
不同播期冬小麦小花发育特性与同化物代谢的相关性
2019-04-09 03:50朱元刚肖岩岩初金鹏钤太峰代兴龙贺明荣
植物营养与肥料学报 2019年3期
朱元刚,肖岩岩,初金鹏,张 秀,钤太峰,马 鑫,代兴龙*,贺明荣*
(1 作物生物学国家重点实验室/农业部作物生理生态与耕作重点实验室/山东农业大学农学院,山东泰安 271018;2 德州市农业科学研究院,山东德州 253015;3 德州市实验中学,山东德州 253015)
小麦穗粒数是决定产量形成的关键因素[1]。而穗粒数的形成与穗部小花发育的完善水平密切相关[2]。小麦小花发育是小麦个体发育和群体质量协调发展的综合体现[3],不同小穗和不同小花位的小花因基因型、栽培措施和环境条件的影响而呈现发育不均衡性,导致其结实特性与最终穗粒数存在较大差异[4,12]。小花发育的研究一直是国内外学者的研究热点,前人在小麦的幼穗分化、小花结实及其与植株生长环境的响应等方面已有较多的形态学和生理学研究[1-19]。研究发现,穗粒数的多少很大程度上由存活下来的小花数即开花期可孕花数决定,而可孕花数是由小花分化总数和退化数量共同影响[5]。一般而言,小花分化总数主要由遗传基因控制,对于可孕花数的影响力较小,而小花退化成为决定可孕花数的关键[6-7]。小花退化与穗茎的快速生长同时发生,因此认为开花前茎和穗之间的快速生长造成对有限营养资源的竞争,导致穗部生长营养供应不足,引发了小花的退化[8-10]。矮秆基因 (Rht) 的成功引入实现了可孕花数的提高,这主要归因于降低了茎秆对同化物的竞争力,从而促进开花前较多的干物质分配到穗中[11]。此外,众多研究描述了在不同穗型[12-13]、种植密度[14-15]、氮肥施用[5,16-17]以及光周期[18]、温度[19]等条件下穗部生长变化对于小花发育的影响,较多的研究成果对于明确小花发育结实特性和生理变化特征以及探明小花发育结实成粒的规律意义重大,有力地促进了深化小麦产量形成生理和提高穗粒数的研究。
本课题组在前期研究中证实推迟播期从10月1日至15日能够获得籽粒产量维持,主要归因于提高的穗粒数和降低的单位面积穗数相互弥补从而维持了单位面积粒数[20]。然而,小花生长发育特性与同化物积累、分配以及积累速率等之间的关系尚未深入了解。本试验以泰农18和济麦22为试验材料,设置5个播期处理,重点分析了小麦小花发育过程中分化和退化动态规律及结实特性的差异,探讨了单位面积可孕花数与同化物积累、分配以及积累速率之间的相关性,旨在探索提高小麦粒数的调控机制,以期为丰富冬小麦高产高效稳产栽培理论提供依据。
1 材料与方法
1.1 试验地概况
本试验于2014—2015年和2015—2016年,连续两个小麦生长季在山东省泰安市岱岳区大汶口镇东武村山东农业大学试验田 (35°57′N,117°3′E) 进行。试验地前茬作物为夏玉米,土质为沙壤土,耕层 (0—20 cm) 土壤有机质为15.9 g/kg,全氮1.07 g/kg,有效氮52.8 mg/kg,速效磷25.8 mg/kg,速效钾67.5 mg/kg,pH值为8.13。
1.2 试验设计
选用大穗型小麦品种泰农18 (T18) 和多穗型小麦品种济麦22 (J22) 为试验材料。设置5个播期处理,分别为9月24日 (S1)、10月1日 (S2)、10月8日 (S3)、10月15日 (S4) 和10月22日 (S5)。泰农18和济麦22的种植密度分别为每平方米405株和180株,南北走向种植。试验采取随机区组设计,3次重复,小区面积75 m2(25 m × 3 m) (12行,行距0.25 m),出苗后在三叶期定苗。自小麦第一个分蘖冒出地面2 cm起,用不同颜色塑料环分别标记各处理主茎和第一分蘖、第二分蘖及第三分蘖,用于后期各指标的取样调查。
前茬玉米收获后秸秆全部于小麦播种前翻压还田。播前施入基肥包括纯N 120 kg/hm2、P2O5120 kg/hm2、K2O 120 kg/hm2,在拔节期追施纯N 120 kg/hm2,氮、磷、钾分别为尿素 (N 46%)、过磷酸钙(P2O512%) 和氯化钾 (K2O 60%)。底施的尿素于播种前均匀撒施于各小区后耕翻,拔节期开沟施肥,播种后立即浇水以确保出苗。试验期间充分供水,各生育季均采用杀虫剂、杀菌剂和除草剂防治害虫、疾病、杂草,两个生育季生长过程中冬小麦均未发生病虫草害。其他管理措施均与高产田管理相一致。两个生长季的收获时间分别为2015年6月10日和2016年6月8日。
1.3 测定内容与方法
1.3.1 群体分蘖动态 小麦播后于三叶期调查基本苗数,同时每个试验小区选取长势均匀一致的区域划定1 m × 6行的小区,自小花开始分化至开花期每隔3 d以及成熟期进行群体大小的动态调查,并根据行距计算单位面积茎蘖数,计算公式:单位面积总茎蘖数 = 平均1 m每行茎数/行距 (m)。单位面积有效茎蘖数为成熟期有效穗数。单位面积无效分蘖数为同一时期单位面积总茎蘖数与有效茎蘖数之差。
1.3.2 小花分化发育进程特征调查 自小花开始分化至开花期每隔3 d取样一次,每小区随机选取0.5 m × 2行的样方 (不含边行),将样方内小麦植株按主茎和分蘖标记分开,其中泰农18调查主茎和第一分蘖,济麦22调查主茎和第一、第二和第三分蘖,然后将每个单茎再分为茎 + 鞘、叶和穗等三部分。每个主茎和分蘖随机选取10个幼穗使用显微镜(OLYMPUS-CX22RFS1,Japan) 和解剖镜(OPTIKA-SZM-LED2,Italy) 观测小花分化发育动态,小花的发育特征按照Waddington等[21]的标准观测,记录分化小花数、可孕小花数 (即具有完整绿色花药和羽状柱头的小花数) 和各阶段发育特征及成熟期结实穗粒数。观测完毕后,所有样品用于干物质积累和氮素积累测定。
单位面积最大分化小花数 = ∑每穗最大分化小花数 × 单位面积茎数 (开花前)
单位面积可孕小花数= ∑每穗可孕小花数 × 单位面积有效茎数 (开花期)
单位面积结实粒数= ∑每穗结实粒数 × 单位面积有效穗数 (成熟期)
单位面积退化小花数 = ∑每穗最大分化小花数 ×单位面积无效分蘖数 + (每穗最大分化小花数 - 开花期可孕花数) × 单位面积有效茎数
单位面积小花结实率 = 单位面积结实粒数 ÷ 单位面积最大分化小花数 × 100%
单位面积可孕小花结实率 = 单位面积结实粒数 ÷单位面积可孕小花数 × 100%
单位面积小花存活率 = 单位面积可孕小花数 ÷单位面积最大分化小花数 × 100%
1.3.3 干物质积累 所有样品经105℃杀青30 min后,80℃烘干至恒重,用电子天平称重。开花期单位面积各器官干重 = ∑ (主茎各器官干重 + 分蘖各器官干重) × 4。穗部干物重与茎秆干物重的比值即为干物重穗茎比。退化阶段整株和穗的干物质积累速率分别为此阶段整株和穗的干物质的积累量与持续天数的比值。
1.3.4 氮素积累 将称重后的干样样品使用微量磨样机进行粉碎后使用Rapid N exceed型快速定氮仪(Elementar,Germany) 采用杜马斯快速定氮法测定各
器官含氮量。各器官干重与其相应氮素含量的乘积即为其氮素积累量,穗部氮素积累量与茎秆氮素积累量的比值即为氮素穗茎比。退化阶段整株和穗的氮积累速率分别为此阶段整株和穗的氮的积累量与持续天数的比值。
1.3.5 产量及其构成因素 成熟期在各试验小区内选取长势均匀一致的区域,划定3 m2(2.0 m,6行)将所有小麦穗由人工剪下自然晒干然后脱粒,风干后称重,并用谷物水分测定仪测定籽粒含水量,调整含水量至12%的籽粒产量,即小区产量 = 实测产量 × (1 - 籽粒含水量) ÷ (1 - 12%)。收获前各小区内选取长势均匀一致的区域连续取50个单穗脱粒后记录穗粒数。于小区测产的风干籽粒中随机取样进行粒重调查,并将数据折算为12%含水量标准的粒重。1.3.6 生育期内气象数据采集与利用 气象资料由位于试验地附近的气象站 (距离试验地约500 m)自动记录。累积生长度日 (growing degree day,GDD,℃ d) 又称为热时间 (thermal time) 的计算公式参考Ferrante等[22]和Zheng等[23]的方法:

式中:Tmax为一天的最高温度 (℃);Tmin为一天的最低温度 (℃);Tb为小麦发育基本温度 (本试验为 0℃)。
1.4 数据处理
试验数据采用Microsoft Excel 2003软件整理数据和图表制作,采用DPS7.05 (Data Processing System,V7.05) 软件分析系统进行统计分析 (Duncan新复极差法)。
2 结果与分析
2.1 播期对冬小麦产量及产量构成因素的影响
由于两生长季播期处理间在单位面积穗数、穗粒数和粒重间的表现规律一致,因此统计了各指标的两年平均值。由表1数据可知,播期显著影响了成熟期冬小麦的单位面积穗数和穗粒数,随播期推迟单位面积穗数呈逐渐降低趋势,穗粒数与之相反。由于单位面积穗数降低的比例与穗粒数提高的比例基本一致,所以两品种的单位面积粒数在处理间均无显著差异。播期对粒重无明显影响,两个品种的粒重在播期处理间基本持平。所以,冬小麦籽粒产量未受播期影响,两品种的籽粒产量在播期处理间均无显著差异。
2.2 播期对冬小麦各生育时期的影响
由播期对冬小麦各生育时期的影响 (表2) 可以看出,两生育季小麦播种至出苗所需天数随播期的推迟而增加。最早播期9月24日处理 (S1) 播种至出苗所需天数为6~7天,10月1日 (S2)、10月8日(S3) 和10月15日 (S4) 处理播种至出苗所需天数为7~8天,而最晚播期10月22日处理 (S5) 播种至出苗所需天数增加至8~9天。随着生育进程的推进,播期处理间生长发育差距逐渐缩短,各播期处理达到拔节期的时间间隔仅相差1天,而到达孕穗期、开花期和成熟期的时间基本一致。
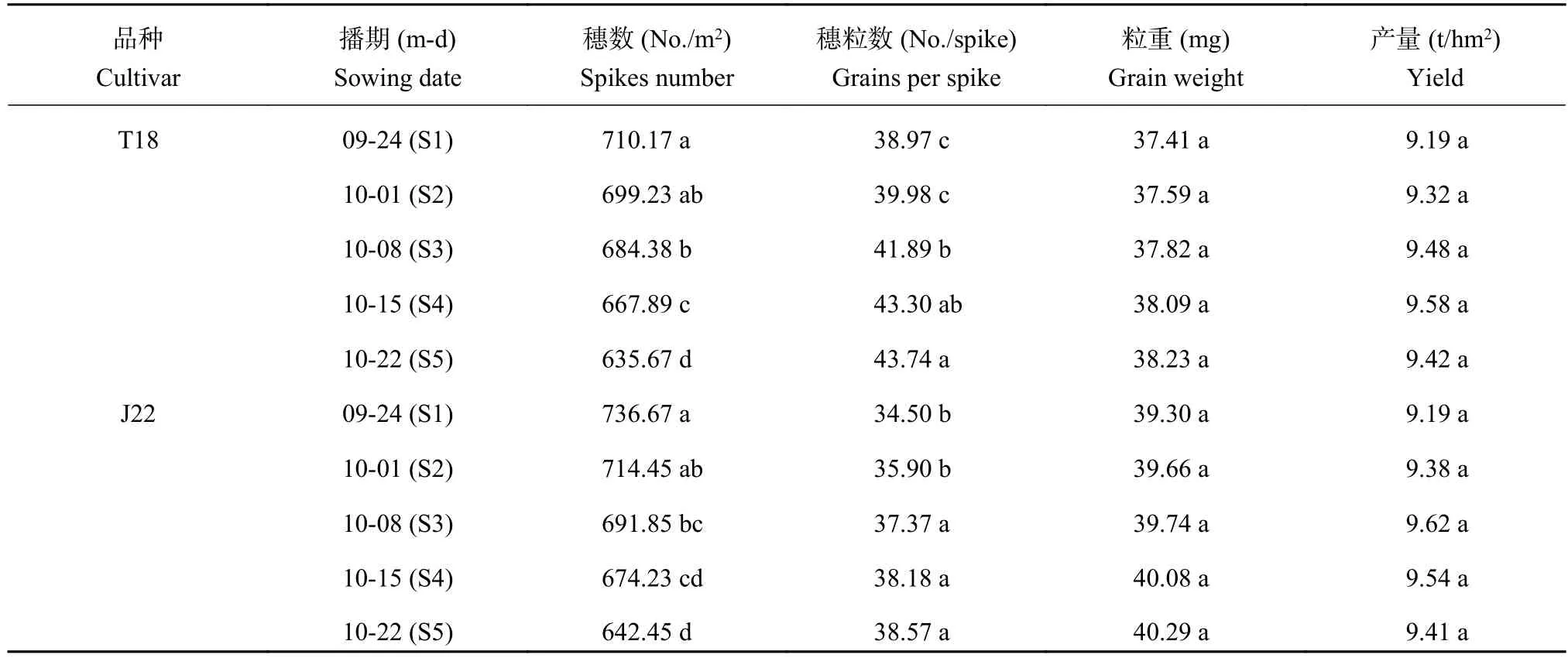
表 1 播期对冬小麦单位面积穗数、穗粒数、粒重和产量的影响Table 1 Effects of sowing date on spikes per square meter, grains per spike, grain weight and yield
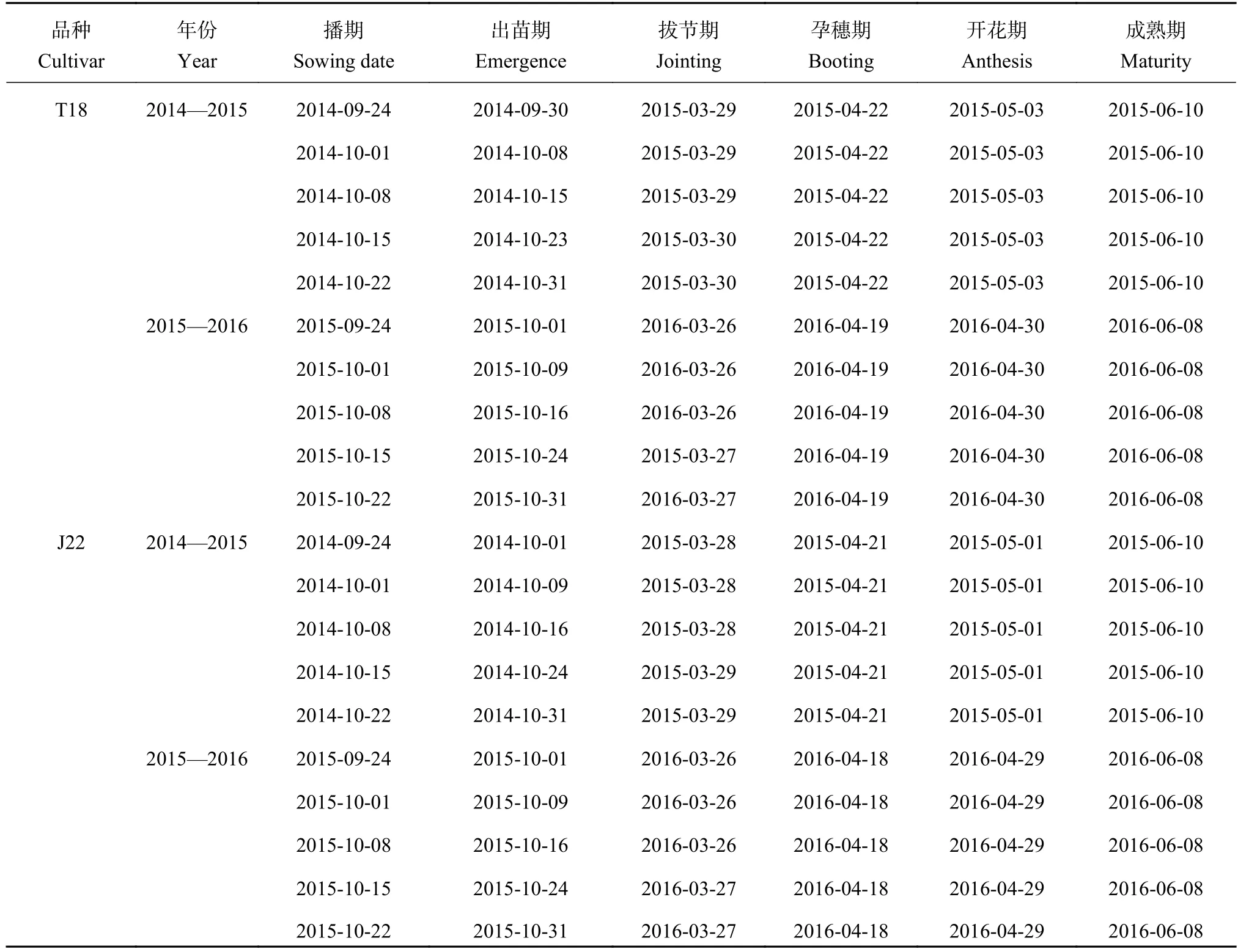
表 2 小麦品种T18和J22达到出苗期、拔节期、孕穗期、开花期和成熟期的日期 (y-m-d)Table 2 The date for emergence, jointing, booting, anthesis and maturity for wheat cultivars of T18 and J22
2.3 播期对冬小麦穗分化和结实特性的影响
由表3可知,两品种的单位面积可孕小花数和结实粒数在处理间均无显著差异。而两品种的单位面积最大分化小花数随播期推迟呈减少趋势,除S1和S2处理间差异不显著外,各处理间差异均达到显著水平。其中,相比S1处理,T18的S3、S4和S5处理单位面积最大分化小花数分别降低11.3%、17.6%和24.4%,J22的S3、S4和S5处理单位面积最大分化小花数分别降低9.0%、15.7%和27.0%。可见,尽管推迟播期显著减少了小麦单位面积最大分化小花数,但是维持了可孕小花数和最终结实粒数,从而为产量的维持提供了有利保障。
播期处理间的小花存活率和小花结实率表现也存在差异 (表3)。小麦小花存活率和小花结实率均随播期的推迟而呈上升趋势,除S1和S2处理差异不显著外,其余各处理间存在不同程度的显著差异。相比S1处理,T18的S3、S4和S5处理小花存活率分别提高17.9%、29.1%和37.3%,J22的S2、S3、S4和S5处理小花存活率分别提高5.4%、12.4%、23.1%和39.8%;T18的S3、S4和S5处理小花结实率分别为17.0%、27.9%和35.7%,J22的S2、S3、S4和S5处理小花结实率分别为5.0%、11.6%、22.0%和38.5%。而可孕花结实率基本维持不变,说明小麦花后可孕花败育程度未受播期影响。由此看来,可孕花数和结实粒数的维持主要由小花存活率的提高实现的。
2.4 适期晚播条件下冬小麦小花原基分化、退化与GDD的关系
由图1可见,在不同播期处理下,随着播后累积生长度日GDD的增加,两个冬小麦品种单位面积小花数量变化依次进入由快至慢的分化阶段和由快速下降到缓慢下降的退化阶段。不同播期处理到达小花开始分化、退化所需GDD差异显著,表现为随播期的推迟小花分化、退化时间提前,所需GDD明显减少。相比S1处理,S2、S3、S4和S5处理到达小花开始分化所需GDD分别减少137.81、253.23、349.12和458.19℃ d (两生育季平均值,下同),到达小花开始退化所需GDD分别减少152.42、261.67、370.54和 492.50℃ d。
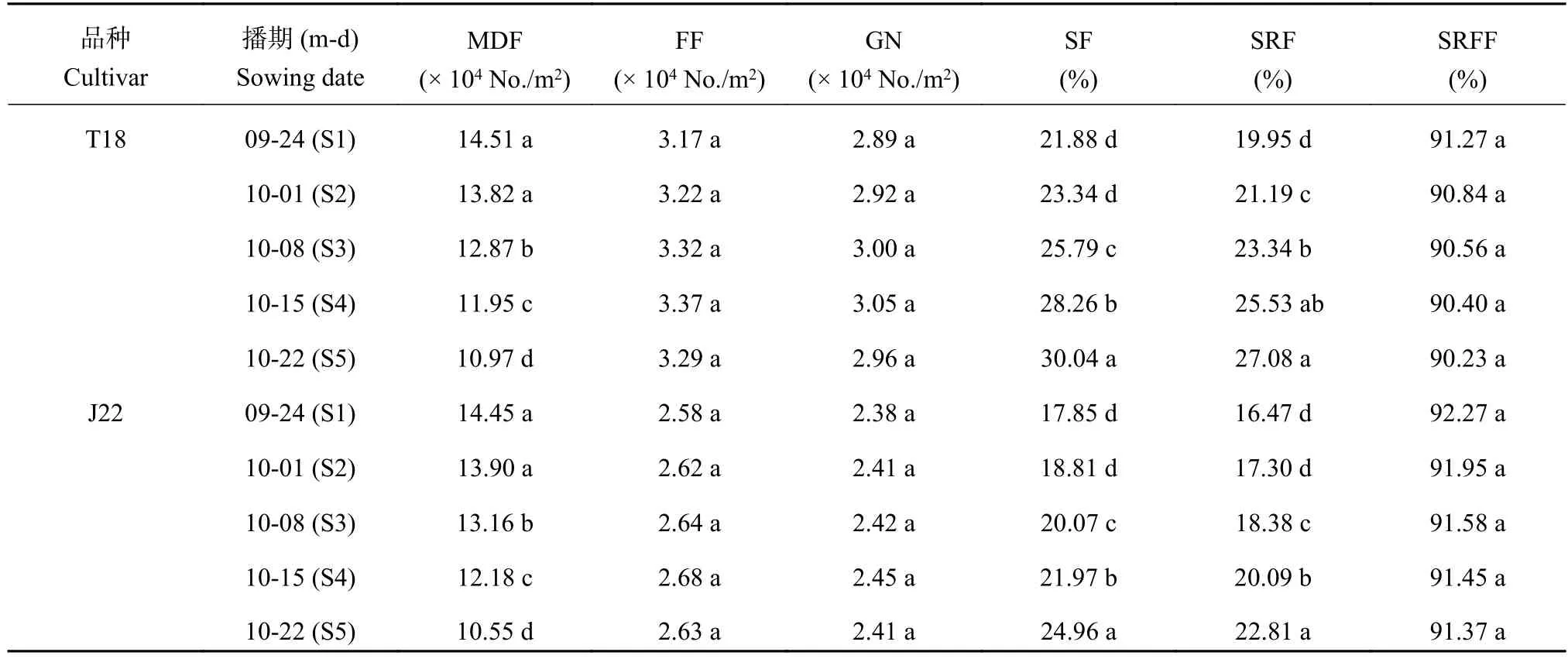
表 3 播期对冬小麦穗分化和结实特性的影响Table 3 Effects of sowing date on the characteristics of floret differentiation and seed setting of winter wheat
2.5 晚播对冬小麦小花分化、退化速率的影响
由冬小麦小花分化、退化速率与GDD的关系(图2)可见,两个品种冬小麦小花分化速率处理间存在显著差异,初始分化速率和最大分化速率均随播期的推迟呈下降趋势,其中T18的S1、S2、S3、S4和S5处理的平均分化速率分别为546.13、531.99、467.35、445.95 和 420.66 个/ (m2·℃ d),J22 的 S1、S2、S3、S4和S5处理的平均分化速率分别为549.22、530.86、468.38、450.21 和 406.31 个/ (m2·℃ d)。在小花退化阶段,处理间的小花退化速率存在明显差异,均表现为随播期推迟而显著降低,其中,T18的S1、S2、S3、S4和S5处理的平均退化速率分别为604.99、556.51、487.44、427.27和368.90个/(m2·℃ d),J22 的 S1、S2、S3、S4 和 S5 处理的平均退化速率分别为639.58、597.32、551.60、493.47和399.88 个/ (m2·℃ d)。总体来看,相比早播处理,推迟播期小麦的群体分化速率有所降低,从而影响了最大分化小花数,但是退化速率的降低能够弥补分化数量的不足,这可能是决定晚播处理小花存活率提高、可孕小花数和最终穗粒数维持的关键因素。
2.6 播期对冬小麦开花期干物质积累量与退化阶段积累速率的影响
由表4可知,两品种开花期的单位面积穗干重虽然随播期的推迟呈上升趋势,但处理间未达显著水平,而茎干重的表现与穗干重相反,随播期的推迟呈下降趋势。两品种的干物重穗茎比随播期推迟呈上升趋势,相比S1处理,T18的S5处理的干物重穗茎比提高了11.8%,J22的S5处理干物重穗茎比提高了12.2%。两品种的小花退化阶段的整株和穗的干物质积累速率均随播期的推迟呈上升趋势,且处理间存在较大差异。表明在小花退化阶段,小麦的光合同化能力随播期推迟明显改善,且向穗部的物质转运能力也相应增强。相比S1处理,两品种的S3、S4和S5处理的整株干物质积累速率平均提高19.2%、26.9%和36.2%;两品种的S5处理的穗干物质积累速率明显提高,平均提高16.1%。
2.7 播期对冬小麦开花期氮素积累量与退化阶段积累速率的影响
两品种开花期的单位面积穗氮积累量和茎氮积累量均表现为随播期的推迟而降低 (表5),且茎氮积累量的下降幅度更加明显。氮积累量穗茎比的变化规律与干物重穗茎比基本一致。相比S1处理,T18的S4和S5处理的氮积累量穗茎比分别提高6.7%和8.2%,J22的S4和S5处理氮积累量穗茎比分别提高7.3%和10.0%。两品种退化阶段的整株和穗的氮积累速率变化规律与干物质基本一致,表明在此阶段小麦的地上部氮素吸收能力随播期的推迟而显著提高。尽管开花期穗部的单位面积氮素积累量随播期的推迟而降低,但在退化阶段穗部的氮素随播期延迟表现出加速积累的趋势。相比S1处理,两品种的S3、S4和S5处理的整株氮积累速率平均提高17.9%、27.0%和34.1%;两品种的S5处理的穗氮素积累速率平均提高11.7%。
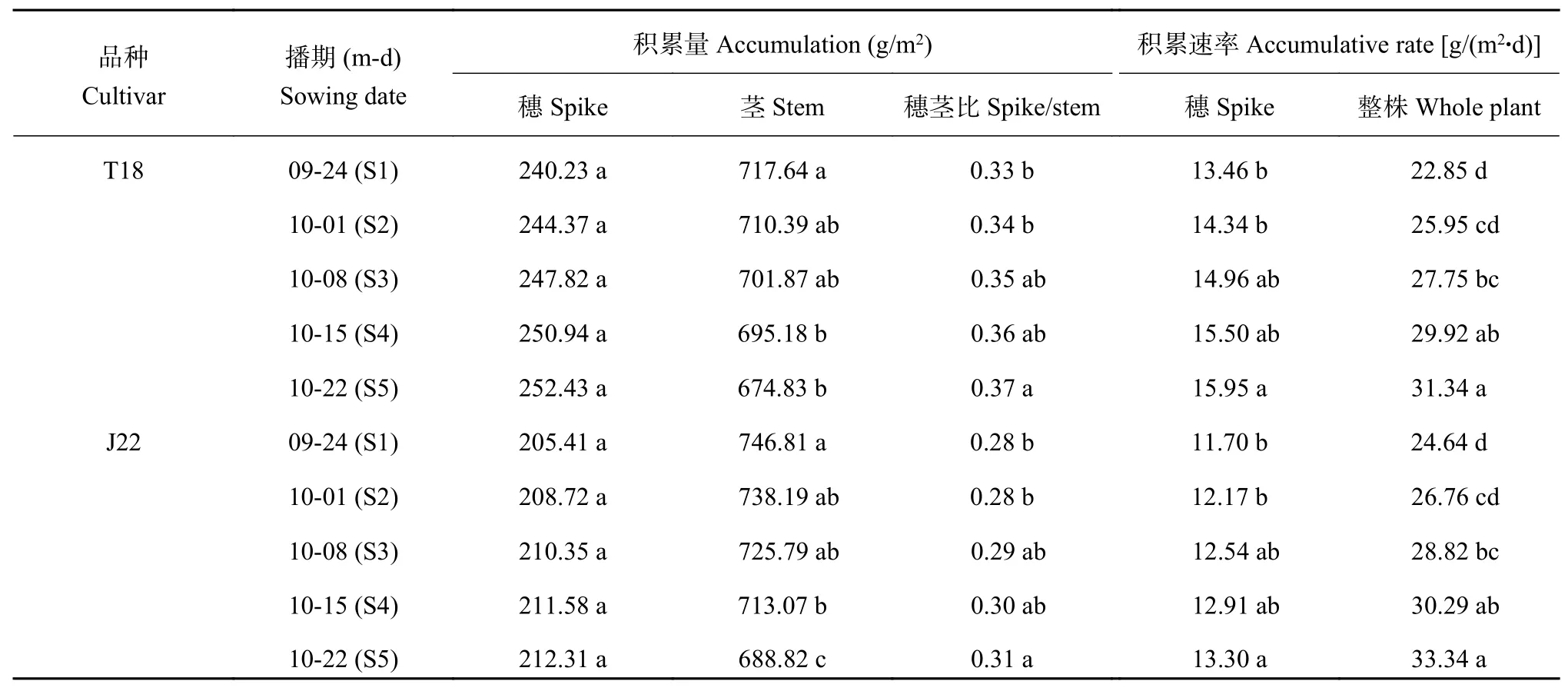
表 4 不同播期小麦开花期不同植株器官单位面积干物质积累量和积累速率Table 4 Accumulation and accumulative rate of dry matter of different plant organs at anthesis stage under different sowing dates
2.8 开花期可孕花数与同化物积累和分配的相关性分析
相关分析结果 (表6) 表明,开花期单位面积可孕花数与穗干物重、干物重和氮积累量穗茎比、整株和穗的干物质和氮的积累速率以及小花存活率均呈极显著正相关;与茎氮素积累量呈极显著负相关;与茎干物重和穗氮素积累量呈负相关,但均未达到显著水平。小花存活率与穗干物重、干物重和氮素积累量穗茎比、整株和穗的干物质积累速率呈极显著正相关;与整株和穗的氮积累速率呈正相关;与茎秆氮积累量呈极显著负相关,茎干重呈负相关;与穗氮素积累量呈负相关,但相关性较小。
2.9 播期对小麦单位面积有效茎蘖和无效分蘖退化小花数的影响
播期推迟显著影响了单位面积有效茎蘖和无效分蘖的退化小花数 (表7),且处理间差异显著,单位面积有效茎蘖和无效分蘖的退化小花数均随播期的推迟呈下降趋势,且无效分蘖退化小花数下降程度更加明显。相比S1处理,T18的S3、S4和S5处理无效分蘖退化小花数分别降低18.4%、29.2%和41.1%,J22的S3、S4和S5处理无效分蘖退化小花数分别降低11.8%、23.8%和47.3%;T18的S3、S4和S5处理有效茎蘖退化小花数分别降低14.6%、21.8%和28.8%,J22的S3、S4和S5处理有效茎蘖退化小花数分别降低12.7%、19.2%和27.4%。推迟播期显著降低了无效分蘖退化小花占总退化小花的比例,可能是由于早播处理无效分蘖占总茎蘖比重高于晚播处理的缘故。
相关分析结果 (图3) 表明,单位面积退化小花数与开花期主茎穗干物重所占单位面积穗干重的比例、主茎穗氮积累量所占单位面积穗氮积累量的比例呈极显著负相关,表明提高主茎穗的比例可以有效降低退化小花数,从而利于稳定可孕花数和提高小花存活率。
2.10 播期对小麦可孕小花和退化小花GDD生产效率的影响
播期显著影响了冬小麦可孕小花和退化小花GDD生产效率 (表7),其中两品种的可孕小花GDD生产效率随播期的推迟呈上升趋势,而退化小花GDD生产效率随播期推迟呈先上升后下降的趋势。相比S1处理,T18的S3、S4和S5处理可孕小花GDD生产效率分别提高20.8%、28.2%和37.6%,J22的S3、S4和S5处理可孕小花GDD生产效率分别提高21.2%、30.8%和38.0%。相比S2处理,T18的S3、S4和S5处理退化小花GDD生产效率分别降低4.2%、9.4%和15.6%,J22的S3、S4和S5处理退化小花GDD生产效率分别降低2.6%、7.5%和19.8%。可见,尽管推迟播期冬小麦花前GDD累计总量减少,总分化小花数和退化小花数降低,但可孕小花GDD生产效率显著提高,表明可孕小花的GDD的利用效率得到明显改善,而降低了退化小花GDD生产效率,从而减少了退化小花对GDD的无效利用。
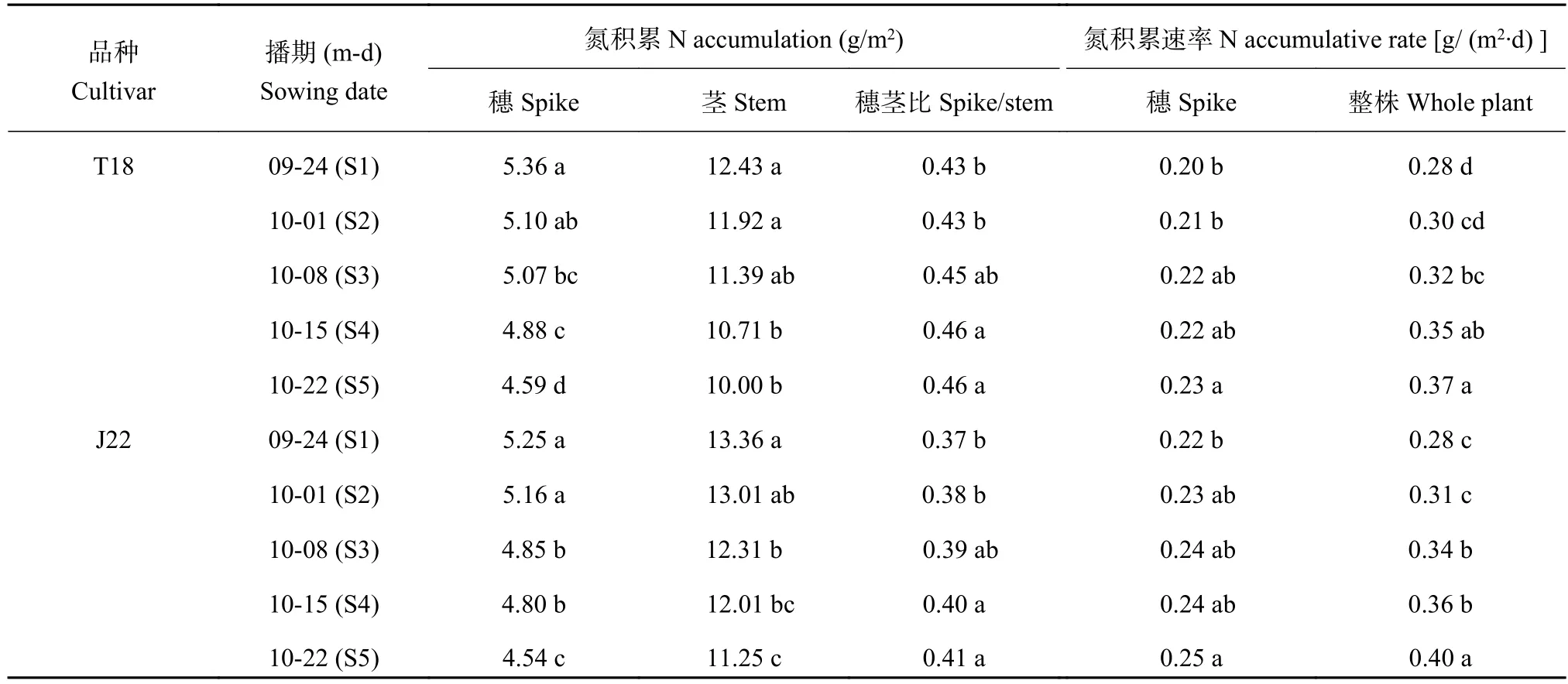
表 5 播期对开花期不同植株器官单位面积氮素积累量和积累速率的影响Table 5 Effects of sowing date on the accumulation and accumulative rate of nitrogen of different plant organs at anthesis
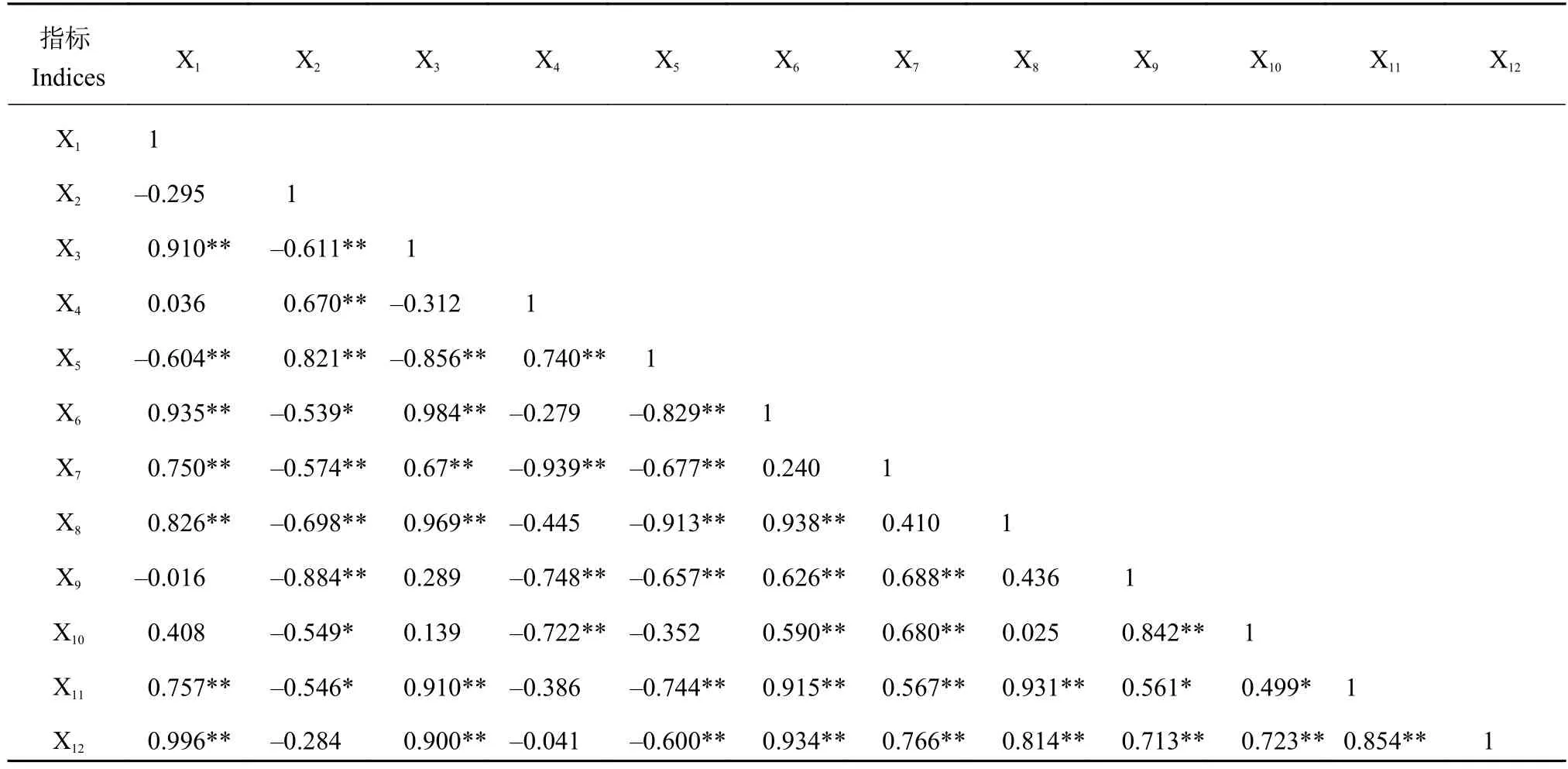
表 6 开花期单位面积可孕花数与同化物积累和分配、干物质和氮积累速率的相关分析Table 6 Correlation between fertile florets number and photosynthetic accumulation and distribution and accumulative rate of dry matter and nitrogen at anthesis stage
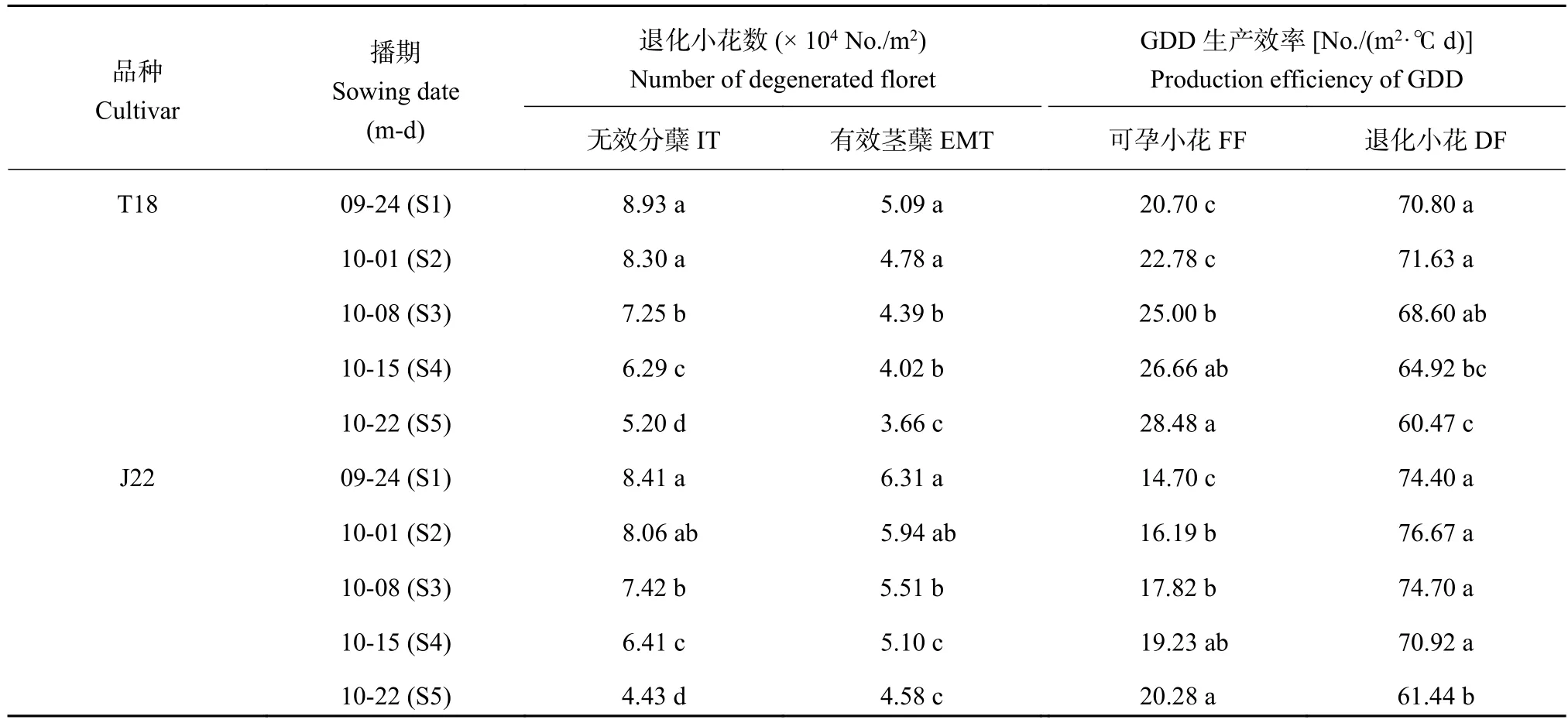
表 7 播期对小麦单位面积无效分蘖和有效茎蘖退化小花数以及可孕小花和退化小花GDD生产效率的影响Table 7 Effect of sowing dates on the number of degenerated florets in ineffective tillers (IT) and effective main shoots and tillers (EMT) and the GDD production efficiency of fertile florets (FF) and degenerated florets (DF) of winter wheat
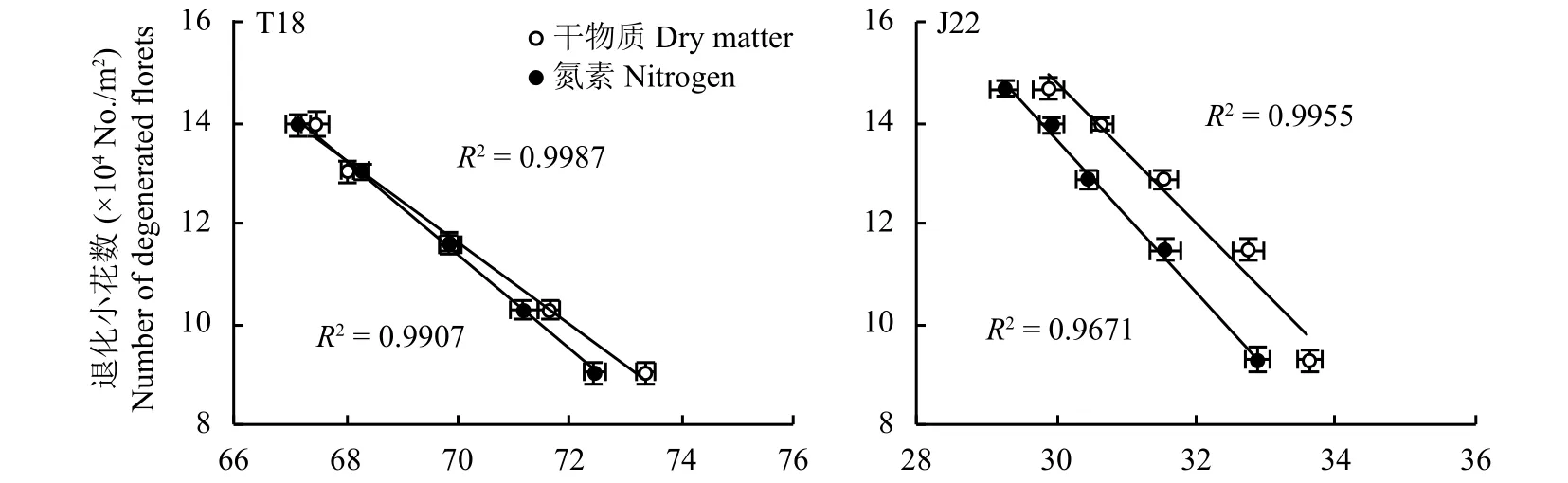
图 3 开花期同化物积累和分配与单位面积退化小花数的相关分析Fig. 3 Correlation analysis between photosynthate accumulation and distribution and the number of degenerated florets per square meter at anthesis
3 讨论
“绿色革命”以来,小麦产量的大幅提高主要得益于单位面积粒数的增加[6,17]。提高单位面积粒数也成为小麦高产栽培和育种研究的重要目标[24]。因此,了解单位面积粒数的变化对于解释小麦产量变化和进一步挖掘小麦产量潜力至关重要。穗粒数的形成是小花分化、发育、退化和结实等一系列生理过程的最终体现[12]。可孕小花数与穗粒数密切相关,而开花前的幼穗分化发育质量决定了可孕小花数目[4]。本研究结果表明,在推迟播期条件下单位面积可孕小花数与最终粒数基本相等,可孕花结实率也未发生较明显变化,这说明单位面积粒数主要由开花期可孕花数决定的。由于小花结实率是小花存活率和可孕花结实率的乘积,在可孕花结实率恒定的前提下,小花存活的比例决定了小花结实率的高低。因此,在最大分化小花数减少的情况下,小花存活率的提高成为单位面积可孕花数和粒数维持的关键。有研究表明,小麦穗粒数的多少除受分化的小花数影响外,还取决于小花的退化程度[25]。从小花分化和退化速率动态来看,推迟播期导致小花分化速率降低的同时小花退化速率也得到了明显减少,最终退化小花数的减少弥补了小花分化数量的不足,从而保证了单位面积可孕花数的维持。
养分的平衡供应是作物生长的基础,是影响作物形态发育、器官建成和产量形成最重要的因素[26]。前人研究证明,开花期单位面积穗干重与可孕小花数和籽粒数目呈显著正相关[11,19],表明可孕花数存活的数量受开花前分配给穗部的同化物供给量的调控。由于开花前幼穗和茎并进的快速生长对同化物存在激烈竞争,由此认为小花的退化也与同化物在茎与穗中的分配有关[13]。本研究发现,开花期可孕花数和小花存活率与开花期穗干重、开花期干物质穗茎比均呈极显著正相关,这表明开花期可孕花数的存活数量不仅与穗器官干物质供给总量有关,而且与干物质在穗部和茎秆的分配比例密切相关。而开花期可孕花数与穗部氮素积累总量关系不显著,与氮素穗茎比呈极显著正相关,这说明穗部氮素积累总量对于花期可孕小花数的形成的影响力较小,而氮素在穗部和茎秆的分配比例对于可孕花数的存活作用更加明显。因此,可孕花数的维持和存活率的提高也取决于养分供应在幼穗和茎秆之间的良好的协调分配。此外,植株和穗部在小花退化过程中的干物质、氮素积累速率均随播期推迟而升高,而且与小花存活率、开花期可孕花数均展示了显著的正相关关系,说明小花退化过程中地上部整株的干物质和氮素积累速率的加快有利于较多的干物质和氮素输入幼穗中,进而为可孕小花的发育形成提供了良好的物质基础。这也表明穗粒数不仅与小花退化阶段幼穗的增长速率相关,与植株的生长速率也密切相关。研究表明,植株内源激素在调控同化物分配以及韧皮部运输与卸载方面起着重要的作用[27-28]。由此推测,植株内源激素参与调控了不同播期条件下小花发育过程中植株不同部位同化物的供应与分配,同时也对植株和穗部生长速率产生了影响,其内在的生理机制需进行进一步的探究。
适宜的群体结构是作物获得高产的基础[29]。小麦群体由主茎和分蘖共同构建,分蘖的发生、生长发育和存活消亡对麦田的群体结构和产量起重要的调节作用[30-31]。由于开花前穗部小花分化、发育以及退化等一系列生理过程的发生伴随着分蘖群体的消减,因此消亡的分蘖数量直接影响单位面积分化小花和退化小花的数量。本试验条件下单位面积有效茎蘖和无效分蘖所产生的退化小花数均随播期的推迟而减少,且无效分蘖产生的退化小花数下降幅度更加明显,这归因于推迟播期未形成过多的无效分蘖以致造成无效消耗。Guo和Schnurbusch[32]研究表明,去除分蘖降低了分蘖与主茎的资源竞争,使更多的资源用于主茎中穗和茎秆的生长,有利于缓和穗和茎秆之间的竞争,从而将更多地资源用于小花发育。也有研究指出增加总穗数中主茎穗的比重,有利于提高或稳定小麦产量[33-34]。本试验结果表明,开花期主茎穗的干物重和氮素积累在单位面积群体总穗中的比重与退化小花总数呈显著负相关,这说明提高主茎在群体中同化物所占比例能够有效维持单位面积可孕花数和减少退化小花数,从而降低了小花死亡消耗。适宜的播期有利于构建良好群体结构、优化群体生理指标,以充分利用生态环境中的光温资源[20,35-36]。随着播期的推迟,小花分化所需GDD明显减少,开花前GDD可孕花生产效率明显提升,而GDD退化小花生产效率显著下降,这跟无效分蘖的减少密不可分。由此来看,推迟播期提高了主茎在群体中的比重,减少了无效分蘖对资源的无效耗费,利于优化群体结构和提高资源利用效率。因此,在适期延播条件下适当增加基本苗,提高主茎成穗,有利于增穗增粒,是作为进一步提升产量的有效途径。
4 结论
本研究通过比较播期推迟条件下冬小麦小花发育特性及同化物分配变化上的差异,结果表明小花退化速率缓慢、可孕花存活率较高是晚播处理小麦获得单位面积粒数维持的重要原因。开花期单位面积可孕花数除与穗器官干物质积累量呈正相关外,还与干物质和氮素在穗部和茎秆的分配比例密切相关。此外,小花退化阶段幼穗的增长速率与植株的生长速率也是影响可孕花存活的重要因素。提高主茎穗在单位面积总穗中的比例可以有效减少退化小花数,尤其以无效分蘖产生的退化小花数下降更为明显。推迟播期明显提高了可孕花数的GDD生产效率,而降低了无效小花的GDD生产效率。
猜你喜欢
中国农学通报(2022年29期)2022-11-25
烟草科技(2022年9期)2022-09-24
作物学报(2022年3期)2022-01-22
麦类作物学报(2021年12期)2022-01-08
中国土壤与肥料(2021年5期)2021-12-02
中国土壤与肥料(2021年5期)2021-12-02
山西农业科学(2021年4期)2021-04-19
江苏农业科学(2020年22期)2020-03-03
塔里木大学学报(2019年4期)2020-01-03
现代农业科技(2018年22期)2018-01-15
