
互花米草入侵对鸟类的生态影响
2019-05-13 07:06朱晓静鲁长虎
生态学报 2019年7期
陈 潘,张 燕,朱晓静,鲁长虎,*
1 南京林业大学生物与环境学院,南京 210037 2 安徽师范大学环境科学与工程学院,芜湖 241000
生物入侵严重威胁全球生物多样性[1- 2],直接或间接影响本地生物类群以及生态系统的功能和服务,是目前导致生物多样性丧失的最主要原因之一[3- 4]。入侵植物可以强烈的改变生态系统中的生物和非生物特征[5],影响本地植被、昆虫、哺乳类、鸟类等许多生物类群[6]。入侵不仅改变本地植被群落影响鸟类栖息地的组成,也通过改变食物网结构影响鸟类的种群数量和物种多样性[7],入侵植物对鸟类的生态影响正受到越来越多的研究关注[8]。
互花米草(Spartinaalterniflora)隶属禾本科(Gramineae)米草属(Spartina),为多年生草本植物,原产北美洲大西洋沿岸和墨西哥湾,由于其耐盐、耐淹和促淤护堤作用,曾被世界多地引种,用于改善沿海生态环境[9]。但互花米草往往快速增殖取代本地植被,改变原有生态系统的结构和功能,同时影响鸟类等多种生物类群。互花米草入侵沿海湿地导致的鸟类栖息地退化和生态影响在世界范围内引起了长期、广泛的关注,例如:新西兰[10]、英国南部海岸河口[11]、澳大利亚[12- 13]、美国旧金山海湾[14- 15]、西班牙[16]等地均有相关研究报告。互花米草自20世纪70年代引入中国,在过去的几十年其分布面积快速扩散,沿海各省从辽宁到广西均有分布[17]。其中,近94%的互花米草分布在江苏、上海、浙江和福建的沿海湿地,对本地土壤地貌、生物类群、生态系统功能等造成严重影响[18],已成为我国受关注度最高的入侵物种之一[19]。我国东部沿海湿地是许多珍稀水鸟的重要栖息地,也是东亚-澳大利亚迁徙线路候鸟的重要停歇点[20],分布多个国家级自然保护区,例如:盐城湿地珍禽国家级自然保护区、崇明东滩鸟类国家级自然保护区等。目前,互花米草已在多个保护区内扩散形成优势植被群落,对鸟类群落造成多种生态影响[21- 23]。本文系统梳理了国内外关于互花米草入侵对鸟类生态影响的研究进展,分析了入侵对鸟类栖息地、食物资源、繁殖、群落等方面的抑制和促进作用,对未来研究方向以及互花米草的清除治理提出了建议。
1 互花米草入侵改变鸟类栖息地格局
植被群落是鸟类栖息地的重要组成部分,是构建鸟类合适栖息地的基础[24]。由于互花米草较强的适应性及温和的入侵地环境条件,入侵后常常能够快速增殖,在竞争中取代本地物种,在潮滩形成单优势植物群落,改变本地植被群落类型和分布[25]。互花米草入侵对植被群落的影响在一定程度上也改变了沿海滩涂鸟类栖息地景观格局[26- 27]。
米草属植物对沿海滩涂的侵占,在世界范围内引起依赖滩涂生境鸟类栖息地的改变。英国威尔士戴菲河大米草(Spartinaanglica)仅增加10%的面积,就已显著改变鸟类栖息地结构,造成黑腹滨鹬(Calidrisalpina)、剑鸻(Charadriushiaticula)、蛎鹬(Haematopusostralegus)等数量明显下降[28]。英国南部海岸河口大米草入侵光滩面积的增加导致鸻鹬类的可利用觅食地面积减少[11]。美国太平洋沿岸互花米草在中低潮滩的扩张导致涉禽觅食地的丧失,迁徙鸟类被迫改变觅食地选择[29- 30]。互花米草入侵美国威拉帕海湾形成高密度单一化的植被群落,严重限制了滨鸟的栖息地面积,使水鸟越冬和繁殖关键生境减少了16%—20%,导致鸻鹬类在光滩的觅食时间减少了50%[31]。对美国旧金山海湾湿地鸟类的环境容纳量分析发现,互花米草扩张使鸟类的栖息地面积减少了33%[32]。虽然美国上个世纪利用疏浚弃土种植互花米草建造的人工沼泽为一些鸟类提供了栖息地,但因理化特征的差异导致鸟类群落结构组成明显与自然湿地不同,不能完全替代自然湿地功能[33]。
遥感监测显示,互花米草入侵导致我国沿海红树林、潮滩芦苇(Phragmitesaustralis)、碱蓬(Suaedaglauca)等鸟类适宜生境面积锐减[17],造成沼泽湿地中一些专性和濒危鸟类栖息地丧失[34]。互花米草入侵显著地将泥滩转变为草地,改变了栖息地的景观格局,研究显示绝大多数鸟类选择泥滩或芦苇生境,而被互花米草入侵后的泥滩和芦苇生境可利用性降低,不利于鸟类的栖息[21]。在我国江苏盐城国家级自然保护区,互花米草入侵改变了植被群落格局,严重阻碍了丹顶鹤(Grusjaponensis)的活动,降低了该区域作为丹顶鹤栖息地的价值[27]。对互花米草的清除实验表明,治理后的恢复生境可为雁鸭类等重要水鸟提供良好的栖息地,生物多样性维持的生态服务功能有了明显的恢复[35]。西班牙奥迪埃尔沼泽在清除入侵植物密花米草(Spartinadensiflora)并栽种本地植被的两年后,鸟类多样性和栖息地质量得到显著提高[16]。
但也有一些研究认为互花米草入侵可能促进了生态系统的演化,为一些鸟类提供了合适栖息地[15,23]。对崇明东滩互花米草入侵的长期监测显示,互花米草种群在一个5年的高速增长期后会有一个更长时间的衰退。互花米草会促使滩面淤高造成潮水量减少,导致互花米草退化和死亡,这又为本地碱蓬、芦苇的生长提供了演替条件[36]。互花米草还可以在其他植被不宜生长的光滩生长,作为先锋植物为红树林等其他植被的演替创造条件[37]。美国威拉帕海湾的研究显示,在低潮滩互花米草往年枯死的植株会被潮水冲刷掉,且春季新生植株相对比较稀疏,为鸟类提供了可利用的栖息地,春季迁徙期一些鸟类会在互花米草群落中觅食,并利用互花米草群落作为隐蔽所躲避猛禽等天敌的捕食[38]。有学者认为,我国沿海光滩新形成的互花米草群落可以为一些鸟类提供觅食、筑巢等栖息地[39- 40]。近年有研究认为入侵植物与本地生态系统各类动植物间存在长期的相互作用,对入侵植物的快速清除有时可能对生态系统带来副作用,反而危及部分鸟类的栖息[41]。一些鸟类已经适应入侵互花米草对自然生境的改造,例如入侵互花米草群落是加州长嘴秧鸡(Ralluslongirostris)的重要栖息地[42],在崇明东滩对斑背大尾莺(Megaluruspryeri)的研究也证实了这一观点[23]。
2 互花米草入侵对鸟类食物资源的影响
外来植物的入侵不仅改变本地植被群落,也通过改变食物链影响着鸟类[22]。作为较高营养级,许多盐沼鸟类依靠本地植物种子、块茎以及昆虫和大型底栖动物作为食物来源[21]。互花米草入侵导致植被、昆虫、大型底栖动物的分布和多样性发生变化[43- 45],这可能间接影响到鸟类的可利用食物资源和觅食行为。
2.1 对植物性食物的影响
本地优势植被的种子、块茎是一些鸟类的重要食物资源[43]。入侵的互花米草竞争替代了本地优势植被,减少了鸟类的食物资源。研究显示,雁鸭类、小天鹅和白头鹤等大型鸟类均以海三棱藨草(Scirpusmariqueter)等本地植被的球茎为食,并不采食互花米草,因此互花米草入侵降低了栖息地对鸟类群落的承载力[21]。对崇明东滩进行互花米草移除后,本地植被的恢复为雁鸭类提供了丰富的植物性食物资源,鸟类物种多样性得到提高[35]。虽然大多数本地鸟类不会取食互花米草种子和根茎,但一些小型鸟类也会在互花米草群落中觅食[15,22],可能是受限于种间竞争压力选择次优食物资源。
2.2 对昆虫群落的影响
昆虫与植物、鸟类关系密切,是生态系统的重要组成部分,大多数鸟类食谱都包含一定数量的昆虫[6]。外来植物入侵后,不同昆虫类群响应机制不同,导致昆虫数量和群落组成发生变化[46]。崇明东滩互花米草入侵后冬季昆虫食物资源与鸟类捕获率之间的关系研究显示,互花米草生境中昆虫密度显著低于芦苇生境,并且鸟类粪便中的食物结构都与芦苇群落中的食物资源结构更为相似[22]。对互花米草与本地植被中昆虫群落的比较研究认为,互花米草分布区昆虫数量、多样性均显著低于芦苇分布区。在三类群落(芦苇、碱蓬、互花米草)中,互花米草群落的昆虫种类和数量最少[47],不同群落中的优势昆虫种类不同,昆虫群落结构发生明显变化[44]。但也有一些研究显示,入侵植物和本地植物组成的混合群落增加了昆虫的物种多样性,为鸟类提供了更丰富的食物资源[48]。目前,还没有研究显示互花米草的入侵可以显著增加昆虫的种类和数量,可能是因为本地昆虫与互花米草缺乏协同进化的过程,需要进一步的深入研究。
2.3 对大型底栖动物的影响
沿海滩涂湿地具有丰富的底栖动物类群,是许多鸟类的重要觅食场所,尤其对迁徙鸟类的中途停歇、能量补充具有重要意义[14,49]。互花米草入侵对大型底栖动物类群的复杂影响,可能通过食物网间接影响着鸟类群落[21]。许多研究揭示了互花米草入侵对大型无脊椎动物类群的影响,但至今还没有统一的结论[45]。不少研究认为互花米草入侵改变了滩涂植被群落、土壤结构等栖息地因子,显著降低了大型底栖动物的密度和丰富度[50- 51]。部分研究认为互花米草入侵后提高了滩涂湿地初级生产力,提高了大型底栖动物的总资源量[52]。还有一些研究认为互花米草入侵对大型底栖动物没有显著影响,种类数量和多样性在不同生境中总体差异不显著[53- 54]。这些不同的研究结果表明,互花米草入侵对底栖动物类群可能存在较为复杂的影响机制。候森林等[55]在江苏射阳口的研究显示,潮间带互花米草的入侵降低了大型底栖动物的物种数、密度、多样性,但在潮沟地带互花米草的入侵却提高了相应指数。还有研究证实,即使互花米草自然演替死亡,但密集的根系改变了栖息地基质的理化性质,导致底栖动物群落在短期内仍无法恢复,间接影响鸟类种群数量的恢复[56- 57]。互花米草的入侵史、入侵程度、研究地特征、底栖动物类群等因素都可能导致不同的研究结果:(1)互花米草入侵时间越长,与本地植被生境底栖动物群落的差异可能越小[45],可能是因为本地动物类群对环境变化的逐步适应;(2)光滩生境更容易受到互花米草入侵的影响,较大的环境改变导致底栖动物类群差异较大[53];(3)不同底栖动物类群对互花米草入侵的响应不同,例如:互花米草入侵后,腹足类、双壳类数量下降显著[53],蟹类动物量却显著增加[58]。
互花米草入侵导致鸟类潜在食物资源的变化,可能逐步改变了鸟类的觅食地选择、觅食行为规律、觅食效率等,最终影响鸟类的身体条件、种群数量、繁殖成功率等。
3 互花米草入侵对鸟类繁殖的影响
鸟类的成功繁殖依赖合适的栖息地[24]。许多研究显示入侵植物给本地鸟类繁殖带来负面影响,降低了繁殖成功率、雏鸟生长速率、巢址忠诚度等[8]。大多数繁殖期鸟类选择本地植被群落筑巢,并避免利用互花米草生境,尤其是互花米草取代本地植被形成的单一密集群落,很难发现本地常见鸟类巢址[34,57]。在我国江苏盐城国家级自然保护区,20%—40%的碱蓬覆盖率是黑嘴鸥(Larussaundersi)、普通燕鸥(Sternahirundo)的理想繁殖生境,互花米草入侵缩减了碱蓬面积,导致该区域鸟类筑巢数量下降[26]。在崇明东滩的研究显示,本地专性鸟类倾向于选择本地芦苇生境筑巢,泛性鸟类则会选择芦苇-互花米草混合生境,没有鸟类对单一互花米草生境表现出正选择[59]。这可能是因为互花米草改变了栖息地结构,造成适宜的巢址微生境多样性下降,过度均质化的生境不能够为鸟类提供合适巢址[60]。
但也有研究显示,入侵植物有时可以为鸟类提供新的繁殖栖息地,没有造成繁殖成功率的下降[48]。在上海崇明东滩,扩散的斑背大尾莺专性在互花米草群落营巢,且繁殖成功率与其在中国北方、日本的原产地几乎没有差异[59]。在美国旧金山海湾,加州长嘴秧鸡、沼泽鹪鹩(Cistothoruspalustris)、阿拉米达歌带鹀(Melospizamelodia)等鸟类也会在互花米草入侵生境中筑巢繁殖[15,42,61]。这些体型较小的鸟类选择在互花米草群落中筑巢,可能是迫于种间竞争压力[59]。因为多数本地鸟类不利用互花米草生境,茂密的植株也为小型种类提供了较好隐蔽。还有研究证实,在一些本地植被退化的生态系统,某些鸟类甚至需要依赖入侵植物完成生活史,优先选择入侵植物作为巢址[62]。例如:美国加州长嘴秧鸡高度依赖互花米草入侵生境筑巢繁殖,对互花米草的清除治理已严重威胁到该物种的正常繁殖[42]。
虽然一些鸟类能够在互花米草生境中繁殖,但可能面临风险。研究显示,阿拉米达歌带鹀在互花米草群落中筑巢的繁殖成功率比在本地植被群落中低30%,可能是因为互花米草大多生长在较低海拔,高潮期潮水会淹没部分鸟巢导致繁殖失败[61]。入侵植物对植被群落结构的影响有时会改变巢址的被捕食率,鸟类在入侵植物中筑巢繁殖较易受到啮齿类的破坏,雏鸟被捕食率显著高于本地植被群落[63]。郝志等[64]在互花米草群落中设置人工巢,结果显示高达67.7%的鸟巢被啮齿类等捕食,可能是因为互花米草群落中啮齿动物天敌的缺乏。互花米草入侵是否导致繁殖鸟类巢捕食率升高,还需要进一步的研究证实。
4 互花米草入侵在鸟类群落层面上的影响
本地植被类型对维持鸟类数量、物种多样性、均匀度具有重要作用。外来植物入侵引起的植被群落结构、栖息地景观格局、可利用食物资源的变化,最终都会影响本地生境中鸟类种群数量和物种多样性[48]。
4.1 对鸟类群落的抑制作用
许多研究表明,互花米草入侵导致本地鸟类种群数量和多样性明显下降,尤其对栖息地专性鸟类和鸻鹬类、雁鸭类、鹤类等中大型水鸟群落抑制作用显著。对崇明东滩春季鸟类群落结构的研究显示,超过90%的鸟类都选择本地植物组成的栖息地,互花米草群落中鸟类的物种数和密度都显著低于其他栖息地类型[34]。震旦鸦雀(Paradoxornisheudei)、东方大苇莺(Acrocephalusorientalis)等专性鸟类避免使用互花米草形成的单一群落,只有一些泛性鸟类选择互花米草栖息地,但数量也显著低于周围芦苇生境[34,65]。在盐城自然保护区,互花米草群落中冬季鸟类多样性指数在各类生境中最低[66]。对崇明东滩繁殖期鸟类群落结构的研究显示,泛性鸟类的物种数和多度在各类生境中差别不大,专性鸟类在芦苇生境和互花米草-芦苇混合生境中差别不大,但在单一互花米草生境中则显著下降[59]。
互花米草入侵对于依赖滩涂湿地觅食的迁徙和越冬期水鸟群落可能更为不利,造成世界范围内许多河口及海岸滨鸟种群数量的下降。相比较互花米草生境,大多数鸻鹬类更倾向于选择光滩或本地植被生境,入侵引起的光滩向草地的转变已严重影响了水鸟的栖息[43,49,57]。在美国威拉帕湾,互花米草面积的逐年增加,造成迁徙过境的水鸟数量逐年下降[14],互花米草的扩张导致1991年至2001年间鸻鹬类的数量减少了67%[31]。在英国南部海岸,大米草的入侵导致滨鸟的死亡率和迁出率上升[11],黑腹滨鹬种群数量的下降甚至在大米草自然演替枯萎后也很难恢复[56]。在我国盐城保护区,丹顶鹤依赖草滩、苇滩等生境觅食,互花米草在生长稳定期盖度可达100%,单一互花米草群落内很难见到丹顶鹤分布[67]。在崇明东滩的互花米草移除示范区内,水鸟的种类与数量均高于对照区,说明互花米草治理后生境能够吸引鸻鹬类、雁鸭类等等水鸟越冬栖息和觅食,鸟类群落结构多样性得到改善[68]。
4.2 对鸟类群落的促进作用
外来入侵植物对本地植被群落结构的改造,对不同的鸟类可能影响不同。一些入侵植物可以提供本地鸟类无法利用的生态位,吸引其他非本地物种栖息利用,在一定程度上促进了本地物种的多样性[69-70]。虽然许多本地鸟类避免使用互花米草生境,但这也为一些鸟类留下了空白生态位,一些小型雀形目鸟类可以很快适应互花米草入侵生境。在崇明东滩的研究显示,一些体型较小的鸟类在种间竞争的压力下会选择镶嵌群落或互花米草群落作为次优栖息地[59]。在某些林地生态系统,入侵植物增加了鸟类栖息地复杂程度,提供了更多的适宜生态位和栖息地类型[8]。在互花米草入侵形成稳定植被群落后,非本地繁殖鸟类斑背大尾莺首次出现在崇明东滩。与本地芦苇生境相比,互花米草群落较少的竞争和较密的茎杆或许能够提供更好的栖息与繁殖生境,并为斑背大尾莺提供了主要食物来源[23]。这项研究表明,互花米草入侵可能提供了本地植被不具备的适宜生境,新的栖息地选择会扩大一些鸟类的分布范围。在美国旧金山海湾的研究也证实,互花米草为体型较小的沼泽鹪鹩提供了合适栖息地,使其扩大了分布范围[15]。
随着外来植物入侵历史的增长,本地生物类群有时可以逐渐适应入侵生境,当移除入侵植物后反而引起物种多样性下降[71]。在较早的研究中认为加州长嘴秧鸡只有在海水高潮期才会利用互花米草生境[72],但随着互花米草入侵历史的增加以及与本地米草的杂交,加州长嘴秧鸡开始适应新的米草群落,并逐渐依赖该生境栖息觅食,对互花米草的快速清除反而造成该种鸟类种群数量的下降[42]。
5 小结与展望
互花米草作为促淤护堤的有效物种,在引种初期起到了良好效果。但随着互花米草的快速扩散增殖,改变了本地植被群落结构,显著降低了栖息地质量,对鸟类等生物类群的负面影响逐渐显现[21]。主要表现在以下方面:(1)植被群落结构不利于鸟类栖息、筑巢、觅食[15,26]。尤其对于鸻鹬类、雁鸭类、鹤类等需要利用开阔栖息地的中大型鸟类,过密且均质化的植被类型可能阻碍了这类水鸟对互花米草生境的利用,在我国盐城保护区丹顶鹤对互花米草沼泽景观具有负向选择[73]。(2)本地鸟类食物资源丰度和多样性下降[22,50]。互花米草入侵引起本地植被、底栖生物、昆虫等鸟类潜在食物资源量下降,即使某些类群数量没有下降,整体食物资源多样性也显著降低,无法满足鸟类的觅食需要。(3)本地鸟类种群数量和物种多样性显著下降[14,59]。互花米草入侵后,许多本地鸟类避免使用互花米草生境,种群数量、物种多样性、繁殖成功率等均受到影响,导致本地鸟类群落结构发生变化。在我国盐城沿海,互花米草入侵已经显著改变了本地植被群落分布,进而影响到鸟类栖息地选择、种群数量、群落结构等,入侵后的植被与鸟类分布格局形成了一定的梯度模式(图1)。
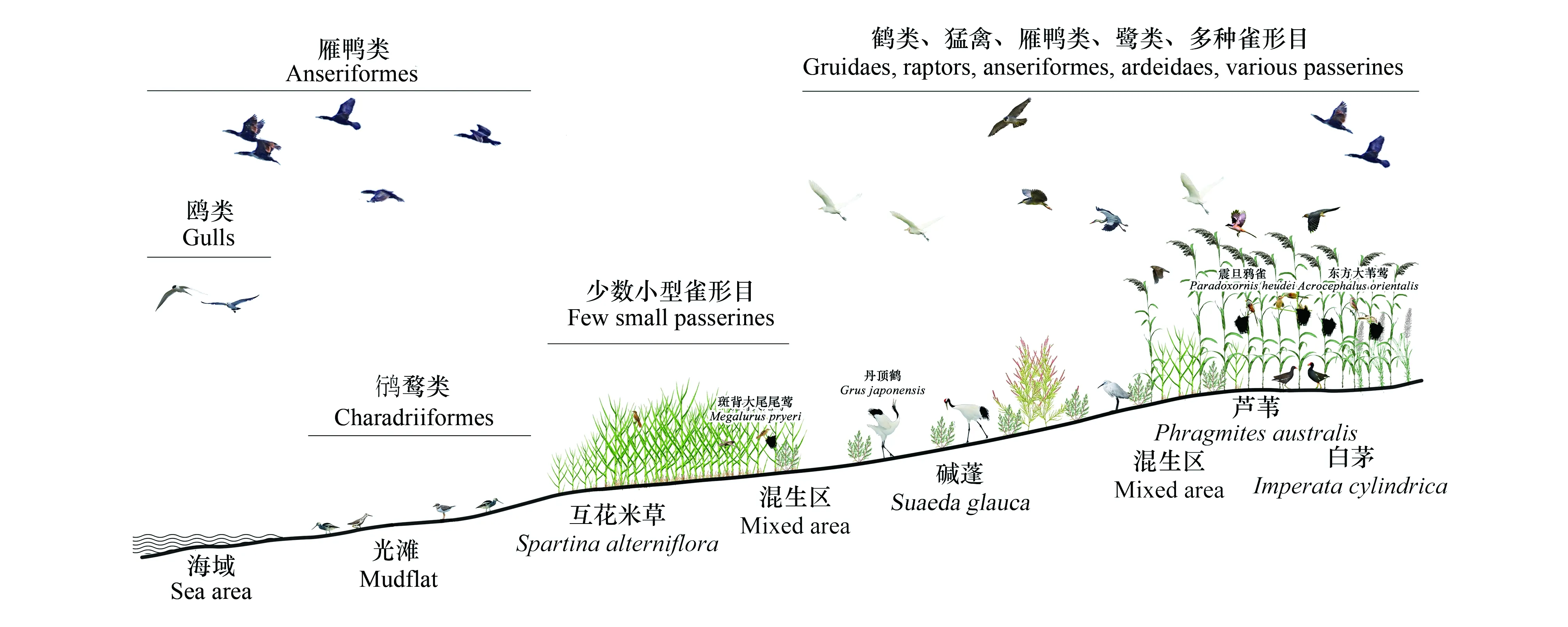
图1 中国盐城沿海互花米草入侵生境植物群落及主要鸟类类群分布模式图Fig.1 Distribution pattern of plant community and main bird groups inSpartina alterniflorainvasive habitat on the Yancheng coast of China
但随着入侵历史的增长,互花米草对本地鸟类群落可能产生不同影响[41]。或许由于长期的协同进化,某些鸟类开始逐步适应互花米草群落,选择入侵生境作为栖息、觅食地,一些种类甚至能够在其中筑巢繁殖[15]。但只限于极少数小型雀形目鸟类,大多数本地鸟类未表现出对互花米草生境的适应,鸟类群落是否随入侵时长发生变化需要进一步的研究。互花米草入侵可能还为非本地鸟类提供了本地鸟类无法利用的空白生态位,增加了一些鸟类的分布范围[23],在一定程度上丰富了本地鸟类物种多样性。因此,互花米草入侵对鸟类群落甚至整个生态系统的影响可能需要更多研究进行综合评价。
未来的研究应关注以下两个方面:(1)开展长时间、大尺度、多因子的监测研究。入侵植物对鸟类的生态影响往往随入侵生境类型、物种特征、斑块成熟程度、入侵历史等因素发生变化,不同地点、不同时期对同一入侵植物的评价可能不同[8]。了解互花米草入侵生境中鸟类的种群、群落乃至生态系统的动态变化, 需要标准化的研究方法进行长期的观察和跟踪。应根据长时间、大尺度、多因子的研究数据,建立互花米草入侵对鸟类群落以及生态系统影响的综合评价模型,从而能够预测互花米草入侵对鸟类等生物类群的潜在影响以及生态系统结构和功能的变化趋势。(2)开展多物种的比较研究,制定合理的互花米草管理对策。目前,对互花米草入侵背景下特定鸟类物种的觅食、繁殖仅有少数研究[23,42,62]。互花米草入侵可能对不同鸟类物种产生不同影响,全面系统的了解互花米草入侵对鸟类群落的影响机制,需要开展多物种的种间比较研究,还需要考虑互花米草入侵对鸟类的食物、天敌等其他生物类群的影响等。在我国崇明东滩实施的互花米草治理项目优化了鸟类栖息地,取得了显著的生态效果[74],互花米草的清除和本地植被的恢复可以显著改善本地鸟类群落状况。但对美国加州互花米草的清除却对已适应并依赖互花米草生境的鸟类造成影响[42],由于动植物间的协同进化关系,对入侵植物的清除有时甚至产生强烈的负面生态效应[75]。在不同地区开展互花米草治理过程中可能需要考虑更多因素,而不是简单的快速清除,应根据入侵生境类型、鸟类物种、植被状况等,制定因地制宜、科学有效的管理对策,逐步恢复本地植被生境。
猜你喜欢
农业灾害研究(2022年7期)2022-09-08
安徽农业科学(2022年14期)2022-08-04
海洋通报(2021年5期)2021-12-21
生态学报(2021年3期)2021-03-31
能源与环境(2020年4期)2020-09-02
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
汉语世界(The World of Chinese)(2018年3期)2018-10-22
厦门航空(2018年4期)2018-04-25
广东农业科学(2017年5期)2017-08-29
