用说明提取乳酸菌全基因组DNA。基因片段扩增引物为27F(5′-AGAGTTTGATCCTG-3′),1492R(5′-TACCTTGTTACGACT-3′);PCR扩增及反应按常规方法进行[14],扩增产物送生工生物工程(上海)股份有限公司测序。
1.2.3 不同梯度温度生长试验 共设4、10、15、20、25、30、37、45、50℃ 9个温度梯度,每个梯度设3个重复。在无菌操作台内,用5 mL EP管装入MRS肉汤液体灭菌培养基,接种3%的活化后的菌液;然后把接种好的乳酸菌对应分组编号,置于温培养箱中厌氧培养,并保证除温度外其他培养条件相同。每个温度下各乳酸菌菌株培养3 d,目测不同乳酸菌在不同温度下的生长状态,采用4级记录符号(-、±、+、++)记录结果,乳酸菌在繁殖时,菌体主要沉淀在培养基底部,有的则呈絮状悬浮在培养基中。在观察时,培养基底部有明显乳酸菌集群沉淀,并且培养基内,一半以上充满菌体,呈絮状悬浮在试管中,则判定为乳酸菌在这个温度下生长极好(++);乳酸菌在培养基底部有沉淀,悬浮的絮状乳酸菌占培养基的一半,视为乳酸菌生长良好(+);在培养基底部有少量乳酸菌沉淀,或在培养基中有少量乳酸菌絮状悬浮,认为此时菌体较少(±);当培养基内基本观察不到乳酸菌沉淀,以及培养基内没有乳酸菌絮状悬浮,就判定此时培养基内无菌体(-)。
1.2.4 生长曲线绘制 各个温度下,不同种乳酸菌均培养3 d,用UV-1800型紫外分光光度计测各温度下不同乳酸菌菌液的D(600 nm)值并记录,绘制生长标准曲线,观察乳酸菌对温度的耐受性。
1.2.5 统计分析 采用EXCEL 2010软件进行数据统计和曲线图制作,测序序列采用CLUSTALW进行多重比较,并用MEGA 5.0软件构建系统发育树[15]。
2 结果与分析
2.1 乳酸菌菌株16S rDNA基因序列测定及系统进化树的构建
青贮样品经低温培养,分离纯化,结合革兰氏染色和过氧化氢酶试验,共分离出4株乳酸菌菌株(Y1、Y5、Y7、Y9)。经16S rDNA PCR扩增序列长度分别大于1 450 bp,在GENEBANK中进行16S rDNA基因同源性序列比对鉴定,与标准乳酸菌菌株序列相似性均达到了99%。选用4种乳酸菌参考菌株(表1)与分离的乳酸菌菌株构建系统进化树(图1)。经过同源比对及4株乳酸菌菌株聚类结果分析表明,4株菌株分别为乳杆菌属(Lactobacillus)和肠球菌属(Enterococcus),其中菌株Y1为清酒乳杆菌(L.sakei)、Y9为弯曲乳杆菌(L.curvatus)、Y5为植物乳杆菌(L.plantarum)、Y7为屎肠球菌(E.faecium)。
2.2 不同温度下培养基浑浊度及分析
通过目测不同温度下在MRS肉汤液体培养基培养3 d后的乳酸菌浑浊度,得到不同温度下MRS液体培养基的浑浊度目测结果(表2)。

表1 构建乳酸菌系统发育树的参考菌株Table 1 Referenced lactic acid bacteria used for construction phylogenetic tree
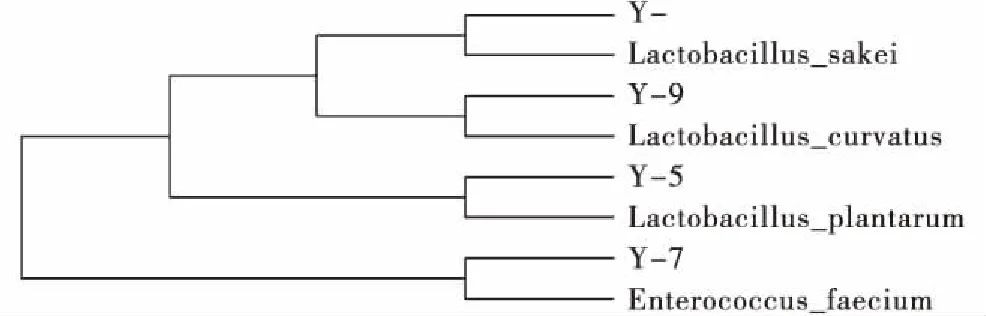
图1 基于16S rDNA基因序列建立的乳酸菌系统进化树Fig.1 Phylogenetic tree of 16S rRNA sequences
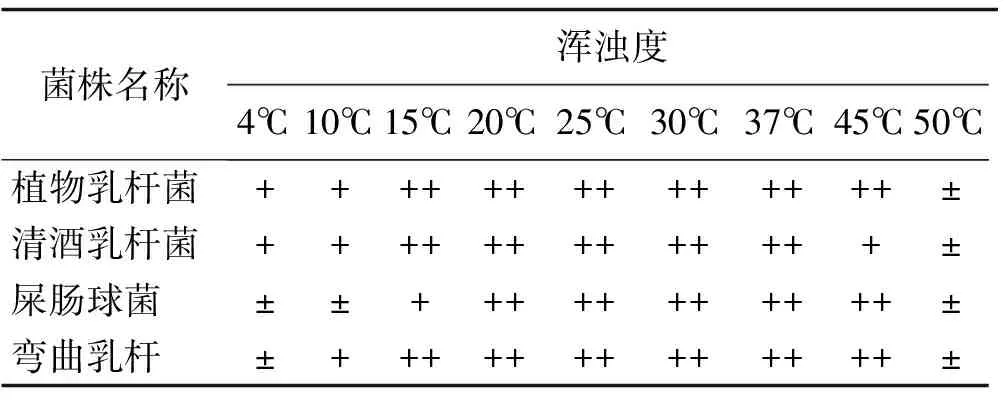
表2 不同温度下MRS液体培养基的浑浊度目测结果Table 2 Visual measurement results of turbidity of MRS liquid medium at different temperatures
注:++表示生长极好、+表示生长良好、±表示菌体很少、-表示无菌体
在不同温度培养3 d后,各个菌株生长情况各不相同,同一温度下不同菌株生长状况不同,同一菌种在不同温度下生长状况也不相同。4℃时有2株乳酸菌生长良好,其余2株乳酸菌能微弱生长;10℃时有3株乳酸菌生长良好,1株乳酸菌菌体较少;15℃时,3株乳酸菌生长极好,1株乳酸菌良好生长;20~37℃时所有菌株均生长极好;45℃时,3株乳酸菌生长极好,1株乳酸菌良好生长;50℃的条件下,所有菌株均生长微弱。试验表明,弯曲乳杆菌和植物乳杆菌生长极好的温度在15~45℃;清酒乳杆菌生长极好的温度在15~37℃;屎肠球菌生长极好的温度在20~45℃。
2.3 乳酸菌在不同温度处理下的生长趋势
乳酸菌经过72 h培养,不同温度下的培养基D(600 nm)值绘制的各菌株在不同温度下的生长曲线不同(图2)。温度在4~20℃时,4种乳酸菌随温度的升高生长量增大,其中植物乳杆菌、清酒乳杆菌和弯曲乳杆菌生长量较大,屎肠球菌生长量相对较少,20℃时菌体浓度达到较高水平;温度在20~37℃时,4种乳酸菌的生长量不再增加,均达到稳定,植物乳杆菌、清酒乳杆菌和弯曲乳杆菌菌体浓度稍有增加,屎肠球菌菌体浓度无显著变化;温度在37~50℃时,4种乳酸菌的生长量呈下降趋势,乳酸菌活性受到抑制,植物杆菌和弯曲乳杆菌活性最高,清酒乳杆菌活性最低,屎肠球菌稍高于清酒乳杆菌,菌体浓度不高。当温度升至50℃时,4种乳酸菌的活性均降低到非常低的水平。因此,4种乳酸菌最适生长温度基本稳定在20~37℃,在培养温度20℃以下低温范围,植物乳杆菌对低温的耐受性最好,屎肠球菌对低温耐受性最低,清酒乳杆菌和弯曲乳杆菌对低温的耐受性居中,4种乳酸菌均有较好的耐低温表现。
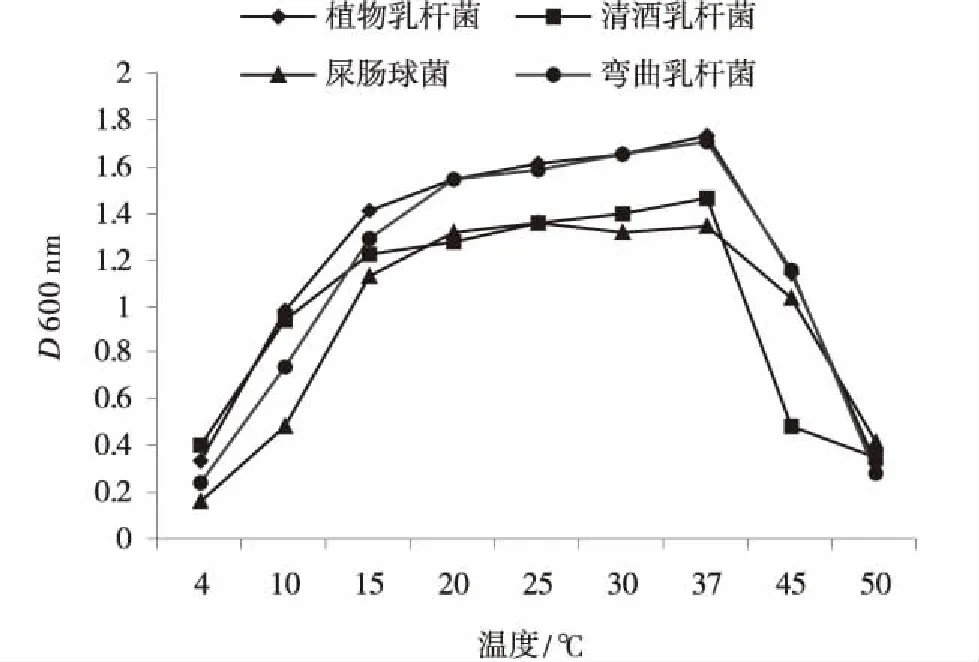
图2 各乳酸菌在不同温度下培养72 h生长状况Fig.2 Growth conditions of lactic acid bacteria in different temperature after 72 h culture
3 讨论
3.1 乳酸菌的鉴定
青贮饲料的乳酸菌多样性受青贮原料种类的影响,不同原料青贮乳酸菌分类鉴定报道主要有乳杆菌、片球菌、明串珠菌、肠球菌、乳球菌、链球菌以及魏斯特氏菌7个属[14-15]。试验分离出的4株乳酸菌用形态观察及生理生化鉴定方法不能把这些菌株鉴定到种的水平。16S rDNA同源序列分析方法常应用于微生物种和亚种的鉴定,当2个鉴定种间的16S rDNA序列同源性大于97.5%,可认为是同一种菌[16-17]。把提取的乳酸菌DNA进行16S rDNA序列扩增,测序结果与标准菌株对比,4株菌的16S rDNA 序列与参考菌株的16S rDNA 序列相似性都在99%,因此,可以认为鉴定结果准确。
3.1 温度对目测浑浊度的影响
目测MRS肉汤液体培养基浑浊度是判定乳酸菌生长情况最直观的方法,根据目测浑浊度等级确定植物乳杆菌、清酒乳杆菌、弯曲乳杆菌和屎肠球菌的最适生长温度均为20~37℃。低温会降低菌体酶活性,高温会破坏菌体酶蛋白结构,使得蛋白酶变性,从而让乳酸菌繁殖受到抑制,菌体浓度降低,所以使得目测浑浊度降低。在适宜乳酸菌生长的温度范围内,菌体酶活性高,繁殖速度快,则体现浑浊度大。同一菌株在不同温度下的繁殖性能不同,则菌液浑浊度观察结果不同。然而不同的菌种对温度的耐受性不同,所以表现出来的就是不同乳酸菌在同一温度下不同的浑浊度。以同类方法研究,尚天翠[18]报道的乳酸菌的最适生长温度在25~38℃;王夫杰等[19]报道的3株风味乳酸菌生长温度为32~37℃。
3.2 不同温度条件对乳酸菌生长的影响
由于乳酸菌对温度的适应性不同,则其在不同温度下的生长状况各异,在各自适应生长的温度下,繁殖性能良好,随着温度的改变,生长适应性减弱,繁殖受到抑制。利用20℃低温筛选出的乳酸菌只有4株,数量偏少,是低温抑制了高、中温菌种的生长活性。低温会抑制中温微生物以及高温微生物的生长,耐低温乳酸菌的活性也会受到抑制。在温度较低时,乳酸菌进行物质代谢交换会因为低温环境的刺激而缩小,代谢速率减慢,使得乳酸菌生长活性受到抑制,导致乳酸菌繁殖代谢性能减弱,所以表现为乳酸菌菌体数量少,D(600 nm)值数值低。乳酸菌属于好冷性细菌,在一定温度范围内,短时升温会促进乳酸菌繁殖。然而,足够的高温会破坏菌体蛋白结构,使蛋白变性失活,从而使得乳酸菌的基数减少,生长减弱,菌体数量变少。生长曲线反映出在高温范围D(600 nm)值数值变小,生长曲线呈向下波动,菌体的生长迅速衰弱。
4 结论
植物乳杆菌、清酒乳杆菌、弯曲乳杆菌和屎肠球菌,它们的最适生长温度为20~37℃;当温度高于37℃时,4种乳酸菌的生长量下降,当温度达到50℃时,4种乳酸菌的生长水平非常低;在20℃以下低温范围培养均有较好的耐低温表现。因此,4种乳酸菌均为耐低温菌株,有望成为川西北高寒牧区青贮料调制用青贮添加剂。
猜你喜欢
净水技术(2022年8期)2022-08-10
当代水产(2022年1期)2022-04-26
人民长江(2021年10期)2021-12-09
安徽建筑(2020年11期)2020-11-23
华人时刊(2018年15期)2018-11-10
爱你(2017年7期)2017-11-14
中国调味品(2017年2期)2017-03-20
现代检验医学杂志(2016年3期)2016-11-15
科学之友(2015年6期)2015-08-04
中学科技(2014年6期)2014-08-11
