
家兔CETP基因多态性与肉质性状的关联分析
2019-01-17 10:28党树璋贾先波陈仕毅赖松家
四川农业大学学报 2018年6期
王 杰,党树璋,罗 刚,贾先波, 陈仕毅, 赖松家*
(1.四川农业大学动物科技学院;2.四川农业大学动物遗传研究所,成都 611130)
CETP基因编码血浆胆固醇酯转移蛋白(cholesteryl ester transfer protein,CETP),参与了胆固醇逆向转运的调节和高密度脂蛋白-胆固醇(HDL-C)的代谢,被认为是影响人类健康长寿的重要候选基因[1]。研究表明[2-4],猪的CETP基因位于6号染色体,在脂肪代谢转运蛋白中具有重要作用,被认为是影响动物体脂沉积、生产性能的候选基因。据联合国粮农组织调查,兔肉在欧洲许多国家,如意大利、捷克、西班牙、比利时、卢森堡、葡萄牙、法国,还有一些北非国家(埃及、阿尔及利亚)受到消费者的青睐,兔肉生产在国民经济中发挥着重要作用[5-6]。兔肉能提供丰富的营养物质,首先蛋白含量很高,尤其是限制性氨基酸水平较高。家兔背最长肌和后腿的蛋白含量高达22%。同时矿物质、维生素、抗氧化因子含量高,而兔肉胆固醇和脂肪含量低,被认为是消费者首选的功能性食品[5]。但是,家兔育种,尤其是我国的肉质育种方向研究滞后,国外优良的家兔配套系在我国未得到大面积推广,国内也未见培育效果好的配套系,家兔的分子育种研究少之又少[7]。迄今为止,未见CETP基因在家兔育种方面的研究报道。为此,本研究以3个家兔品种为材料,采用PCR产物直接测序技术,探讨CETP基因对家兔肉质性状的影响,以期为家兔肉质早期育种提供理论基础。
1 材料和方法
1.1 材料
本实验356份兔耳组织样品(伊拉兔124只,香槟兔143只,天府黑兔89只)采集于四川农业大学家兔实验场,并于-70℃冰箱保存备用。本实验家兔在70日龄屠宰,耳组织样品按试剂盒(Axygen,USA)提供的方法提取全基因组DNA,并将其保存于-20℃冰箱,实验所用家兔背最长肌和后腿肌样品保存于4℃冰箱备用。
1.2 方法
1.2.1 肉质性状测定的方法
家兔屠宰率按常规方法计算。pH值测定采用pH仪(Model PH-STAR CPU,MeisterR,Germany)完成,测定探针深入肌肉3cm;肉色测定采用R.L.Van Laack等报道的比色体系[8],数据由亮度(L*),红度(a*)和黄度(b*)组成。pH和肉色测定两次,分别为刚屠宰的肉和屠宰后24 h的肉。肌内脂肪(intermucal fat,IMF)采用矫正的Soxhlet方法[9]测定刚屠宰的肌肉。
1.2.2 PCR 与测序
根据Ensembl报道的家兔CETP基因序列NO.ENSOCUG00000013137采用Premier 5软件设计一对引物:CETP-F:TCCACTTGTTCCCTCAGCCTC 和CETP-R:CAGCGGCTGTGTACTCACTCG。PCR反应采用25 μL反应体系:50 ng全基因组DNA,上下游引物各 1 μmol/L,1.5 mmol/L MgCl2,200 μmol/L dNTPs(dATP,dTTP,dCTP 和 dGTP),0.4 个单位的 Taq DNA聚合酶(MBI),2.5 μL 的 100×buffer。PCR 扩增程序:95℃预变性 5 min;40个循环,94℃ 45 s,56℃ 45 s;72℃ 40 s;最后72℃延长10 min。PCR产物用2%的琼脂糖凝胶电泳检测扩增结果。扩增效果良好的PCR产物送上海英骏生物技术有限公司纯化测序。
1.3 数据分析
1.3.1 基因多态性分析
采用DNAstar软件进行序列对比判断突变位点及其基因型,并进一步计算得到基因型和基因型频率。利用POPGENE软件测试哈温平衡的χ2值。多态信息含量(PIC)通过下列公式计算得到。PIC>0.5为高度多态;0.25<PIC<0.5 为中度多态;PIC<0.25 为低度多态。

1.3.2 基因型与家兔屠体性状关联分析
本实验数据采用 SPSS 21(IBM,Armonk,NY,USA)中一般线性模型(GLM)进行关联分析,分析的线性模型为:Yijk=μ+Gi+Sj+eijk,Yijk为个体性状观测值,μ为个体性状均值,Gi为基因型效应,Sj为个体性别效应,eijk为随机误差效应。数据计算方式均采用最小二乘法和标准误,并用Bonferroni T检验。
2 结果与分析
2.1 家兔CETP基因多态性
实验结果发现,在CETP基因的第4外显子的第11 bp处发生了单核苷酸变异,产生3种基因型为AA、GG和AG,见图1。3个家兔群体的基因多态性分析结果,见表1。由表1可以看出,GG为优势基因型,G为优势基因。最小等位基因频率变化范围为:0.363 6~0.421 3,表示该突变位点较多态。多态信息含量PIC结果显示,本研究的3个家兔群体均处于中度多态,能表现出明显的群体特征。而且χ2检测结果显示家兔群体偏离哈温平衡,表示3个家兔群体均未达到遗传平衡的状态。
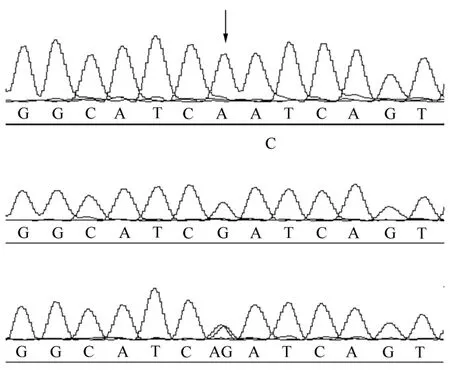
图1 家兔CETP基因型测序图谱Figure 1 Sequencing map of three genotypes of the rabbit CETP gene
2.2 家兔CETP基因多态与屠体性状的关联分析
3个家兔群体不同基因型与屠宰率的相关分析结果,见表2。可以看出,不同基因型家兔全净膛屠宰率差异不显著(P>0.05),但 GG 和 AG 基因型的全净膛屠宰率数值高于AA基因型。GG基因型家兔的半净膛屠宰率最高,与AA和AG基因型差异显著(P<0.05)。
不同基因型与家兔背最长肌肉质性状的相关分析结果,见表3。可以看出,AA基因型的新鲜兔肉的红度最高,与GG和AG基因型的红度差异显著(P<0.05)。而且,AA 基因型的 24 h 肉的红度也最高,与 AG 基因型差异不明显(P>0.05),但与 GG 基因型差异显著(P<0.05)。AG基因型的兔背最长肌肌内脂肪含量最高,而且与AA基因型差异显著(P<0.05)。
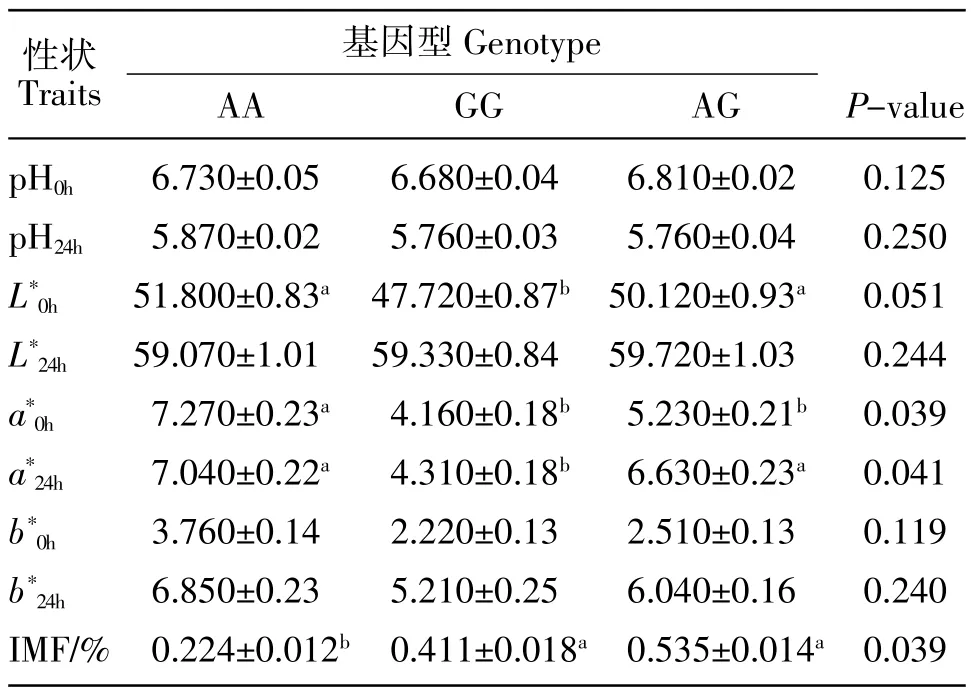
表3 家兔CETP基因多态性与背最长肌肉质性状的相关分析Table 3 Association between SNP of CETP gene and meat traits in rabbit longissimus dorsi muscle
不同基因型与家兔后腿肌肉质性状的相关分析结果,见表4。可以看出,AG基因型新鲜兔肉亮度最大,与 AA、GG 基因型亮度差异显著(P<0.05)。AA基因型24 h兔肉红度最大,与GG和AG基因型差异显著(P<0.05)。AA基因型的家兔后腿肌内脂肪含量最高,与 AG 基因型差异显著(P<0.05),但与 GG基因型差异不显著(P>0.05)。
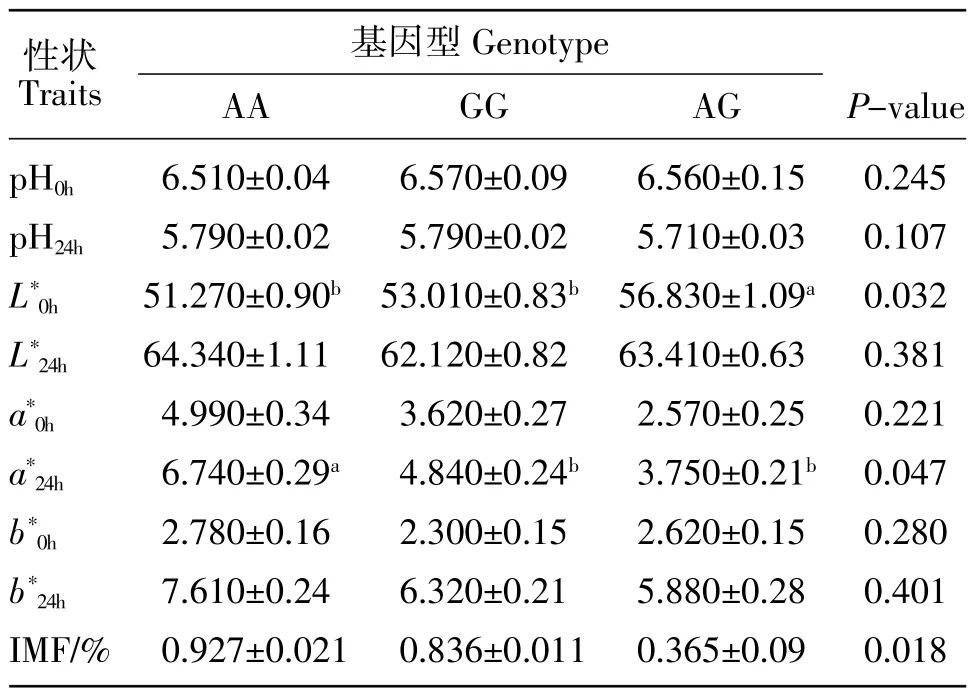
表4 家兔CETP基因多态性与后腿肌肉质性状的相关分析Table 4 Association between SNP of CETP gene and meat traits in rabbit biceps femoris muscle
3 讨论
由实验结果可以看出,家兔CETP基因的3种基因型明显影响了屠宰率。无论是全净膛屠宰率还是半净膛屠宰率,GG基因型的数值都是最高的。说明了GG基因型可有效提高家兔的屠宰率,G等位基因可能具有提高屠宰率的作用。pH值、肉色和肌内脂肪是衡量肉质最重要的标准。无论是新鲜的还是储存24 h的背最长肌和后腿肌,其pH值不受核苷酸变异的影响(P>0.05),但储存 24 h 的肉,pH 值均会下降,这与F.Hulot和J.Ouhayoun研究结果[10]一致,家兔刚屠宰时兔肉pH值接近中性,但屠宰后会迅速下降。而且,C.P.Zeferino等[11]发现储存24小时和48小时的兔肉pH值没有显著变化。肉色研究结果与 G.A.María 等[12]的研究一致,本研究只是发现CETP基因显著影响了家兔的两个时间的背最长肌红度(P<0.05),且 AA 基因型红度最大,AG 型次之,GG型最小。CETP基因只对后腿肌的L*0h和a*24h有显著影响(P<0.05)。与猪肉相比,无论是背最长肌还是后腿肌的肌内脂肪都是最低的[13],本研究发现CETP基因变异对家兔背最长肌和后腿肌的肌内脂肪均有显著影响(P<0.05),值得一提的是,家兔背最长肌AG基因型肌内脂肪数值最高,与AA基因型差异显著(P<0.05),后腿肌却相反,AA 基因型肌内脂肪数值最高,与AG基因型差异显著(P<0.05),其原因有待于进一步深入研究。
4 结论
本研究发现CETP基因第4外显子第11 bp处有3个基因型,且G为优势等位基因。该变异位点显著影响了家兔半净膛屠宰率,背最长肌的红度。同时,该变异位点显著影响了家兔背最长肌和后腿肌的肌内脂肪含量。由此可以推测,CETP基因可能是影响家兔肉质性状育种的候选基因。
猜你喜欢
国外畜牧学·猪与禽(2020年8期)2020-11-30
中国牛业科学(2020年4期)2020-08-15
中国食品(2020年13期)2020-07-29
新世纪智能(高一语文)(2020年12期)2020-06-01
食品安全导刊·下旬刊(2019年7期)2019-10-21
食品安全导刊(2019年21期)2019-01-05
农家致富顾问·下半月(2018年1期)2018-10-21
电子技术与软件工程(2017年8期)2017-05-10
科技视界(2016年27期)2017-03-14
湖北农业科学(2016年18期)2016-12-08
