
DCC基因对裸鼠皮下结直肠肿瘤的生长和淋巴管生成的影响
2019-04-27 02:23套格苏姜洪伟
中国实验诊断学 2019年4期
套格苏,王 举,胡 琦,姜洪伟*
(1.内蒙古医科大学研究生学院,内蒙古 呼和浩特010110;2.内蒙古自治区人民医院 胃肠外科)
DCC基因是由Fearon等[1]研究结直肠肿瘤时用定位克隆的方法确定并命名的一个肿瘤抑制基因,其编码产物的氨基酸序列与神经细胞黏附分子有明显的同源性。研究发现DCC转录物在结直肠肿瘤组织中显著降低,在增殖性息肉上皮细胞中DCC蛋白表达十分明显,而在被认为是结直肠癌前体的具有明显不典型增生的晚期腺瘤中却未见明显DCC表达[2]。还有学者认为DCC基因失活可能是影响大肠癌预后的重要因素之一,肝转移者发生DCC表达失活的主要机制是发生了基因突变[3]。本研究通过获取基因片段,将目的基因插入到表达载体,并将人结直肠癌SW1116细胞悬液注射到裸鼠皮下,使裸鼠成瘤后进行相关的干预,再对相关指标进行测量,最后进行进一步分析。
1 材料与方法
1.1 实验材料
实验动物BALB/C雄性裸鼠15只购自上海斯莱克实验动物有限公司。动物品系为无特定病原体BALB/C裸鼠,雄性,6-8周。聚合酶链反应(PCR)管、PCR仪等。
1.2 实验方法
①通过基因合成方法获得带有DCC功能区的亚克隆质粒,通过金唯智基因合成获得目的片段。将目的基因插入到表达载体,即将提取的质粒pUC57-DCC与载体pCDNA3.1通过HindIII和XbaI双酶切系统进行酶切链接。转化到大肠杆菌,小量提取质粒。酶切鉴定并大量提取质粒,将提取的两个质粒进行酶切鉴定。②人结直肠癌SW1116细胞培养:DMEM培养基+10%胎牛血清+1%青霉素-链霉素溶液,37℃5%CO2恒温恒湿的细胞培养箱中培养。对已经长满的细胞,弃去培养基,用磷酸缓冲盐溶液漂洗一次,加入胰酶消化液,37 ℃孵育,待细胞完全脱落,加完全培养基中和胰酶,离心6 min(1 000 r/min,离心半径为10 cm),弃上清。无血清培养基洗2次,加5 ml无血清培养基悬浮细胞,计数。用无血清培养基将细胞浓度调至1×107个/ml。③将裸鼠在无特定病原体动物房饲养,自由饮食和饮水,适应性饲养7 d。第7天将SW1116细胞悬液注射到裸鼠皮下,每只小鼠注射0.2 ml。④建模第15天,观察裸鼠成瘤情况,测量肿瘤大小。将裸鼠随机分为3组(15只体重、年龄相等雄性裸鼠),对裸鼠进行如下干预实验:第1组:生理盐水组(n=5,腹腔注射生理盐水);第2组:DCC干预治疗组(n=5,瘤体内多点注射DCC质粒,注射50 μg,质粒浓度908 ng/μl);第3组:5-氟尿嘧啶(5-FU)治疗组(20 mg/kg体重,腹腔注射,连续15 d)。⑤经上述干预处理后,每周用游标卡尺测量小鼠肿瘤的长与宽,称量裸鼠体重,计算抑制瘤率;造模第30天处死裸鼠[由于给药后第15天瘤体无明显差异,因此多饲养了1周,即给药后第22天(造模后第30天)取样检测],剥离肿瘤,称重,拍照,计算抑瘤率。抑瘤率=(1-实验组平均瘤重/对照组平均瘤重)×100%。
1.3 检测相关指标
1.3.1细胞凋亡检测 取1-2 mm大小的肿瘤组织,然后加入5-10倍体积的胰酶,37 ℃消化7 min,用5 ml含10%胎牛血清的培养液终止消化,过滤得到细胞悬液,将细胞悬液离心6 min(1 000转r/min,离心半径为10 cm),得到肿瘤单细胞。后将细胞用磷酸缓冲盐溶液洗1次后,重悬于1×结合缓冲液(Binding Buffer)中(将10×Binding Buffer用蒸馏水稀释)。其中对照用400 μl 1×Binding Buffer,其他处理组用100 μl 1×Binding Buffer。每管加5 μl 磷脂结合蛋白(FITC-Annexin V)和5 μl碘化丙啶(50 μg/ml)。轻柔弹起混匀,室温(25 ℃)避光孵育15 min。加400 μl 1×Binding Buffer至每管中,混匀。1 h内上流式分析软件分析实验数据。
1.3.2Western blotting检测 ①蛋白样品的处理与定量:用BCA法检测细胞裂解液的蛋白浓度,测好浓度的蛋白分装放于-80℃保存。②聚丙烯酰胺凝胶电泳(SDS-PAGE)。
1.3.3淋巴管计数标准 参照Weidner法进行淋巴管计数,在低倍显微镜(×100)下观察全切片淋巴管分布情况后,选择淋巴管密集的3个区域,以高倍显微镜(×200)计数每个区域的淋巴管数量,以3个区域平均值为该标本的淋巴管密度。
1.4 统计学方法
所有数据应用SPSS 22.0进行统计分析。计量资料符合正态分布,用均数±标准差表示,两组间比较采用独立样本t检验,多组间比较采用方差分析,进一步两两比较采用LSD法。计数资料以例数和百分率表示,采用χ2检验或Fisher确切概率法。检验水准α=0.05。
2 结果
2.1 目的基因插入到表达载体
将提取的质粒pUC57-DCC和载体pCDNA3.1通过HindIII和XbaI双酶切系统进行酶切。双酶切鉴定可观察到在300 bp处有特异性条带。双酶切和质粒测序结果表明pCDNA3.1-DCC中含有序列正确的DCC基因。见图1、图2。
2.2 对裸鼠瘤重的影响
生理盐水组、DCC干预治疗组、5-FU治疗组裸鼠的肿瘤重量分别为(0.353±0.119)g、(0.123±0.015)g、(0.147±0.041)g,差异具有统计学意义(F=8.903,P=0.016);进一步两两比较结果显示,与生理盐水组相比,DCC干预治疗组、5-FU治疗组裸鼠肿瘤重量均有所减轻,差异均具有统计学意义(P<0.05),但DCC干预治疗组与5-FU治疗组相比,瘤重差异无统计学意义(P>0.05)。
2.3 细胞凋亡实验结果
3组不同干预措施对裸鼠肿瘤细胞凋亡率的影响,经χ2检验结果显示,3组裸鼠的肿瘤细胞凋亡率差异有统计学意义(χ2=21.362,P=0.000)。与生理盐水组相比,DCC干预治疗组、5-FU治疗组总凋亡率均升高,差异具有统计学意义(P<0.05),见表1。
2.4 转染后DCC蛋白的表达
转染后,Western blotting检测可以发现,曝光900秒扫膜未检测到条带,已重复3次,均未检测到DCC蛋白,见图3。可能与DCC蛋白的表达量特别低,信号较弱有关。
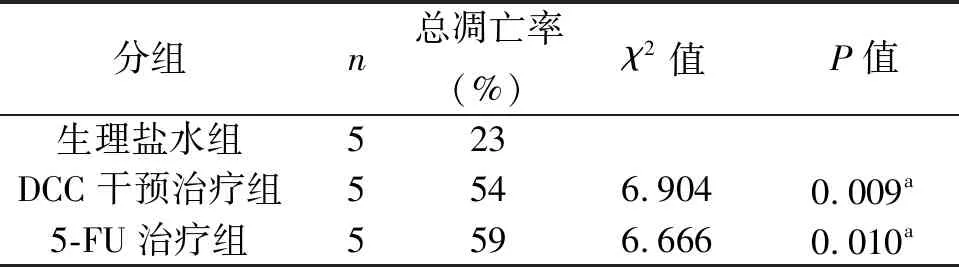
表1 3组不同干预措施对肿瘤细胞凋亡率的影响(%)
注:5-FU为5-氟尿嘧啶;a为与生理盐水组比较,P<0.05
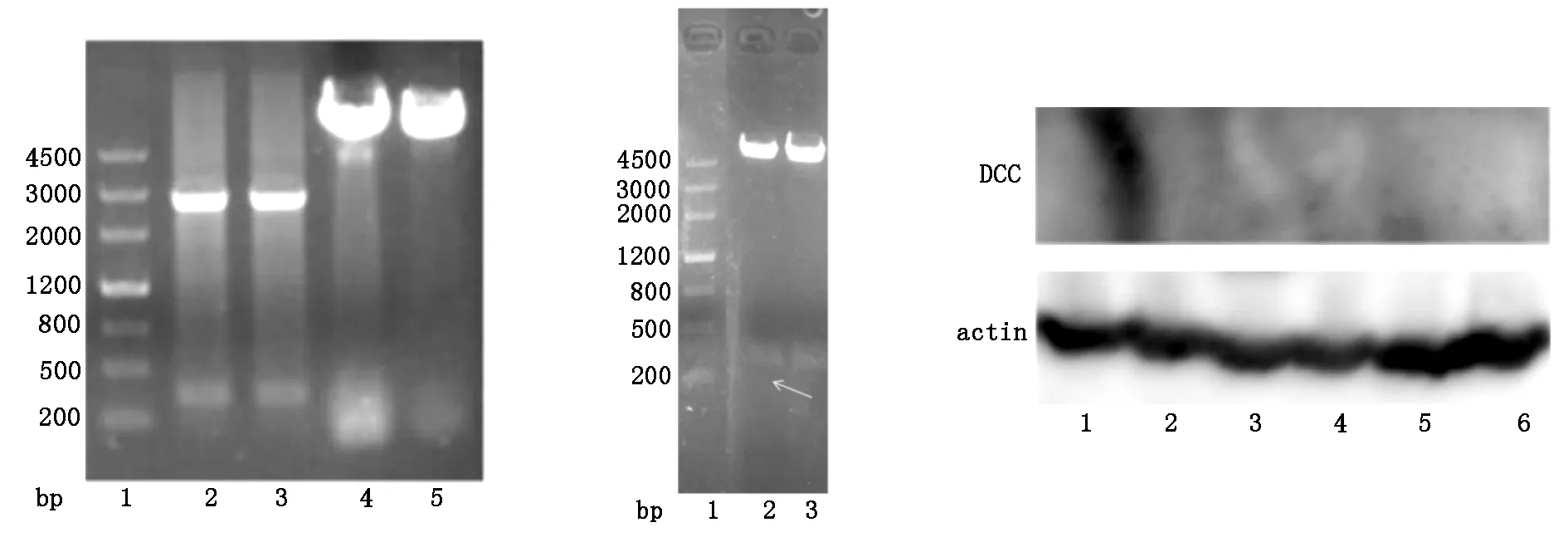
注:1为Marker;2、3为pUC57-DCC;4、5 注:1为Marker;2、3为pCDNA3.1-DCC为pCDNA3.1 注:1、2为生理盐水组;3、4为DCC干预治疗组;5、6为5-FU治疗组;5-FU为5-氟尿嘧啶
图1 pUC57-DCC和pCDNA3.1(+)图2 质粒酶切鉴定电泳图 图3 Western blotting检测转染后DCC蛋白的表达酶切电泳图
生理盐水组、DCC干预治疗组、5-FU治疗组的DCC蛋白表达分别为1.556±0.088、2.061±0.075和2.052±0.114,差异有统计学意义(F=28.427,P=0.001)。与生理盐水组相比,DCC干预治疗组、5-FU治疗组的DCC蛋白表达量均增加(均P<0.001)。
2.5 淋巴管计数结果
生理盐水组、DCC干预治疗组、5-FU治疗组的淋巴管计数情况分别是(14.67±0.626)个、(8.43±1.002)个、(7.49±0.514)个,差异有统计学意义(F=6.489,P=0.039)。与生理盐水组相比,DCC干预治疗组、5-FU治疗组裸鼠的淋巴管数量均减少,差异具有统计学意义(均P<0.05)。
3 讨论
DCC基因位于人体染色体18q21.3,含有1.4 Mbp和29个外显子。其表达蛋白是I型跨膜蛋白,由1 447个氨基酸组成,相对分子质量为190 000。DCC蛋白参与细胞之间、细胞与基质之间的相互作用,使细胞维持正常的生长和分化方式。当DCC基因发生缺失和突变后,DCC蛋白生成障碍,细胞的生长和分化紊乱促使细胞朝着恶性方向演变。DCC基因在绝大多数正常组织中均有表达,但在大多数结直肠癌组织中的表达却有下调趋势。DCC基因抑癌的机制与其能够诱导细胞凋亡有关。作为依赖性受体,当配体Netrin-1存在时,DCC与其结合对细胞凋亡起到抑制作用,但当细胞缺乏Netrin-1配体时,DCC则诱导细胞凋亡。
DCC基因的胞内功能域(3 727-3 792 bp)具有诱导结肠癌细胞凋亡的作用[4]。DCC基因改变是癌变发生过程中的早期基因事件,其缺失或失活是肿瘤发生发展的一个促发因素。DCC蛋白的表达缺失与肿瘤浸润转移及细胞分化相关[5]。研究者发现大肠癌中DCC表达缺失频率较高,可能通过上调Bcl-2蛋白表达和下调Bax蛋白表达阻止细胞凋亡,从而促进大肠癌的发生[6]。
Itoh等[7]研究了人结直肠癌中DCC mRNA的表达量,结果显示与非癌组织相比,30例结直肠癌中的17例存在DCC mRNA低表达,且结直肠癌肝转移的4个标本均表现为DCC mRNA的表达水平下降,该研究结果表明,DCC的功能性丧失,有可能是癌症转移的重要机制之一。
DCC基因的表达缺失可作为判断大肠癌转移潜能及预后的独立指标[8]。大肠癌组织中确实存在DCC蛋白表达缺失,且与大肠癌患者Dukes分期有关;大肠癌组织中存在高频率DCC基因启动子甲基化的发生,且与直肠癌的关系更为密切;DCC基因启动子甲基化可能是大肠癌中DCC失表达的主要机制之一。转染的DCC基因可以抑制细胞增殖,导致癌胚抗原在SW1116细胞中表达下调,从而降低其浸润和转移能力[9]。
肿瘤抑制基因的存在主要是由3个方面提出的:一是通过与正常细胞杂交抑制肿瘤细胞的转化表型;二是多种肿瘤的染色体缺失;三是肿瘤细胞特异染色体区域杂合性缺失。许多研究结果支持多种肿瘤抑制基因的剂量丢失或改变在广泛的癌症发生和(或)进展中起关键作用[10]。在中枢神经系统中,DCC表达较高,是轴突导向和神经元迁移的关键因子。最近,细胞培养实验表明,Netrin-1和DCC是体细胞重编程的调节器[11]。
DCC诱导细胞凋亡的丧失也与结直肠肿瘤的数量和侵袭性有关,在诱发抑癌基因突变的背景下,导致高度浸润性腺癌的发展。这些研究表明,DCC作为一个肿瘤抑制剂可通过其能力触发肿瘤细胞凋亡[12]。
本研究采用PCDNA3.1(+)-DCC重组质粒转染15例皮下结直肠肿瘤裸鼠后,检测了DCC蛋白表达情况,结果表明DCC蛋白表达量降低,在3次Western blotting检测中均未发现条带(DCC蛋白),说明在结直肠癌中DCC基因失表达。此外,癌组织中测得淋巴管的数量也显著减少。更有说服力的指标是DCC干预治疗组裸鼠的瘤重明显小于生理盐水组。研究结果还显示,与生理盐水组比较,DCC组总凋亡率有升高,说明DCC基因可诱导细胞凋亡,并可通过诱导细胞凋亡来抑制裸鼠皮下结直肠癌癌细胞的生长和增殖。
综上,DCC基因能够抑制结直肠肿瘤的生长和淋巴管的生成,且抑癌机制与其能够诱导细胞凋亡有关。目前对DCC基因的研究有了很大的进步,但是关于DCC基因的失活机制、DCC基因与其他基因是如何协同作用于结直肠癌的发生、发展过程,如何诱导细胞分化等有待进一步研究。
猜你喜欢
微循环学杂志(2022年2期)2022-12-28
黑龙江医药(2022年18期)2022-10-13
疯狂英语·初中天地(2022年5期)2022-07-06
疯狂英语·初中版(2022年5期)2022-05-11
组织工程与重建外科杂志(2022年1期)2022-03-05
外科研究与新技术(2021年1期)2021-05-20
世界科学技术-中医药现代化(2020年2期)2020-07-25
中国临床医学(2019年2期)2019-05-21
医学信息(2016年31期)2017-02-27
腹腔镜外科杂志(2016年11期)2016-06-01
