
基于跨组学技术的育种示踪研究
2019-04-08 01:28程萌杰施利利孙宁张欣丁得亮崔晶边嘉宾通信作者王松文通信作者
天津农学院学报 2019年1期
程萌杰,施利利,2,孙宁,张欣,2,丁得亮,2,崔晶,2,边嘉宾,2,通信作者,王松文,2,通信作者
(1.天津农学院 农学与资源环境学院,天津 300384;2.天津中日水稻品质食味研究中心,天津 300384)
经过六十多年的育种,我国已经选育出7万多优良的水稻品种。在各增产要素中,品种的贡献占40%。多年来,育种者追求的两个重要目标是定向育种和提高育种效率[1]。纵观国内外育种专家的研究发现,一种育种示踪研究是追溯亲本系谱和父母双亲的基因构成及贡献[2],另一种研究是根据设计育种原理进行分步骤育种设计和单元划分,比如:定位所有QTL、鉴定有利等位基因、综合考虑多种因素进行育种设计[3-6]。有研究表明,育种可以分为提高育种和保持育种[7]。在提高育种阶段,往往以提高产量为主要目标。在保持育种阶段,以改善品质或提高抗性为主。以水稻为例,第一次产量大幅度提高是以‘IR8’‘矮脚南特’‘广陆矮4号’等矮杆品种推广为特征;第二次产量大幅度提高是以推广杂交稻为特征。作物育种学的规律是品种更替以里程碑式的品种出现为特征。比如,‘汕优63’于1980年育成,至2002年,‘两优培九’的推广面积超过了‘汕优63’[8]。在保持育种的一段时间内,审定并推广的品种往往没有大幅度的提高,只是品质的改良或抗性的提高。近年来,正在实现由经验育种向精准育种转变,而基于跨组学技术的育种示踪更是研究的热点。育种示踪一方面有助于研究品种形成的机制,另一方面可以实现设计育种,其基本路径是沿着设计育种的路线,一步步改良品种和创造新品种。本文报道本课题组基于跨组学技术进行育种示踪的初步研究结果,以期探索有关育种方法,实现育种的定向性和快周期性。
1 材料与方法
1.1 材料
试验材料如表1所示。
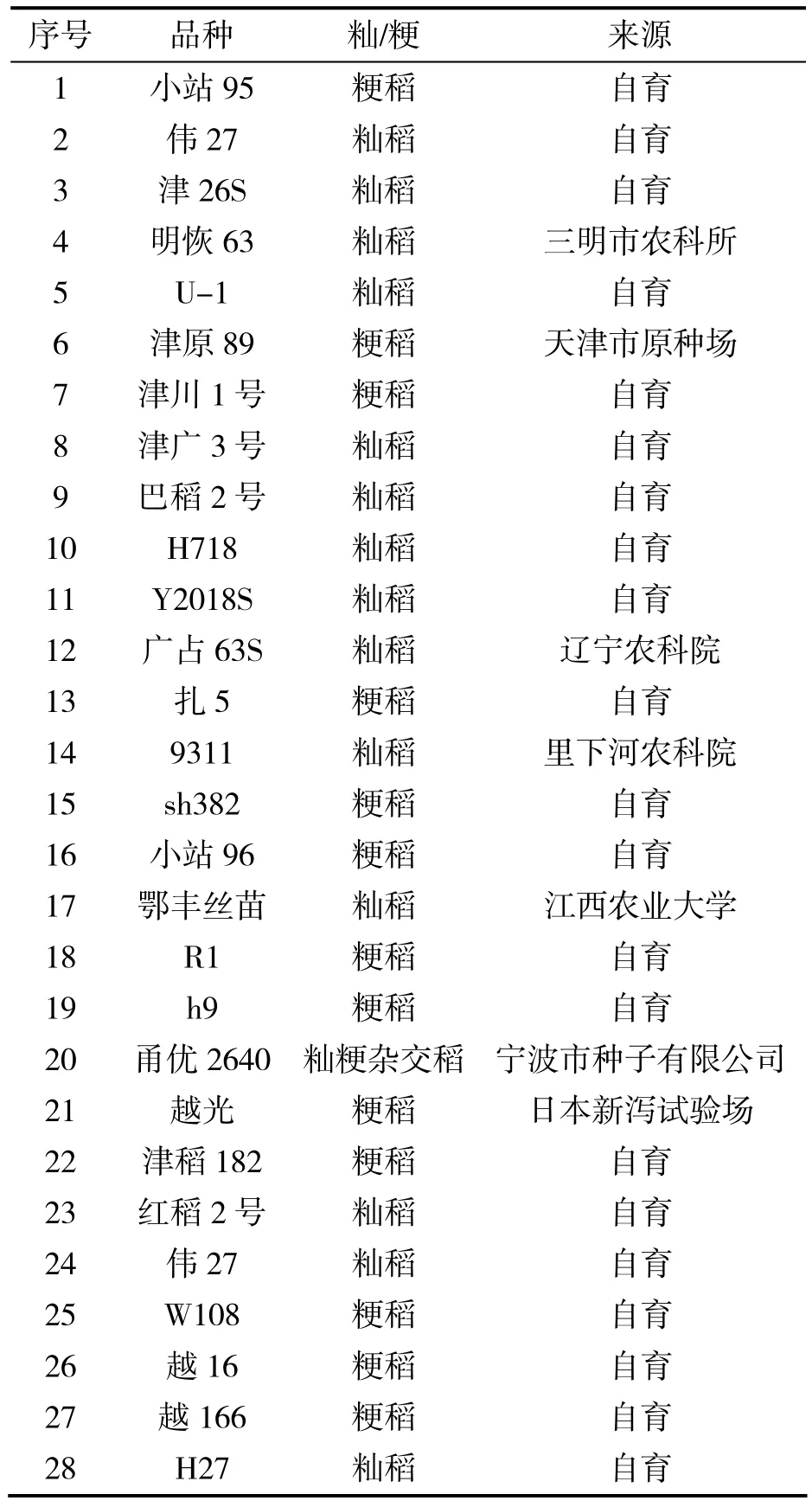
表1 试验材料
试验材料在国家水稻基因组计划构建作图群体材料的基础上进行扩建。基本依据是:(1)用最少的样品尽量多的代表水稻种质的遗传多样性;(2)选取一定量的高产品种和优质水稻品种;(3)部分材料进行了特别改造,如‘U-1’的千粒重31.2 g;‘鄂丰丝苗’高产优质;‘津原89’创造了天津地区水稻高产纪录。
1.2 方法
跨组学示踪包括组学技术和水稻育种综合技术。在组学技术中,包括基因组、转录组、蛋白组、代谢组、表型组,水稻育种综合技术包括常规育种技术、育种新技术(如分子标记技术、基因工程技术等)。本研究依据这些技术,主要利用基因组测序结果、转录组测序结果、蛋白组分析结果,以及课题组多年积累的育种成果进行综合分析。
‘小站95’测序由北京六合华大基因科技有限公司完成。
转录组测序、蛋白组测序由北京诺禾致源生物信息技术有限公司完成。
育种分析、组学分析、生物信息学分析等由课题组独立完成或利用Internet在线软件工具分析完成。
2 结果与分析
2.1 跨组学技术综合分析
跨组学技术构成如图1所示。
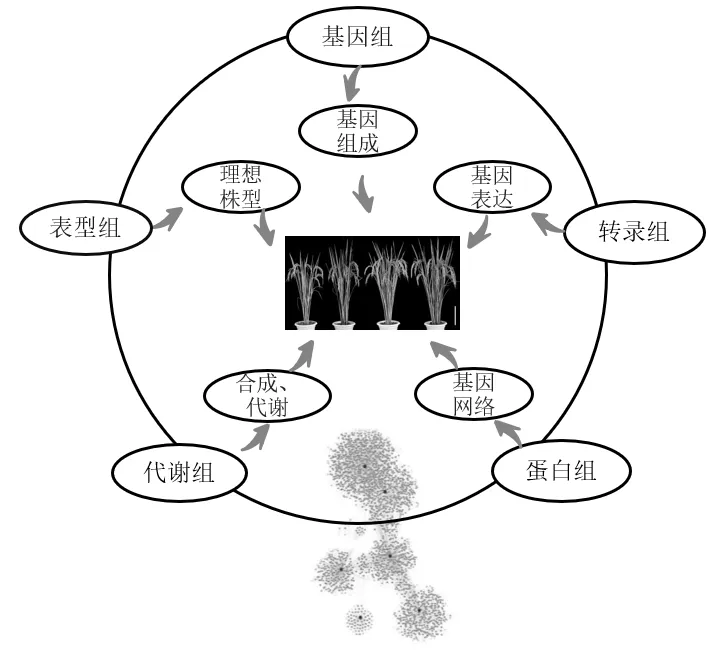
图1 跨组学分析技术构成
采用重测序技术对‘小站95’进行了基因组测序,在基因组测序的基础上对部分染色体区段、基因组可追溯片段(traceable block,TB)进行了基因组从头测序。采用转录组测序对‘津原89’‘津川1号’等高产优质品种进行分析。蛋白质组测序采用iTRAQ技术分析。本研究对‘津优294’(‘津26S’/‘U-1’,课题组选育品种)、‘津原89’(‘津原11’/‘津原E28’)等品种进行了育种示踪。
结果表明,‘小站95’具有粳型特征,第1染色体短臂上端有籼稻序列特征,0~7 569 978 bp为籼稻特征区,8 169 158~28 125 264 bp为粳稻区,28 704 482~38 029 446 bp为籼粳杂交区(分子特征为杂合),39 300 025 bp~末端为粳稻区。因为杂合区的分离,后代出现母本型、父本型、杂交分子型,在杂合区段具有产量QTL、籼粳分化区及与杂种优势相关的QTL。
通过对转录组的分析,测得与淀粉合成相关的代谢途径,涉及了10个淀粉合成的酶类及基因,其中,在4个关键酶中,第1个限速酶——糖苷合成酶的活性有显著差异。
通过对蛋白质组的分析,高产品种和优质食味品种(产量中等,多数品种的产量低于高产品种)的淀粉合成酶活性如图2所示。
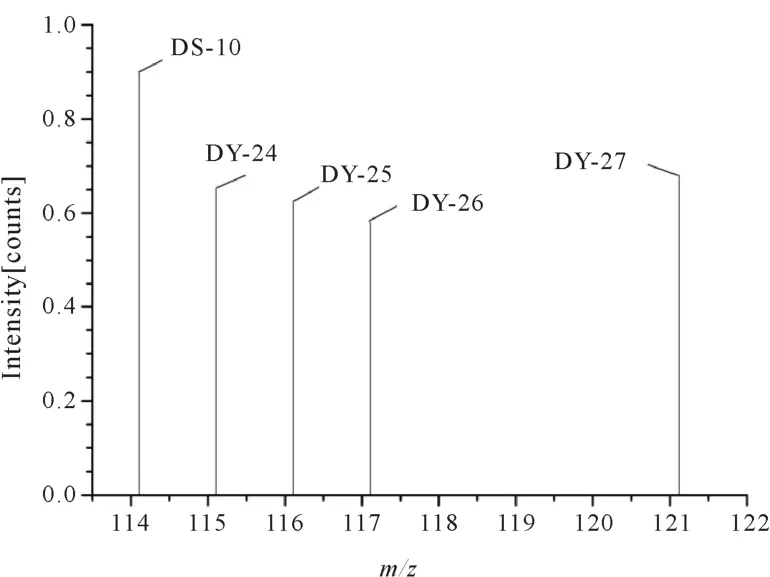
图2 不同品种的糖苷合成酶活性
图2表明,高产品种与优质食味品种糖苷合成酶活性具有明显差别,‘U-1’‘H27’‘W108’均为高产品种,‘越166’‘越16’为优质食味品种,糖苷合成酶活性明显低于高产品种。
该蛋白质组测试的结果得到了转录组测试结果的印证。比较而言,优质食味品种的糖苷合成酶活性较低,高产品种的糖苷合成酶活性较高。由图3可见,高产品种‘津原89’是优质食味品种‘津川1号’的3.2倍。
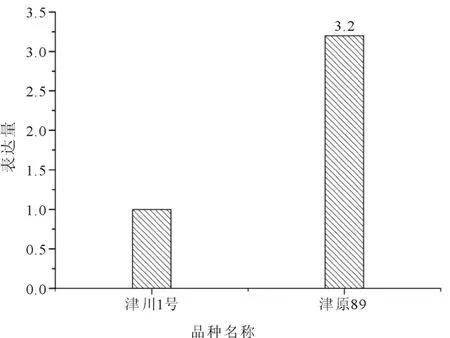
图3 高产品种与优质食味品种糖苷合成酶活性比较
2.2 高产品种的育种示踪
无论是从野生稻向现代水稻品种演化,或是从第一代恢复系到现代恢复系演化,都表现为产量、产量构成因子的线路变化,如图4。

图4 现代品种的二维示踪
图4表明,‘矮脚南特’/‘东乡野生稻’(1次杂交)、‘矮脚南特’/‘东乡野生稻’//‘9311’(经过复交)的产量都明显低于现代品种‘U-1’///‘矮脚南特’/‘东乡野生稻’//‘9311’。从另一个维度看,‘明恢63’的产量不如‘9311’(‘扬稻6号’),‘明恢63’和‘9311’的产量都明显低于现代品种的产量水平。
从系谱分析看,‘明恢63’ /‘小站95’的后代中选出‘伟27’,‘U-1’/‘伟27’的后代中选育出‘津稻294’。从跨组学的分析看,‘小站95’是具有部分籼稻血缘的地方品种,‘U-1’/‘小站95’可以选育出分蘖力强、结实率高的‘伟27’。‘U-1’/‘伟27’后代中可以选育出高产、优质、抗病性好的‘津稻294’,‘津稻294’具有植株高矮好、穗子大小好、分蘖力强弱好的新品种。‘津稻294’具有‘小站95’‘明恢63’‘U-1’等水稻品种的血缘,‘津稻294’聚合了籼、粳稻两个基因组的优良基因,经过两次杂交,跨越了从地方品种到现代品种的快速发展,实现了高产、优质、抗病的有机统一,成为不为多见的优良品种,这个品种为水稻家族增添了新成员。
3 讨论
3.1 育种示踪技术及其在育种中的应用
Zeng等利用3个品种示踪了28个重要农艺性状基因[5],研究表明,籼、粳两个基因组可以进行有效的基因组交流,R1、R2、R3等新品系具有高产品种的产量结构,同时这些新品系也具有‘日本晴’的优质特性。‘9311’(‘扬稻6号’)为亲本的杂交后代并没有像‘明恢63’那样出现131个恢复系,审定了249个新组合。但R1、R2、R3等新品系实现了优化设计育种。
Zhou等以中国华南地区广泛种植的优质水稻品种‘黄华占’及其谱系品系为对象,通过重测序相关研究,鉴定得到重要的功能区域,为现代水稻育种提供了遗传基础和新思路[2]。在这一研究中,结合QTL定位、HTBs(‘黄花占’追溯片段)鉴定,在QTL分析的基础上对应了生物功能基因,这是分子育种和育种示踪的重大进步。本研究与前人研究有所不同,通过跨组学技术的综合应用,结合品种系谱的溯源与示踪,培育出高产优质的新品种。
3.2 跨组学技术应用与现代品种选育
陆姝欢运用跨组学技术研究了小球藻培养过程的代谢机制,开辟了跨组学技术的新领域[9]。在作物育种中,少有运用跨组学技术进行育种示踪的报道。但是,粳稻品种‘日本晴’已经有了水稻基因组的精细图[10]。在籼稻品种中,‘9311’实现了随机测序[11],在绘制‘9311’基因组草图的基础上,已有‘两优培九’杂种优势的机制解析研究[12]。近年来,随着水稻基因组计划的深入开展,已经克隆了2 218个水稻重要农艺性状基因[13-16]。在基因分子生物学的新领域,已经有图位克隆、基因功能分析、两个基因互作[17]、作物品种的代谢途径分析[18]、产量与氮素利用的平衡[19]等研究。由于分子育种滞后于现代品种选育、常规育种技术与现代育种技术难以整合、生物学信息学发展难于结合骨干品种更新换代等原因,利用跨组学技术进行现代品种选育尚缺乏系统的研究,本文的研究结果及相关研究为现代育种提供了新的思路和方法,必然为方兴未艾的分子育种提供方法学和技术上的支持。
猜你喜欢
军事文摘(2022年16期)2022-08-24
今日农业(2021年11期)2021-08-13
中国农业科学(2021年6期)2021-03-25
中国生殖健康(2020年4期)2021-01-18
中国生殖健康(2020年4期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
红楼梦学刊(2020年2期)2020-02-06
中国现代中药(2019年5期)2019-07-03
科海故事博览·下旬刊(2019年6期)2019-04-16
中国生殖健康(2018年4期)2018-11-06
