
基于28S rDNA部分序列的金沙江德泽段鲶寄生撒氏虫属(Thaparocleidus)单殖吸虫的系统发育*
2019-07-30 00:40郭苗李寄仟徐伟江范丽仙
云南师范大学学报(自然科学版) 2019年4期
郭苗,李寄仟,徐伟江,2,范丽仙,2
(1.云南师范大学 生命科学学院,云南 昆明 650000;2.生物能源持续开发利用教育部工程研究中心,云南 昆明 650500)
撒氏虫属(ThaparocleidusJain,1952)的模式种是寄生于Wallagoattu鳃上的Thaparocleiduswallagonius[1-2].Gussev分别于1976和1978年将隶属于11个属(指环虫属DactylogyrusDiesing,1850,UrocleidusMueller,1934,锚盘虫属AncylodiscoidesYamaguti,1937,HaplocleidusMueller,1937,撒氏虫属Thaparocleidus,MizelleusJain,1957,NeomurraytremaTripathi,1959,WallagotremaTripathi,1959,伪锚盘虫属PseudoancylodiscoidesYamaguti,1963,ParancylodiscoidesAchmerow,1964,JainiusAchmerow,1964)的48种单殖吸虫归于鲶盘虫属(SilurodiscoidesGussev,1976)[3].依据国际惯例时间优先原则,Lim认为鲶盘虫属为撒氏虫属的次主观异名,将隶属于Silurodiscoides,Parancylodiscoides,Wallagotrema和Urocleidus的70种单殖吸虫重新归属于撒氏虫属,确认了撒氏虫属的有效性[4].在此基础上,指环虫目20个属中保留7个有效属,其他属则被列为同物异名[5],同时律定的92种撒氏虫中有77种为有效种[6-8].我国动物志采用鲶盘虫属阶元进行编目[9].国际上近年对该类群的研究及本研究采用撒氏虫属阶元进行编目和研究.
撒氏虫属单殖吸虫的鉴定通常依据几丁质结构的形态学特征:后吸器(包括锚钩或吸铗、边缘小钩、联结片等)、雄性交接器(包括支持器及交接管)及阴道结构等[10-13].每种单殖吸虫的后吸器及交接器形态各异,具有种和属的鉴定特征[14-15],故可通过几丁质结构,即后吸器和交接器的形态及大小鉴定单殖吸虫[16-19].撒氏虫属后吸器具7对胚钩型边缘小钩,2对中央大钩,其中背中央大钩远大于腹中央大钩,2联结片,其中腹联结片多呈V形等[20].然而,形态学鉴定容易受到客观条件及确立者主观意见的影响,如单殖吸虫个体发育的周期[21-22]、感染方式、汲取营养方式[23]、宿主的差异(体型、地理分布和环境等)[24]及实验操作方法[23,25]等均会导致单殖吸虫形态学鉴定上的问题.故仅以形态学特征作为鉴定依据易产生同物异名以及近似物种难以鉴定等分类学和系统发育研究上的疑难问题.
Cunningham等首次将分子生物学技术应用于单殖吸虫的研究[26-27],其内容主要包括对物种进行鉴定,同时研究遗传演化及种间亲缘关系等[26].为更好地确定撒氏虫属单殖吸虫的分子系统发育关系,学者们进行了广泛的研究.吴相云基于28S rDNA部分序列的研究结果显示寄生于广东番禺苏氏圆腹鱼芒鲶(Pangasiumsutchi)的螺茎撒氏虫(T.campylopterocirrus)与撒氏虫未定种(Thaparocleidussp.BDY)聚为一枝,其余12种撒氏虫聚为一枝,故认为该属为并系群[11];imková等基于28S rDNA部分序列及18S+ITS1+5.8S联合序列证明寄生于鱼芒科(Pangasiidae)鱼类的撒氏虫为单系群[28];而Rajvanshi等对T.devraji进行了重新描述,并基于28S rDNA部分序列证明该属为单系群[29].撒氏虫属是否为单系群有待进一步考证.
本文基于28S rDNA部分序列对9种撒氏虫:牛栏江撒氏虫(T.niulanjiangensissp.nov.)、撒氏虫属未定种 (Thaparocleidussp.isolate G1-6)、撒氏虫属未定种(Thaparocleidussp.isolate I3-1)、凶恶撒氏虫(T.asoti)、奇异撒氏虫(T.varicus)、大茎撒氏虫(T.magnicirrus)、变形撒氏虫(T.obscura)、多形撒氏虫(T.mutabilis)和撒氏虫属未定种(Thaparocleidussp.isolate MG-2018)进行分子系统发育研究,以期探讨撒氏虫属单殖吸虫的分类、同物异名及系统发育关系等问题.
1 材料与方法
1.1 实验材料
鲶鱼宿主分别于2017年6月、11~12月、2018年12月采自云南省金沙江水系牛栏江支流德泽段(N25°57′5″,E103°36′9″).将鳃丝置于生理盐水中,在光学显微镜Olympus CX-41下检获虫体,制作水封片于显微镜下观察其形态结构,并利用DigiRetina 16显微成像系统和TCapture Imaging Compose显微数码采集分析系统拍照并测量.随后,将虫体小心移至加入适量无水乙醇的1.5 mL离心管中,置于-4 ℃保存,用于分子实验研究.
1.2 分子鉴定
保存于无水乙醇的样品按照美国OMEGA公司D3390-01石蜡DNA试剂盒提取其DNA,并对28S rRNA基因部分序列进行扩增.PCR反应的上游引物为C1[30]:5′-ACCCGCTGAATTTAAGCAT-3′;下游引物为D2[31]:5′-TGGTCCGTGTTTCAAGAC-3′.反应条件:95 ℃预变性3 min,42 ℃退火2 min,72 ℃延伸90 s;95 ℃变性45 s,50 ℃退火45 s,72 ℃延伸90 s,4个循环;95 ℃变性20 s,52 ℃退火20 s,72 ℃延伸90 s,25个循环;最后72 ℃延伸5 min.取5 μL产物进行1%琼脂糖凝胶电泳检测,测序由昆明硕擎生物科技有限公司完成.
运用DNAMAN7.0[32]对所测序列采用统一标准进行拼接,随后通过Bankit软件编辑,提交NCBI数据库并获得GenBank登录号.基于BLAST(Basic Local Alignment Search Tool)选择GenBank中同属物种进行序列分析,用MEGA7.0[33-34]计算序列间转换/颠换值,并基于双参数模计算遗传距离,以伪锚盘虫属(Pseudancylodiscoides)单殖吸虫作为外群,通过最大似然法(maximum likelihood,ML)、邻接法(neighbor-joining,NJ)和最大简约法(maximum parsimony,MP)构建50%合一树.
2 结 果
2.1 基于28S rDNA部分序列撒氏虫属单殖吸虫的分类鉴定
目标序列的宿主鱼、采集地、GenBank登录号及长度等信息见表1.基于双参数模的遗传距离见表2.
经测序获得目标序列873 bp共计保守位点449个,变异位点414个,简约信息位点297个,自裔位点117个,平均转换/颠换值R为1.2.A、T、G和C碱基的平均含量分别为23.3%、27.9%、28.1%和20.6%.
牛栏江撒氏虫与GenBank中21种撒氏虫属单殖吸虫的同源性序列平均相似度在83.48%~92.48%之间,其中与大茎撒氏虫(GenBank登录号:MH213065)的相似度最高为92.48%,基于双参数模遗传距离的范围为0.080~0.341,该种吸虫与大茎撒氏虫(GenBank登录号:MH213065)、撒氏虫属未定种(Thaparocleidussp.isolate MG-2018)和撒氏虫属未定种(Thaparocleidussp.NY1)的遗传距离最近,为0.080(见表2),三者的56个碱基存在差异,差异值为6.4%(56/873);撒氏虫属未定种(Thaparocleidussp.isolate G1-6)与上述21种撒氏虫属单殖吸虫的同源性序列平均相似度在83.64%~99.74%之间,其中与奇异撒氏虫(GenBank登录号:DQ157668)的相似度最高为99.74%,基于双参数模遗传距离的范围为0.005~0.285,该种吸虫与奇异撒氏虫的遗传距离最近,为0.005(见表2),二者的68个碱基存在差异,差异值为8.0%(68/857);撒氏虫属未定种(Thaparocleidussp.isolate I3-1)与上述21种撒氏虫属单殖吸虫的同源性序列平均相似度在85.51%~99.14%之间,其中与奇异撒氏虫(GenBank登录号:MH094192)的相似度最高为99.14%,基于双参数模遗传距离的范围为0.005~0.285,该种吸虫与奇异撒氏虫(GenBank登录号:MH094192)的遗传距离最近,为0.004(见表2),二者的61个碱基存在差异,差异值为7.2%(61/847).
牛栏江撒氏虫与大茎撒氏虫、弓阴撒氏虫相似,三者支持器均为槽状,但弓阴撒氏虫末端中空膨大呈Ω状,大茎撒氏虫与牛栏江撒氏虫末端弯曲处均具一突起和一环形小孔,但前者末端有多个三角突起,而后者于突起对立面伸出一柄状结构.系统进化树显示该种吸虫单独成支(图3 Clade 6),且支持率分别为86%、78%和54%,综合形态学及分子系统发育研究,认为该种为科学上1新种;撒氏虫属未定种(Thaparocleidussp.isolate G1-6)与王建强[19]描述的撒氏虫属未定种(Thaparocleidussp.1)相似,但二者支持器结构明显不同,本种支持器简单,一字形,末端分化为两三角形突起,交接管于中央穿过,而后者末端具一小环,交接管从此穿过.进化树显示该种吸虫与奇异撒氏虫(GenBank登录号:DQ157668)聚为一支(图1 Clade Ⅰ),但支持率较低,分别为89%、90%和68%,综合形态学及分子系统发育研究,认为该种为科学上1未记录种;撒氏虫属未定种(Thaparocleidussp.isolate I3-1)与奇异撒氏虫相似,二者支持器末端均有一不规则长舌状物,但本种支持器距末端1/3处弯曲,长舌状物与弯曲凸面同向,而后者支持器末端具多个突起.除此之外,本种阴道结构呈胃肠状,由一段短的狭管和一相互折叠的宽管组成,后者呈腊肠状.进化树显示该种吸虫与本研究奇异撒氏虫(GenBank登录号:MH094192)聚为一支(图3Clade 2),但支持率低,分别为58%、51%和61%,结合形态学及分子系统发育研究,认为该种为科学上1未记录种.
2.2 基于28S rDNA部分序列撒氏虫属单殖吸虫的分子系统发育研究
基于28S rDNA部分序列,利用MEGA7.0构建的以NJ树为基础的NJ/ML/MP 50%合一树显示(图1),撒氏虫属为非单系群.本研究22种撒氏虫聚为Clade Ⅰ和Clade Ⅱ两个大的进化支.Clade Ⅰ包括本研究撒氏虫属未定种(Thaparocleidussp.isolate G1-6)和来自重庆的2种撒氏虫:奇异撒氏虫(T.varicus)(GenBank登录号:DQ157668)、筒鞘撒氏虫(T.infundibulovagina).Clade Ⅱ由Clade 1、Clade 2、Clade 3、Clade 4、Clade 5及Clade 6共六个进化支构成,其中,由Clade 1、Clade 2和Clade 3组成的进化支中8种撒氏虫包括本研究4种撒氏虫:变形撒氏虫、奇异撒氏虫(GenBank登录号:MH094192)、撒氏虫属未定种(Thaparocleidussp.isolate I3-1)和多形撒氏虫 (GenBank登录号:MK353164),来自广东的4种撒氏虫:多形撒氏虫(GenBank登录号:EF100550)、弓阴撒氏虫(T.omegavagina)、撒氏虫属未定种(Thaparocleidussp.2 XW-2007)、螺鞘撒氏虫(T.cochleavagina);Clade 4包括本研究凶恶撒氏虫(GenBank登录号:MG601545)与来自重庆的2种撒氏虫:凶恶撒氏虫(GenBank登录号:DQ157669)、撒氏虫属未定种(Thaparocleidussp.NY2);Clade 5包括来自捷克的2种撒氏虫:T.siluri与T.vistulensis;Clade 6包括本研究3种撒氏虫:牛栏江撒氏虫、大茎撒氏虫(GenBank登录号:MH213065)、撒氏虫属未定种(Thaparocleidussp.isolate MG-2018)和来自重庆的2种撒氏虫:撒氏虫属未定种(Thaparocleidussp.1 XW-2007)、撒氏虫属未定种(Thaparocleidussp.NY1),以及来自广东的大茎撒氏虫(GenBank登录号:EF100549).
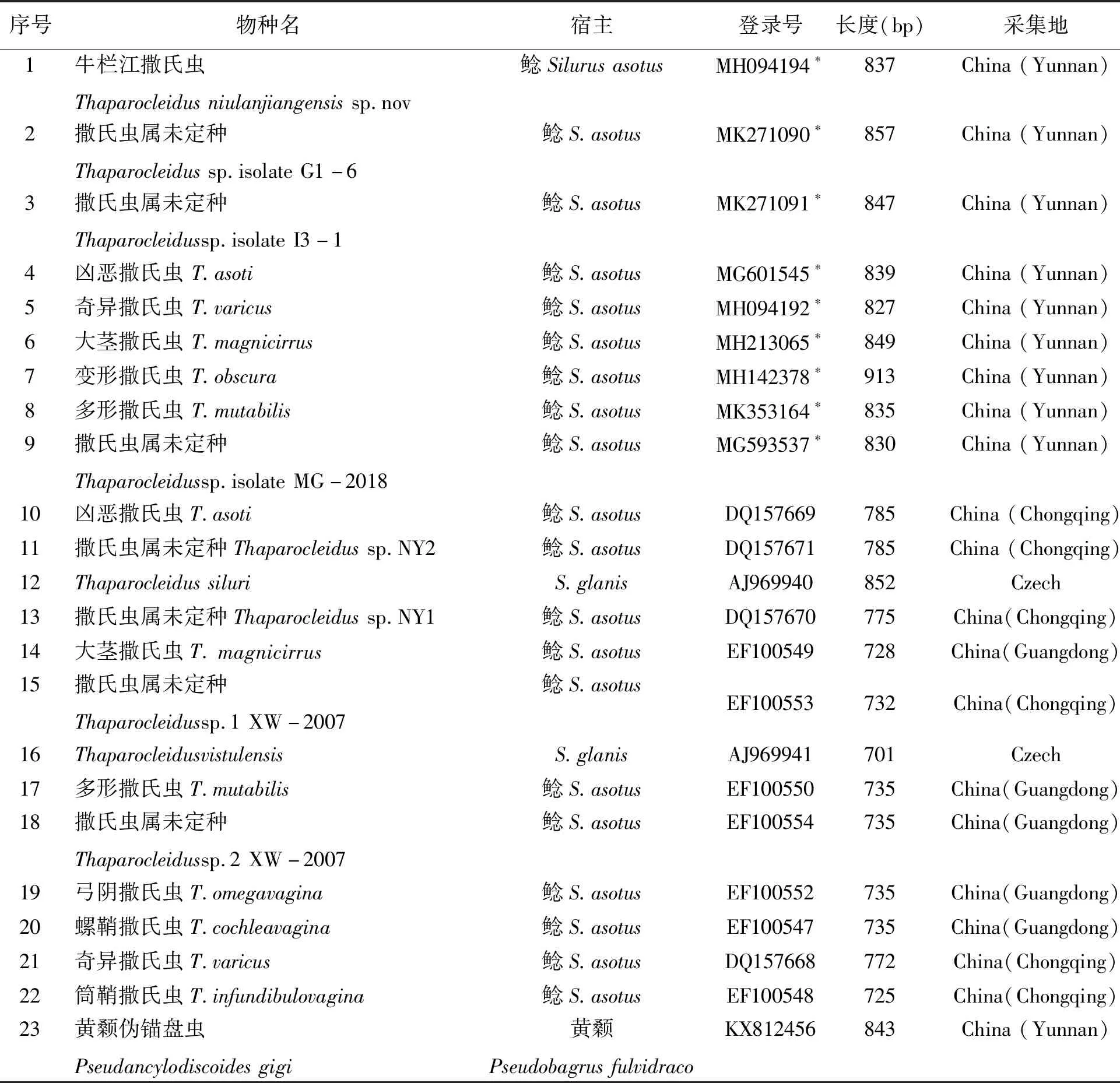
表1 撒氏虫属单殖吸虫宿主、GenBank登录号、长度及采集地Table 1 Information of Thaparocleidus species,including host,accession number of GenBank,localities and length
*表示本研究所获得的基因登录号
表2撒氏虫属单殖吸虫28S rDNA部分序列基于双参数模的遗传距离
Table 2 The genetic distance based on Kimura-two-parameter ofThaparocleidusmonogenean species based on 28S rDNA partial sequence

123456789101112131415161718192021222310.0180.0170.0130.0170.0110.0160.0170.0110.0130.0130.0130.0110.0110.0120.0140.0160.0160.0160.0170.0180.0180.02720.1820.0150.0130.0150.0130.0160.0160.0130.0130.0130.0140.0130.0130.0140.0140.0160.0150.0150.0160.0000.0030.02430.1680.1390.0130.0020.0120.0050.0080.0120.0130.0130.0120.0120.0120.0120.0110.0040.0030.0040.0080.0150.0160.02440.1220.1080.1080.0130.0090.0130.0130.0090.0000.0000.0090.0090.0090.0100.0090.0120.0130.0130.0130.0130.0140.02350.1700.1350.0040.1030.0120.0050.0080.0120.0130.0130.0120.0120.0120.0120.0120.0040.0020.0040.0080.0150.0160.02360.0800.1060.0960.0550.0940.0120.0130.0000.0090.0090.0080.0000.0010.0050.0080.0120.0120.0120.0130.0130.0140.02370.1700.1440.0180.1060.0170.0960.0080.0120.0130.0130.0120.0120.0120.0120.0120.0030.0050.0040.0090.0160.0160.02480.1740.1500.0410.1150.0400.1030.0450.0130.0130.0130.0130.0130.0130.0120.0130.0080.0070.0080.0050.0160.0160.02490.0800.1060.0960.0550.0940.0000.0960.1030.0090.0090.0080.0000.0010.0050.0080.0120.0120.0120.0130.0130.0140.023100.1220.1080.1080.0000.1030.0550.1060.1150.0550.0000.0090.0090.0090.0100.0090.0120.0130.0130.0130.0130.0140.023110.1220.1080.1080.0000.1030.0550.1060.1150.0550.0000.0090.0090.0090.0100.0090.0120.0130.0130.0130.0130.0140.023120.1160.1130.0920.0580.0910.0420.0920.1030.0420.0580.0580.0080.0080.0090.0040.0110.0120.0120.0130.0140.0140.022130.0800.1060.0960.0550.0940.0000.0960.1030.0000.0550.0550.0420.0010.0050.0080.0120.0120.0120.0130.0130.0140.023140.0820.1040.0940.0530.0920.0010.0940.1010.0010.0530.0530.0430.0010.0050.0080.0120.0110.0120.0130.0130.0130.023150.0970.1100.0920.0590.0890.0200.0960.1010.0200.0590.0590.0500.0200.0210.0090.0120.0110.0120.0120.0140.0140.022160.1210.1140.0870.0620.0860.0460.0940.1010.0460.0620.0620.0140.0460.0480.0530.0120.0120.0120.0130.0140.0140.023170.1670.1390.0140.1010.0120.0910.0040.0400.0910.1010.1010.0870.0910.0890.0910.0890.0040.0030.0080.0160.0160.024180.1680.1370.0040.1050.0030.0910.0170.0400.0910.1050.1050.0910.0910.0890.0870.0860.0120.0040.0080.0150.0160.024190.1680.1390.0110.1060.0090.0920.0120.0400.0920.1060.1060.0890.0920.0910.0920.0890.0080.0090.0080.0150.0160.024200.1700.1440.0430.1120.0380.1030.0460.0150.1030.1120.1120.1030.1030.1010.0990.0990.0420.0420.0420.0160.0160.024210.1820.0050.1390.1080.1350.1060.1440.1500.1060.1080.1080.1130.1060.1040.1100.1140.1390.1370.1390.1440.0030.024220.1820.0040.1410.1120.1370.1060.1460.1520.1060.1120.1120.1140.1060.1040.1120.1140.1410.1390.1410.1460.0040.024230.3410.2850.2890.2750.2820.2610.2870.2940.2610.2750.2750.2630.2610.2610.2520.2680.2870.2820.2890.2850.2850.2850.027
对角线以下为遗传距离,对角线以上为标准差,物种顺序如表1
3 讨论与小结
基于28S rDNA部分序列构建的系统发育树显示,撒氏虫属为非单系群.撒氏虫属未定种(Thaparocleidussp.isolate G1-6)与筒鞘撒氏虫和奇异撒氏虫(GenBank 登录号:DQ157668)聚为Clade Ⅰ,其余虫种(除T.siluri、T.vistulensis寄生于S.glanis外均寄生于鲶鱼S.asotus)均聚为Clade Ⅱ;其中奇异撒氏虫(GenBank 登录号:DQ157668)与撒氏虫属未定种(Thaparocleidussp.isolate G1-6)组成的进化支位于系统树的基部,推测在所有采用的22种撒氏虫属单殖吸虫中,奇异撒氏虫进化地位较原始.
吴相云等[11,20]依据雄性交接器的形态学特征,将14种撒氏虫分为4个类群,其中前三个类群分别对应于本研究的Clade Ⅰ、由Clade 1、Clade 2和Clade 3组成的进化支以及由Clade 4、Clade 5和Clade 6组成的进化支.其中,由Clade 1、Clade 2和Clade 3组成的进化支中8种撒氏虫的主要特征为支持器大型,末端具复杂几丁质结构,交接管基部呈瓶状.由Clade 4、Clade 5和Clade 6组成的进化支中11种撒氏虫的交接器具相似的特征:支持器槽状,末端弯曲且具一环形小孔,交接管由此穿过.与吴相云等的研究结果出现分歧的是筒鞘撒氏虫和奇异撒氏虫的分类地位,在其研究中,因交接器明显小于其他撒氏虫、支持器末端具一突起、交接管极细等特征,筒鞘撒氏虫单独占一个Clade,而奇异撒氏虫则聚于本研究由Clade 1、Clade 2和Clade 3组成的进化支中,但在本研究中两者聚为一支(Clade Ⅰ).本研究与Verma等[2,33]的结果一致,推测出现该现象的原因可能是两种撒氏虫的28S rDNA分子序列碱基相似性较高,两者的遗传距离仅为0.004(表2),验证了imková等[28]的结论:形态学差异较大的撒氏虫却显示较低的分子差异.且二者分布地均在重庆市荣昌区,可能符合单殖吸虫物种形成中的同域物种形成.
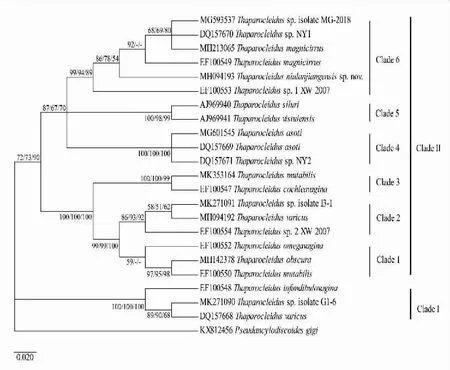
-表示置信度<50%图1 基于28S rDNA部分序列,撒氏虫属单殖吸虫以NJ树为基础的NJ/ML/MP 50%合一树Fig.1 The consensus tree of NJ/ML/MP analysis based on NJ tree topology constructed from 28S rDNA partial sequence of Thaparocleidus monogenean species
目前我国有关撒氏虫属的研究还较少,该属内单殖吸虫的同物异名现象及系统发育关系,如是否为单系群、与其他属中相似种的亲缘关系等均有待进一步的深入研究.随着更多的物种和分类单元被发现,种间变异和种内变异的形态学特征被不断揭示,形态分类学方法存在一定的局限性和不足.分子生物学技术被广泛应用于种间鉴定和种间亲缘关系的研究中,DNA序列分析能在小范围内有效解决由于形态特征缺失或用形态分类学方法无法客观辨别的某些种类的分类地位问题,从基因型的角度补充或修正了一些形态分类学的不足,纠正以往分类上的偏差.但目前数据库中鱼类单殖吸虫的分子系统研究数据非常有限,因此,后续开展单殖吸虫的分子标记和分子系统发育研究时,选择适合各类群的靶基因或开展线粒体全基因组的研究,将极大程度上完善单殖吸虫分子鉴定及系统发育关系研究.
致谢:衷心感谢中国科学院昆明动物研究所杨君兴研究员、潘晓赋副研究员和云南水投—滇池补水工程公司德泽枢纽管理部鱼类增殖流放站杨世论老师在宿主样品采集和鉴定方面给予的帮助.
猜你喜欢
区域治理(2022年40期)2022-11-27
种子(2021年2期)2021-03-31
天津农学院学报(2020年2期)2020-07-13
动漫界·幼教365(小班)(2019年10期)2019-10-28
动漫界·幼教365(大班)(2019年10期)2019-10-28
动漫界·幼教365(中班)(2019年10期)2019-10-28
浙江化工(2019年5期)2019-06-04
办公室业务(2016年19期)2016-11-25
中国继续医学教育(2015年2期)2016-01-06
振动、测试与诊断(2014年6期)2014-03-01
