
青藏高原腹地不同海拔带青藏公路取土迹地恢复草地植物群落的特征
2019-04-04 03:23舒朝成张红艳张雯娜郭正刚
草业科学 2019年3期
王 娟,舒朝成,张红艳,张 静,张雯娜,郭正刚
(兰州大学草地农业生态系统国家重点实验室/兰州大学农业农村部草牧业创新重点实验室/兰州大学草地农业科技学院,甘肃 兰州 730020)
青藏高原是我国西部大开发和一路一带战略建设的重点地区之一,而大量线状工程(如铁路、公路、电网、电缆)是国家这些战略顺利实施的保障。线状工程施工时不可避免的要穿越青藏高原天然草地[1],形成大量工程取土迹地,这些工程取土迹地的原有植被被直接移除[2-3],一方面严重影响高寒草甸中以芽库繁殖为主的植物种群自我更新能力[4],造成草地景观的碎化[5];另外一方面,破坏土壤团聚体结构,增加地表水分蒸发,降低土壤储水能力,造成严重水土流失[6-7]。因此,恢复这些工程取土迹地的植被不仅有利于控制水土流失,增强水源涵养能力、改良土壤结构[8],而且有利于修复高寒草甸景观,提高工程基础的安全性和维持高寒草原生态系统功能的整体性[9]。
鉴于人工恢复工程迹地的植被具有费时费力、成本很高的问题[10],自然恢复成为青藏高原工程取土迹地植被恢复的主要方式[11],其不仅具有低成本、省时、省力特点,而且具有避免草皮移植而损伤其他原生植被的优点[12]。已有研究表明,工程取土迹地的恢复区域植物群落和天然未退化草地植物群落在高度、覆盖度及植物组成等方面存在巨大差异[13],这不仅与取土迹的地面积大小有关[3],而且与海拔有关[2]。海拔是影响植物群落特征的主要因素之一[14-15],虽然青藏高原地区天然植物群落特征与海拔变化关系的研究已经很多[16-18],且发现植物经济类群在不同海拔带的表现存在分异[19],但取土迹地恢复区域植物群落沿海拔变化特征是否类似于天然未退化草地植物群落沿海拔的变化特征,尚需要科学研究提供证据。
虽然反映植物群落特征的指标较多,但覆盖度和高度是植物群落特征的表观指标[20],生物量是植物群落生产力的指示者[21],物种多样性是衡量植物群落内物种丰富程度和分布现状的核心指标[22],也是维持草地生态系统稳定性的关键因素。植物经济类群兼顾草地植物群落的自然属性和经济属性[23],能更好地体现不同生产性能植物在恢复过程中充当的关键地位。因此,本研究以不同海拔带的天然未退化草地植物群落为对照,从经济类群和植物群落总数量指标两个层面分析取土迹地植物恢复区域植物群落覆盖度、高度、物种多样性,以及生产力沿海拔变化的特征,阐明青藏高原腹地恢复区域植物群落特征的垂直分布特征,以及工程取土迹地恢复区域植物群落与天然未退化草地植物群落沿海拔变化特征是否趋同,为取土迹地植物恢复提供科学基础。
1 材料方法
1.1 研究区概况
研究地区位于青藏公路沿线西大滩(94°16.830'E,35°44.578' N)至风火山 (92°55.111' E,34°40.749'N)段,地处青海省西南部的三江源地区,北缘为昆仑山脉,南缘为唐古拉山脉西段,海拔介于4 200-5 000 m,是青藏高原多年冻土的分布区。该区冬季寒冷漫长,夏季凉爽短促,属高原亚寒带半干旱气候。年均温-4.1~-1.3 ℃,昼夜温差大,无霜期 100~200 d,降水量 200~400 mm 且集中在 7-9月份。植被类型主要有高寒草原、高寒草原化草甸和高寒草甸,优势种分别为紫花针茅(Stipa purpurea)、青藏苔草(Carex moorcroftii)和高山嵩草(Kobresia pygmaea),主要伴生种有冷地早熟禾(Poa crymophila)、扇穗茅(Littleda leatibetica)和矮火绒草(Leontopodium nanum)等。
1.2 研究方法
1.2.1 试验设计与取样
调查样地选择1994 年青藏公路扩建和修缮时遗留的取土迹地。由于取土迹地面积和大小不一,植物的恢复状态会存在差异[3]。为了便于比较,本研究选择常见的,分布比较广泛的取土迹地为研究对象,其面积约为150 m2,取土深度约30 cm,且形状为狭长椭圆形对象。自西大滩开始,海拔每上升100 m设置一个海拔梯度,共设7个海拔梯度,分别为 4 315、4 415、4 515、4 615、4 715、4 815、4 915 m。在每个海拔梯度下,选择 3 个取土迹地,调查自然恢复状态下植物的群落特征,相应的在邻近未受干扰的天然草地上选择3个和恢复迹地面积大小一致的样地,作为对照植物群落。每个样地内设置 3 个 1 m×1 m 的样方,共计42个样地,126个样方。样方调查包括记录样方内物种数、测定植物群落总覆盖度和草层高度,并分物种齐地面收集其地上生物量。测量植物高度时,在样方内随机选取100株植株利用钢卷尺测定其株高,如不够100株则全部测定,以其平均值作为该物种的株高,群落高度为各种群高度的平均值之和;物种盖度利用1 m×1 m的样方框,用针刺100次获得群落内每一物种的盖度;地上生物量在105 ℃烘箱内杀青30 min后,在75 ℃烘至恒重后称重。以草地植物资源的自然属性和经济属性为依据,从植物形态特征、种类组成以及自然和经济生产等方面将其划分为豆科(Leguminosae)、禾本科 (Graminea)、莎草科 (Cyperaceae)和杂类草(weed)4个经济类群[23],各经济类群的覆盖度为各经济类群内所有植物分种覆盖度之和,高度为各经济类群内所有植物物种的平均高度,生物量为各经济类群内所有植物种的地上生物量之和。野外调查与取样时间为2016 年7月中下旬。
1.2.2 计算方法
物种重要值计算公式如下:

式中:Pi为种i的重要值。
物种多样性测度采用多样性指数、丰富度指数和均匀度指数,其中多样性指数采用Shannon-Wiener指数(H),计算公式如下:
丰富度指数(S)采用Patrick指数,计算公式如下:
S=样方内出现的物种总数。
均匀度指数(J)采用Pielou,计算公式如下:
J=H/lnS。
1.3 数据处理
以海拔和群落类型(天然和恢复)作为变量因素,采用Two-way ANOVA分析植物群落地上生物量、高度、覆盖度、物种丰富度指数、多样性指数、均匀度指数,以及植物群落各经济类群地上生物量、高度和覆盖度的方差显著性。若差异不显著,不做继续分析;若差异显著,则采用Duncan法对不同海拔梯度天然未退化草地植物群落和恢复区域植物群落的地上生物量、高度、覆盖度、植物群落的各多样性指数以及植物群落各经济类群地上生物量、高度和覆盖度进行多重比较。采用 Excel 2007 和 SPSS 23.0 软件对数据进行整理和分析,使用 Sigma Plot 12.5 软件制图。
2 结果与分析
2.1 植物群落高度及覆盖度沿海拔梯度的变化
2.1.1 群落高度和覆盖度沿海拔梯度的变化
海拔、群落类型以及二者的交互作用均显著影响了植物群落高度和覆盖度 (PA<0.01,PC<0.01,PA×C<0.01)。随海拔高度增加,天然未退化草地植物群落高度和覆盖度表现为先无显著变化(P>0.05)后 显 著 降 低 (P<0.05), 当 海 拔 4 315~ 4 515 m时天然未退化草地植物群落的高度和覆盖度均无显著变化,当海拔为4 815~4915 m时天然未退化草地植物群落的高度差异不显著(图1)。恢复区域植物群落的高度和覆盖度随海拔升高均呈先升高后降低趋势,分别在 4 515 m 和 4 615 m 时最大。天然未退化草地植物群落和恢复区域植物群落高度及覆盖度在不同海拔带表现不一致,海拔为4 515 m和4 615 m时,天然未退化草地植物群落和恢复区域植物群落覆盖度和高度差异不显著,海拔为4 715 m时两个群落的覆盖度无显著差异,在其他海拔时天然未退化草地植物群落覆盖度和高度显著高于恢复区域植物群落的覆盖度和高度。
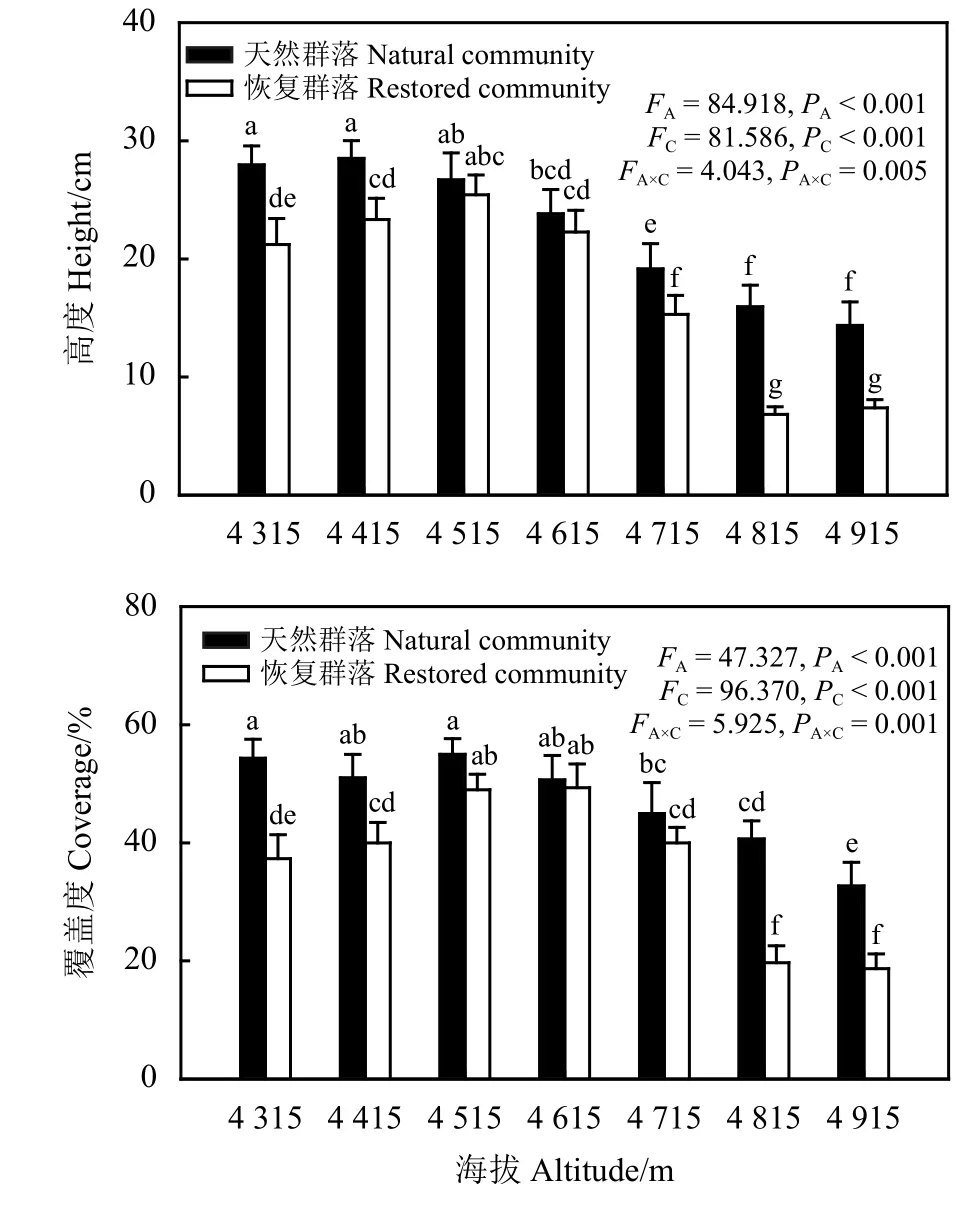
图 1 植物群落高度及覆盖度沿海拔梯度的变化Figure 1 Variation of height and coverage in plant communities along elevation gradients
2.1.2 各经济类群高度、覆盖度沿海拔梯度的变化
海拔和群落类型均显著影响植物各经济类群的高度(PA<0.01,PC<0.01),二者交互作用显著影响豆科、杂类草和莎草科植物高度(PA×C<0.01),但对 禾本科植物高度没有显著影响(图2)。随海拔增加,天然未退化草地植物群落和恢复区域植物群落中豆科、杂类草和莎草科植物的高度呈先增加后降低趋势,其中豆科植物在海拔为4 615 m时最高,莎草科在 4 415 m 时最高,杂类草植物在 4 515 m 时最高,但海拔为 4 415 m 和 4 915 m 时恢复区域植物群落中没有豆科植物出现,海拔为4 815 m时恢复区域植物群落中没有莎草科植物出现;天然未退化草地植物群落内禾本科植物高度随海拔增加呈降低趋势,而恢复区域植物群落内禾本科植物高度随海拔升高呈先升高后降低趋势。除海拔4 315 m和4 715 m外,豆科植物高度在天然未退化植物群落内显著高于恢复群落内(P<0.05),而杂类草植物和莎草科植物高度除在海拔 4 515~4 815 m 时和 4 415~4 615 m时天然未退化草地植物群落和恢复区域植物群落间差异不显著外,在其他海拔时天然未退化草地植物群落内其植物高度均显著高于恢复区域植物群落内。

图 2 植物各经济类群高度沿海拔梯度的变化Figure 2 Variation of height in different plants of economic value along the elevation gradients
海拔和群落类型显著影响了植物群落各经济类群覆盖度(PA<0.01,PC<0.01),二者交互作用显著影响了豆科、禾本科和莎草科植物覆盖度(PA×C<0.01),而对杂类草植物覆盖度影响不显著(P>0.05)(图3)。天然未退化草地植物群落和恢复区域植物群落内豆科植物覆盖度随海拔升高呈先升高后下降趋势,均在海拔为4 615 m时最高;天然未退化草地植物群落和恢复区域植物群落内禾本科植物的覆盖度基本随海拔升高均呈递减趋势,但海拔4 415 m 时恢复区域植物群落覆盖度低于海拔 4 515 m时恢复区域植物群落覆盖度;天然未退化植物群落和恢复区域植物群落内莎草科植物覆盖度随海拔增加均呈现先上升后降低趋势,天然未退化草地植物群落和恢复区域植物群落内莎草科植物覆盖度均在海拔为 4 515~4 615 m 时最高;天然未退化草地植物群落和恢复区域植物群落杂类草植物覆盖度随海拔升高均呈先上升后降低趋势。天然未退化草地植物群落内豆科和禾本科植物覆盖度均在海拔 4 315 和 4 915 m 时显著高于恢复区域植物群落豆科和禾本科植物覆盖度(P<0.05),而天然未退化草地植物群落内莎草科植物覆盖度则在海拔 4 315 m 和 4 815~4 915 m 时显著高于恢复区域植物群落内莎草科植物覆盖度。
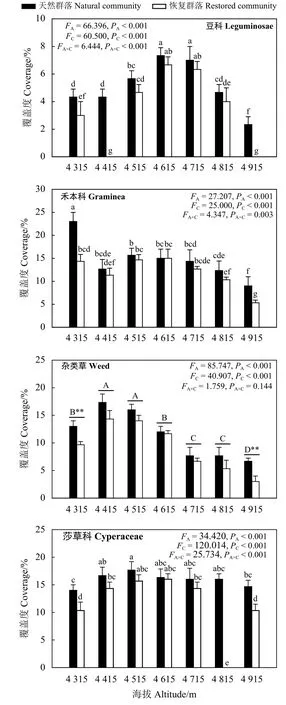
图 3 植物各经济类群覆盖度沿海拔梯度的变化Figure 3 Variation of coverage in different plants of economic value along the elevation gradients
2.2 植物群落地上生物量沿海拔梯度的变化
2.2.1 地上总生物量沿海拔梯度的变化
海拔、群落类型及二者交互作用对植物群落的地上生物量均具有显著影响(PA<0.01,PC<0.01,PA×C<0.01)(图4)。随海拔升高,天然未退化草地植物群落地上生物量表现为先显著降低(P<0.05),后变化差异不明显,当海拔在 4 515~4 715 m 时各海拔梯度间天然未退化草地植物群落地上生物量差异不显著 (P>0.05),海拔 4 815~4 915 m 天然未退化草地植物群落地上生物量差异不显著;而恢复区域植物群落地上生物量随海拔升高呈先降低后升高再降低趋势,在海拔4 615 m时恢复区域植物群落地上生物量最高。除海拔4 615 m外,天然未退化草地植物群落的地上生物量均显著高于恢复区域植物群落。
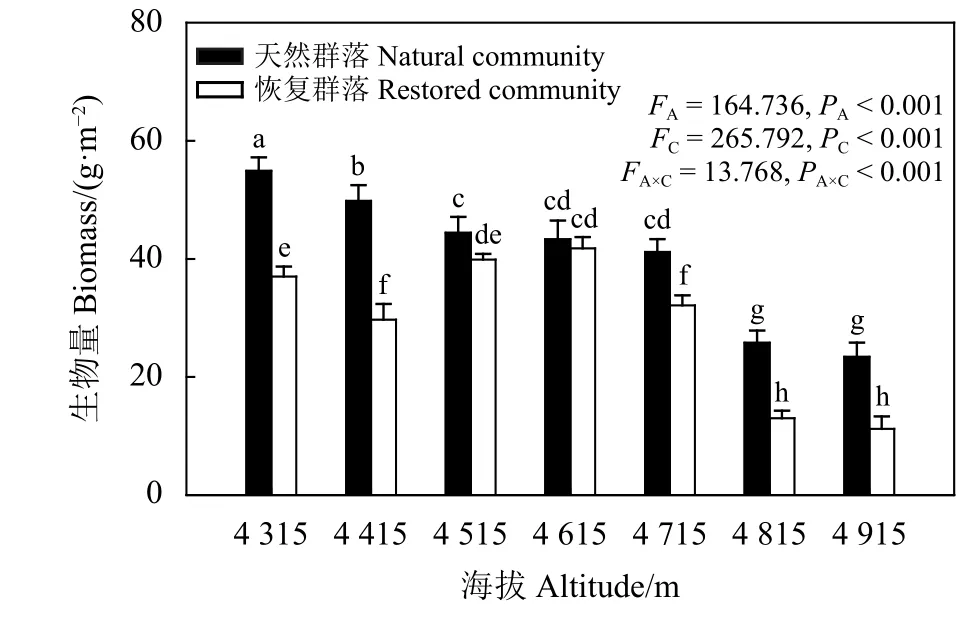
图 4 植物群落地上生物量沿海拔梯度的变化Figure 4 Variation of above-ground biomass in plant communities along elevation gradients
2.2.2 各经济类群地上生物量沿海拔梯度的变化
海拔、群落类型以及二者的交互作用均显著影响植物群落各经济类群地上生物量(PA<0.01,PC<0.01,PA×C<0.01)(图 5)。天然未退化草地和恢复区域植物群落内豆科植物地上生物量随海拔上升均先上升后下降,且均在海拔为4 615 m时最高;天然未退化草地植物群落和恢复区域植物群落内禾本科和杂类草植物地上生物量随海拔上升均呈下降趋势;天然未退化草地植物群落和恢复区域植物群落内莎草科植物地上生物量随海拔上升均呈先升高后降低趋势,天然未退化草地植物群落在海拔为4 515 m时莎草科植物地上生物量最高,恢复区域植物群落也在在海拔为4 515 m时莎草科植物地上生物量最高。除海拔为4 515 m外,天然未退化草地植物群落豆科植物地上生物量均显著高于恢复区域植物群落(P<0.05);禾本科和莎草科植物地上生物量分别除在海拔 4 515~4 815 m 时和4 515~4 615 m 时差异不显著外 (P>0.05),在其他海拔高度下天然未退化草地植物群落禾本科和莎草科植物地上生物量均显著高于恢复群落;杂类草植物地上生物量除在海拔4 615 m时外,在其他海拔梯度下天然未退化草地植物群落的杂类草植物地上生物量均显著高于恢复区域植物群落。

图 5 植物群落各经济类群地上生物量沿海拔梯度的变化Figure 5 Variation of above-ground biomass in different plants communities of economic value along the elevation gradient
2.3 植物群落物种多样性沿海拔梯度的变化
群落类型对植物群落丰富度指数和多样性指数具有极显著影响(P<0.01)(表1),但对均匀度指数影响不显著;海拔对均匀度指数具有显著影响(P<0.05),对多样性指数有极显著影响 (P<0.01),但对丰富度指数影响不显著;二者交互作用对多样性指数具有极显著影响(P<0.01),对丰富度指数和均匀度指数影响不显著。随海拔增加,多样性指数和均匀度指数均呈先增大后降低趋势(P<0.05),在海拔 4 715 m 时最高。除海拔为 4 515~4 715 m外,天然未退化草地植物群落多样性指数均高于恢复区域植物群落多样性指数(P<0.01)。

表 1 植物群落物种多样性沿海拔梯度的变化Table 1 Variation in plant community species diversity along the elevation gradient
3 讨论
恢复工程取土迹地植被是青藏高原地区高寒草甸景观修复的重要内容,也是青藏高原退化草地生态系统恢复的重要组分[9]。海拔变化会重新配置区域内水热资源[24],青藏高原生态脆弱,区域内气象条件变化明显,高海拔生境内分布有大面积的腹地[25],因此植物群落沿海拔梯度的变化特征较为复杂,而取土迹地恢复植物群落属于次生群落,其沿海拔变化特征与天然植物群落相比,既具有趋同性,又具有分异性[2, 5]。本研究结果表明,天然未退化草地植物群落高度、覆盖度及地上生物量随海拔梯度增加呈降低趋势,而恢复区域植物群落高度、覆盖度和地上生物量则呈先增加后降低趋势,主要是天然未退化草地植物群落在长期进化和演替过程中形成了相对稳定的垂直分布格局,随海拔上升,气温和土壤温度降低[26],降水量和降水频次增多[27-28],光照强度和紫外辐射增强[29],其中过多降水和较低温度使部分植物遭受低温胁迫[30],影响植物光合作用等生理生化过程[31],降低微生物活性,减缓有机质降解速率[32],植物正常生长受抑制[33],同时强紫外光抑制细胞伸长,致使茎秆粗短,植物群落高度降低,另外强光照强度和紫外辐射会减少植物叶片干物质量,叶面积缩小,因此植物群落总覆盖度下降,从而导致天然区域植物群落的地上生物量降低。针对恢复区域植物群落而言,其适应海拔梯度的时期相对较短,植物群落垂直格局更容易受到外界水热组合的影响,恢复区域植物群落较天然未退化草地植物群落具有更多的裸露地表,土壤表层蒸发强烈[34],加之低海拔区域降水较中间海拔地区少,而温度较中间海拔地区高,土壤表层水分散失严重,这会严重抑制低海拔地区恢复区域植物群落植物的生长[35],因此低海拔地区恢复区域植物群落植物的高度、覆盖度和地上生物量低于中间海拔区域。高海拔地区降水增多,植物生长的水分限制因素解除,而低温胁迫加剧,土壤肥力下降,太阳辐射增强,导致高海拔地区植物的正常生长受抑制,因此,恢复区域植物群落的高度、覆盖度和地上生物量在高海拔地区低于中间海拔区域。
海拔梯度变化不仅影响着植物群落的高度、覆盖度和地上生物量,同时也影响着植物各经济类群的高度、覆盖度和地上生物量。天然未退化草地植物群落内禾本科植物的高度、覆盖度和地上生物量随海拔梯度增加呈递减趋势,恢复区域植物群落内除禾本科植物的高度随海拔梯度增加呈先增加后降低外,禾本科植物的覆盖度和地上生物量随海拔梯度增加呈递减趋势,主要是禾本科分蘖能力强,叶面积较大且叶片分布在不同高度层,禾本科植物在降水量低、热量高的环境中较其余植物经济类群具有较高的竞争优势[36],因此海拔较低时禾本科植物光合速率相对较高[37],其高度、覆盖度和地上生物量较高。海拔较高时,虽然降水增多,但禾草对温度更加敏感[38],低温抑制其伸长生长,迫使其分蘖数减少[39],竞争能力减弱,出现海拔较高时天然未退化草地植物群落内禾本科植物群落的高度、覆盖度和地上生物量下降的现象。工程迹地因裸露地表蒸发强烈,低海拔区域土壤含水量较中间海拔区域低,因此海拔较低时恢复区域植物群落内禾本科植物可利用水分相对减少,这一定程度上抑制了禾本科植物株高生长,但刺激了分蘖数[39],增加了生物量,因此恢复区域植物群落的覆盖度和地上生物量较中间海拔区域是高,而海拔较高时则比中间海拔时低。天然未退化草地植物群落和恢复区域植物群落内杂类草植物经济类群的高度和覆盖度随海拔梯度增加呈先增加后降低趋势,地上生物量随海拔增加呈逐渐降低趋势,这是因为杂类草植物经济类群因同样对热量需求较高,因此其地上生物量在低海拔地区相对较高,但由于部分物种分布范围的差异性,其高度和覆盖度在中低海拔区域最高;随海拔高度增加,天然未退化草地植物群落和恢复区域植物群落内豆科植物、莎草科植物经济类群的高度、覆盖度及地上生物量均表现为先增加后降低趋势,这可能是由于莎草科植物和部分豆科植物在长期的进化过程中适应了青藏高原寒冷的气候特点[30],但对降水和太阳辐射因素仍较为敏感[40],在众多因素综合作用下中度海拔区域条件最好,因此豆科植物、莎草科植物经济类群在天然未退化草地植物群落和恢复区域植物群落内的覆盖度、高度和地上生物量最高。天然未退化草地植物群落和恢复区域植物群落及各经济类群植物群落的高度、覆盖度和地上生物量在中间海拔4 615 m左右时差异不显著,而在其它海拔梯度下天然未退化草地植物群落高度、覆盖度及地上生物量均显著高于恢复区域植物群落,这表明工程迹地上的群落在中海拔区域时天然未退化草地植物群落和恢复区域植物群落及各经济类群植物群落恢复的最好。
天然未退化草地植物群落和恢复区域植物群落中多样性指数随海拔梯度变化均呈先升高后降低趋势,而丰富度指数和均匀度指数则随海拔梯度的上升无明显差异,主要是由于海拔升高时水热因素的差异所致[24],加之太阳辐射增强影响到各经济类群的分布[41],使其垂直分布特征产生变化。低海拔地区植物生长季节热量充足、水分不足,物种多样性低;高海拔地区虽然水分充足,但热量不足,水热条件不适于某些物种生存,亦降低物种多样性;中度海拔梯度地区,尽管水分、热量不是最高,但水热组合最佳,适于植物生长,因此物种多样性高。
4 结论
以不同海拔带天然未退化草地植物群落为对照,从经济类群和植物群落两个层面分析了青藏公路取土迹地植物恢复区域植物群落的特征,发现青藏公路取土迹地恢复区域植物群落和天然未退化植物群落随海拔升高,总群落高度、覆盖度、生物量和多样性指数变化一致,但各个经济类群的高度、覆盖度、生物量变化不一致。青藏公路取土迹地植被经历了22年(1994-2016年)的恢复后,虽然恢复区域植物群落与天然未退化草地植物群落随海拔升高其各自的测度指标变化趋同,但各个经济类群指标沿海拔升高的变化趋势趋异。因此,恢复区域植物群落已经成为稳定群落,还是一直处于正向演化状态,需要从群落稳定性方面继续开展深入研究。
猜你喜欢
科学技术创新(2022年30期)2022-10-21
兰台内外(2021年33期)2021-12-17
农业与技术(2021年23期)2021-12-14
黑龙江水利科技(2020年8期)2021-01-21
原子与分子物理学报(2020年5期)2020-03-17
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
军民两用技术与产品(2018年11期)2018-07-16
现代农村科技(2018年6期)2018-01-18
广东农业科学(2017年5期)2017-08-29
