
温度对椰枣种子发芽和生理特征的影响
2019-09-10 21:32李东霞黄丽云徐中亮符海泉张宁
南方农业学报 2019年8期
李东霞 黄丽云 徐中亮 符海泉 张宁





摘要:【目的】研究不同溫度及变温处理对椰枣种子发芽和相关生理指标的影响,为椰枣种子发芽和种苗繁育提供理论依据。【方法】以酋长椰枣(Fard)为试验材料,以高效双向温度梯度系统为培养环境(培养温度20~50 ℃,温差2~30 ℃),分析不同温度梯度和温差组合下椰枣种子的发芽情况及胚乳和胚轴中的可溶性蛋白浓度、超氧化物歧化酶(SOD)活力、过氧化物酶(POD)活力、谷胱甘肽还原酶(GR)活力和过氧化氢(H2O2)含量差异。【结果】适合椰枣种子发芽的光培养温度范围为22~36 ℃,暗培养温度范围22~38 ℃。播种后第21 d,从椰枣种子未发芽胚乳到正常发芽胚乳其可溶性蛋白浓度呈逐渐下降趋势,其中,未发芽胚乳的可溶性蛋白浓度显著高于其他4种胚乳或胚轴(P<0.05,下同);白色胚轴中的总SOD活力和POD活力显著高于黄色胚轴和胚乳。【结论】温度是椰枣种子发芽的重要因素,在适宜的培养温度范围内,椰枣种子发芽快,发芽率高,胚轴长势好。椰枣种子发芽后胚乳中的可溶性蛋白浓度显著低于发芽前。
关键词: 椰枣;种子;温度;发芽;生理特征
中图分类号: S667.904.3 文献标志码: A 文章编号:2095-1191(2019)08-1764-07
Effects of different temperatures on germination and physiological characteristics of date palm seed
LI Dong-xia, HUANG Li-yun, XU Zhong-liang, FU Hai-quan, ZHANG Ning
(Coconut Research Institute, Chinese Academy of Tropical Agricultural Sciences, Wenchang, Hainan 571339, China)
Abstract:【Objective】In order to provide theoretical basis for date palm seed germination and seedling propagation, this experiment was conducted to study the difference on seed germination and physiological characteristics under different temperature treatments and poikilothermic treatments. 【Method】Seeds of cultivar Fard were used as materials, highly efficient bi-directional temperature gradient system was used as cultivating device. The seed germination, soluble protein concentration of endosperm and hypocotyl, superoxide dismutase(SOD) activity, peroxidase(POD) activity, glutathione reductase(GR) activity, hydrogen peroxide(H2O2) content were detected under 20-50 ℃ cultivating temperature and 2-30 ℃ difference in temperature. 【Result】It was suitable for date palm seed germination when light culture was conducted under 22-36 ℃ and dark culture was conducted under 22-38 ℃. The soluble protein concentration of ungerminated date palm seed endosperm to the normal germina-ted endosperm decreased gradually on the 21st day after sowing. The soluble protein concentration of ungerminated endosperm was significantly higher than that of other four ungerminated endosperms and hypocotyl,the soluble protein concentration of ungerminated endosperm was significantly higher than that of other four kinds of endosperms or hypocotyl(P<0.05,the same was below). The SOD activity and POD activity of white hypocotyls were higher than that of yellow hypocotyls and endosperm. 【Conclusion】Temperature is an important factor affecting seed germination in date palm. Under the suitable culture temperature range, the date palm seeds germinate quickly, the germination rate is high, and hypocotyls grow well. The soluble protein concentration in endosperm after germination is significantly lower than before germination.
Key words: date palm; seed; temperature; germination; physiological characteristics
0 引言
【研究意义】椰枣(Phoenix dactylifera L.)为雌雄异株,多年生单子叶植物,是中东和非洲撒哈拉地区的一种重要果树,“上干下湿”是其最理想的生活环境,树龄可长达百年。椰枣果实的形状有圆形和椭圆形,含糖量高,营养价值极高,因此被誉为阿拉伯民族之树,因其能很好地适应干旱条件,又有“沙漠面包”之称(Parisa Eshraghi和田郎,2011;辛成齐,2015;李东霞等,2017)。通过种子进行有性繁殖是植物繁衍的重要手段之一,而种子发芽会潜在影响植物种群的动态和组成,任何不利于种子发芽的因子均有可能最终影响种群的稳定性(Jusaitis et al.,2004)。椰枣能通过种子进行有性繁殖,而温度是影响椰枣种子发芽的主要环境因子之一,种子发芽周围的环境温度太高或太低均会影响椰枣种子活力,造成发芽和出苗不良。因此,研究不同温度和变温处理对椰枣种子发芽和生理特性的影响,对指导椰枣生产实践具有重要意义。【前人研究进展】Asomaning等(2011)采用双向温度系统培养榄仁树,发现榄仁树在恒温35/35 ℃的发芽率为88%,在变温35/40 ℃的发芽率为100%,40/25 ℃的发芽率为95%,20/40 ℃和40/30 ℃的发芽率为92.5%,说明变温环境更适宜榄仁树发芽。鲁小名等(2012)、汪建军等(2016)研究发现种子能通过感应周围环境的变温、低温或高温而判断四季的变化及周围环境的差异,以选择恰当时机开始其生活史。尚瑞广等(2014)通过人工气候箱对玛咖种子萌发进行研究,发现15~30 ℃是玛咖适宜的萌发温度,而不同时长光照和黑暗处理下玛咖种子均能萌发,因此玛咖种子萌发需光特性不明显。钱长江等(2017)研究发现百脉根种子发芽势和发芽指数均呈先升高后降低的变化趋势,在20 ℃时的发芽势和发芽指数最高。王鸿彬等(2018)研究发现温度变化对江南油杉种子萌发影响显著,其中江南油杉种子在25 ℃下的萌发率最高,当温度升高到30和35 ℃时江南油杉种子萌发相关指标明显降低。另外,龙汉利等(2013)研究发现桢楠种子萌发期间,超氧化物歧化酶(SOD)和过氧化物酶(POD)活性从桢楠种子发芽试验之日至萌发高峰期随时间推移二者的活力逐渐递增,而从萌发高峰期至萌发停止其酶活力又呈逐渐递减趋势,同时可溶性糖和可溶性蛋白含量呈先升高再降低趋势,说明在桢楠种子发芽的不同阶段,SOD和POD酶活力、可溶性糖和可溶性蛋白含量呈现一个不断变化的状态。【本研究切入点】目前,关于温度对椰枣种子发芽和生理特征影响的研究鲜见报道。【拟解决的关键问题】采用双向温度梯度系统设置不同的温度梯度和温差组合,探究椰枣种子对温度和温差的耐受性及在不同温度环境下椰枣种子可溶性蛋白浓度及SOD、POD、谷胱甘肽还原酶(GR)活性和过氧化氢(H2O2)含量差异,为指导椰枣种苗繁育提供理论依据。
1 材料与方法
1. 1 试验材料
供试椰枣为迪拜生产的酋长椰枣(Fard)。
1. 2 试验方法
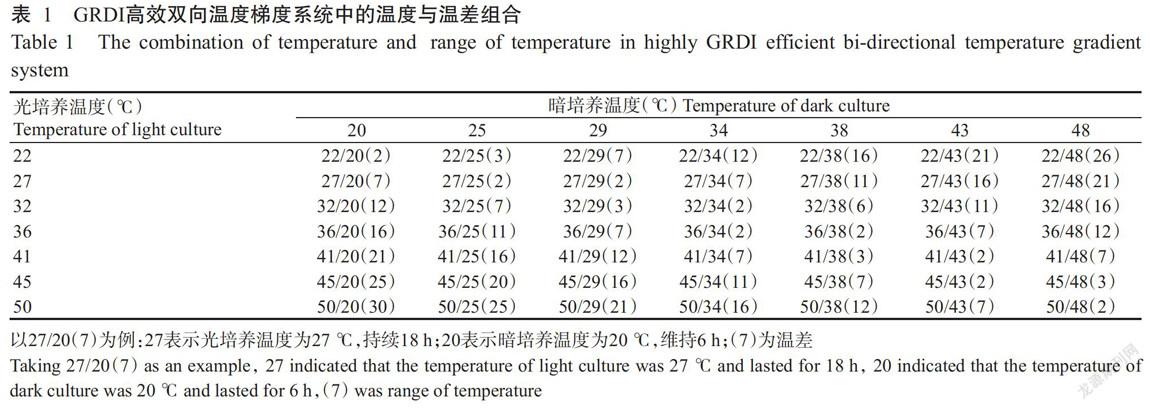
1. 2. 1 试验设计 使用英国Grant的GRD1高效双向温度梯度系统对椰枣种子进行孵育,将仪器最高温度设为50 ℃,最低温度设为20 ℃,选取仪器导出数据中的49个不同光培养和暗培养的温度组合进行椰枣种子发芽试验(表1),其中,光培养18 h的温度范围为22~50 ℃,暗培养6 h的温度范围为20~48 ℃,以最高温度减去最低温度计算温差,计算获得的温差范围为2~30 ℃。
取带有果肉的椰枣果,将果肉剥除,先用洗洁精将粘在种子上的果肉清洗干净,再用自来水将泡沫清洗干净;用2.5%次氯酸钠溶液对种子灭菌5 min,再用灭菌纯水将种子清洗3遍,置于GRD1双向温度梯度系统中进行发芽试验,仪器温度设为20~50 ℃,每天早上用喷壶喷纯水,保持种子湿润。
1. 2. 2 测定项目及方法 分别在播种后7、9、14、18、21和27 d统计不同培养环境下的发芽率;并对播种后21 d未发芽的距离胚0.5 cm的胚乳(以下简称未发芽胚乳)、长有黄色胚轴距离胚0.5 cm的胚乳(黄胚轴胚乳)、白色胚轴距离胚0.5 cm的胚乳(白胚轴胚乳)、黄色胚轴和白色胚轴的可溶性蛋白浓度、总SOD活力、POD活力、GR活力和H2O2含量进行测定。可溶性蛋白浓度采用考马斯亮蓝法测定,SOD活力采用氮蓝四唑(NBT)光还原法测定,POD活力采用愈创木酚法测定,GR活力采用紫外分光光度法测定,H2O2含量采用钼酸显色法测定。以上测定项目均使用南京建成生物工程研究所南京建成科技有限公司生产的相关试剂盒进行测定。发芽率GR(%)=n/N×100(n为播种后某一天正常发芽种子数,N为供试种子数)。
1. 3 统计分析
试验数据使用Excel 2007进行整理和绘图,并以SAS 9.1进行单因素方差分析(One-way ANOVA)和Duncan’s多重比较。
2 结果与分析
2. 1 椰枣种子在双向温度梯度系统中的发芽情况
椰枣种子在适合的温度环境下吸水膨胀,其内部养分转化,从胚乳中分解、运输到种胚,种胚膨胀后从胚孔中顶出,露白,之后逐渐生长为胚轴、胚根和胚芽。在本研究中,种子漏白则记为发芽。由图1可知,播种后第21 d,在不同的双向温度组合下椰枣种子的发芽情况各不相同,其中,27/20 ℃温度组合下,椰枣种子发芽且已长出较长的胚轴;27/43 ℃温度组合下,部分椰枣种子露白;50/20 ℃温度组合下,椰棗种子表皮颜色比27/20 ℃和27/43 ℃温度组合下的种子表皮颜色深,且所有种子均未发芽。
由表2可知,在不同的双向温度组合下,椰枣种子在播种后不同时间的发芽率存在明显差异。其中,播种后第9 d发芽的组合有22/20 ℃、27/20 ℃、32/20 ℃、22/25 ℃、32/25 ℃、36/25 ℃、22/29 ℃、27/29 ℃、32/29 ℃、36/29 ℃、22/34 ℃、27/34 ℃、32/34 ℃、36/34 ℃、22/38 ℃、27/38 ℃和32/38 ℃,其溫差范围为2~16 ℃,维持18 h的光培养温度范围为22~36 ℃,维持6 h的暗培养温度范围为20~38 ℃;播种后第21 d仍未发芽的温度温差组合有41/20 ℃、45/20 ℃、50/20 ℃、41/25 ℃、45/25 ℃、50/25 ℃、45/29 ℃、50/29 ℃、45/34 ℃、50/34 ℃、41/38 ℃、45/38 ℃、50/38 ℃、36/43 ℃、41/43 ℃、45/43 ℃、50/43 ℃、36/48 ℃、41/48 ℃、45/48 ℃和50/48 ℃,其温差范围为2~30 ℃,维持18 h的光培养温度范围为36~50 ℃,维持6 h的暗培养温度范围为20~48 ℃;其余双向温度组合在播种后第14或18 d才开始发芽,其所处坏境的温差范围为2~26 ℃,维持18 h的光培养温度范围为22~41 ℃,维持6 h的暗培养温度范围为20~48 ℃。
由表1和图2可知,播种后第27 d,可将椰枣种子在双向温度梯度系统中的发芽情况分为3种,即不发芽、发芽率分别为20%~60%和61%~100%。不发芽温度:光培养温度为45和50 ℃、培养温度为20~48 ℃;光培养温度为41 ℃,暗培养温度分别为25、38、43和48 ℃;光照培养温度为36 ℃,暗培养温度分别为43和48 ℃。椰枣种子发芽率在60%以上:光培养温度为22、27、32 ℃,暗培养温度为20~34 ℃;光培养温度为36 ℃,暗培养温度为25和29 ℃。
2. 2 不同发芽情况下椰枣种子中可溶性蛋白浓度的差异性分析结果
对双向温度梯度系统中3种不同发芽情况下的椰枣种子胚乳和胚轴中的可溶性蛋白浓度进行测定,结果(图3)发现,从未发芽胚乳到黄胚轴胚乳再到白胚轴胚乳过程中可溶性蛋白浓度呈下降趋势,其中,未发芽胚乳的可溶性蛋白浓度最高,显著高于其他4种胚乳或胚轴(P<0.05,下同);黄胚轴胚乳及黄色胚轴中的可溶性蛋白浓度均显著低于未发芽胚乳,但显著高于正常生长胚轴的胚乳和胚轴,即白胚轴和白胚轴胚乳。
2. 3 不同发芽情况下椰枣种子抗氧化物酶活性的变化
由图4可知,双向温度梯度系统中发芽情况不同的胚乳和胚轴中总SOD活力、POD活力、GR活力和H2O2含量也不同。其中,白色胚轴总SOD活力最高,未发芽胚乳→黄胚轴胚乳→白胚轴胚乳的总SOD活力呈逐渐增加趋势。白色胚轴的POD活力最高,未发芽胚乳的POD活力最低;黄胚轴胚乳、白胚轴胚乳和黄色胚轴中的POD活力无显著差异(P>0.05,下同)。胚乳和胚轴的GR活力无显著差异。胚轴的H2O2含量显著高于胚乳的H2O2含量,但胚乳间的H2O2含量和胚轴间的H2O2含量均无显著差异。
3 讨论
3. 1 适宜的温度对椰枣种子发芽至关重要
影响植物种子发芽率和出苗率的主要因子之一是温度,在种子萌发时其内部会进行非常活跃的代谢反应。每一种植物的种子都有其适宜发芽的温度范围,温度太高、太低或变温幅度太大均会影响种子的活力,造成发芽和出苗不良(刘小金等,2014);植物生长非常关键的两个时期包括种子发芽和成苗早期,而此时对外界环境因子非常敏感。适宜温度是种子发芽所需的必要条件之一,掌握适宜种子发芽的温度范围在指导植物苗木生产实践中具有重要意义(Orozco-Almanza et al.,2003;王鸿彬等,2018)。种子在适宜的温度范围内发芽时,其首先吸水、促进种皮软化,加快一些相关酶促反应的速度,加强呼吸作用,为种子发芽和胚的生长提供足够的能量和物质基础(李雄等,2014;徐恒恒等,2014;董小云等,2018)。当种子所处的温度太高时,种子发芽的相关指标明显降低,究其原因可能是由于温度太高影响了种子内部酶变性、膜的透性和膜结合的活性(宋兆伟等,2009)。有研究认为,恒温比变温更适合江南油杉种子萌发,变温的萌发效果不如恒温的好(王鸿彬等,2018);但也有研究表明,一些植物种子在变温环境下比在恒温环境的发芽率高,导致此结果的原因可能是高低温互变能促进种子内部各类酶活性的生理活动,使得种皮收缩,从而促进了种子与外界坏境的气体交换(Meulebrouck et al.,2008;Qu et al.,2008)。在本研究中,播种后第27 d,椰枣发芽率在60%以上其培养环境的光培养温度范围为22~36 ℃,暗培养温度为20~38 ℃;当光培养温度高于41 ℃、暗培养温度高于36 ℃时,椰枣种子不能正常发芽。由此可见,椰枣发芽的临界温度范围可能在36~41 ℃,培养温度高于此温度范围时,可能因影响种子对水分的吸收而影响发芽,也可能由于高温使得内部酶变性而影响发芽。本研究中,椰枣发芽率高、发芽快的温差范围为2~16 ℃、温度范围为22~38 ℃。唐安军等(2005)研究发现,相对于恒温培养环境,在20/30 ℃变温环境下更有利于棕榈种子的萌发,且萌发的适宜温度范围为25~30 ℃。有研究认为25~35 ℃或更高是许多热带作物的发芽温度,因为在自然环境中分散在土壤中的种子有可能经历39 ℃或更高的土壤温度(Daws et al.,2002;Asomaning et al.,2011)。
3. 2 种子发芽时胚乳中的蛋白含量会降低
在种子发芽过程中,其可溶性蛋白浓度的升高或降低是种子贮藏养分水解的生理响应结果,种子中胚和胚乳中的蛋白积累能为后期的幼苗生长提供营养,对幼苗存活率具有重要作用。桢楠种子萌发过程中,其胚乳中可溶性蛋白浓度在萌发后期降低,究其原因是在蛋白酶的作用下,蛋白分解成亚单位或氨基酸,以便提供种子萌发的能量,使其发芽成苗(龙汉利等,2013)。在本研究中,播种后第21 d,不同发芽程度的椰枣种子其胚乳中可溶性蛋白浓度明显不同,未发芽胚乳中的可溶性蛋白浓度显著高于发芽种子的胚乳,造成此结果的原因可能与桢楠种子萌发后期可溶性蛋白浓度下降的原因一致,即在椰枣发芽过程中胚和胚轴会吸收胚乳中的养分,发芽情况良好的种子其胚乳中的蛋白降解、被胚吸收,供应胚萌发形成胚轴,从而导致胚乳中的可溶性蛋白浓度降低。
3. 3 不同温度对种子内酶活性的影响存在差异
温度影响种子发芽率是通过影响种子内酶活性而影响种子内的代谢反应来实现。种子发芽时周围环境温度的变化会直接影响种子植物细胞内的酶活性和代谢作用,当周围环境的温度高于或低于一定范围时,即对种子细胞中某些蛋白结构产生不可逆的破坏,导致酶失活,致使细胞不能进行正常的生理代谢而死亡(Leverenz et al.,2010;刘润,2016)。抗氧化酶(Antioxidant enzyme)是活细胞内产生的一种可缓解细胞氧化速度的生物催化剂,也是植物代谢过程中产生的一种可调节自身生理代谢的次级代谢产物(陈博阳等,2017),不同的酶活浓度可产生不同生理效应。抗氧化酶活性与植物生长发育过程中种子萌发、细胞分化、衰老、营养物质运输和抗逆性等密切相关。抗氧化酶促系统中的重要成员包括SOD、POD和CAT,通过相互间的协同作用能将植物体内的活性氧维持在较低浓度,从而抵御外界的不良环境因素,保证植物正常的生长代谢(Wolters and Jürgens,2009;Kohli et al.,2013;张晓艳等,2017)。
在抗氧化物酶系统中,SOD属于第一道防线,其催化种子发芽过程中新陈代谢产生的超氧阴离子自由基歧化为O2和H2O2的能力强,但歧化后的H2O2能产生氧化能力更强的羟自由基和单线态分子氧,进而对细胞造成较大伤害(陈博阳等,2017)。POD和CAT能催化H2O2使其形成H2O,阻止H2O2在体内积累,从而减少活性氧对细胞的伤害(程昕昕和刘正,2012)。H2O2会对细胞产生氧化作用(刘晶等,2017),本研究中,发芽率高的椰枣白色胚轴总SOD活力显著高于发芽率低的黄色胚轴和不发芽胚乳,白色与黄色胚轴中的H2O2含量无显著差异的原因是,在适宜温度范围下发芽率高的椰枣白色胚轴POD活力显著高于发芽率低的黄色胚轴和不发芽胚乳,具备较强POD活力的白色胚轴减弱了SOD歧化后产生H2O2对细胞造成的氧化作用。
4 结论
温度是椰枣种子发芽的重要因素,在适宜的培养温度范围内,椰枣种子发芽快,发芽率高,胚轴长势好。椰枣种子发芽后胚乳中的可溶性蛋白浓度显著低于发芽前。
参考文献:
陈博阳,余彬彬,钱晓晴,王露,罗涛,张振华. 2017. 锌和土霉素胁迫对玉米种子发芽和幼苗抗氧化酶活性的影响[J]. 江苏农业学报,33(1): 13-18. [Chen B Y,Yu B B,Qian X Q,Wang L,Luo T,Zhang Z H. 2017. Zinc and oxyte-tracycline stress effects on maize germination and seedling antioxidant system[J]. Jiangsu Journal of Agricultu-ral Sciences,33(1): 13-18.]
程昕昕,刘正. 2012. 不同营养元素对甜玉米种子活力及抗氧化指标的影响[J]. 食品工业科技,33(18): 121-123. [Cheng X X,Liu Z. 2012. Effect of different nutrient elements on the seed vigor and antioxidation in sweet corn[J]. Science and Technology of Food Industry,33(18): 121-123.]
董小云,米超,刘自刚,孙万仓,武军艳,方彦,李学才,赵艳宁,刘海卿. 2018. PEG模拟水分胁迫对白菜型冬油菜幼苗生长及生长特性的影响[J]. 河南农业大学学报,52(3): 313-321. [Dong X Y,Mi C,Liu Z G,Sun W C,Wu J Y,Fang Y,Li X C,Zhao Y N,Liu H Q. 2018. Response of winter rapessed seeding growth and physiological characteristics under PEG drought tolerance[J]. Journal of Henan Agricultural University,52(3): 313-321.]
李东霞,王永,符海泉,徐中亮. 2017. 椰枣愈伤组织诱导对比分析[J]. 中国热带农业,(5): 36-39. [Li D X,Wang Y,Fu H Q,Xu Z L. 2017. Comparative analysis of date palm callus induction[J]. China Tropical Agriculture,(5): 36-39.]
李雄,尹欣,杨时海,杨云强,胡向阳,杨永平. 2014. 温度对高山植物紫花针茅种子萌发特性的影响[J]. 植物分类与资源学报,36(6):698-706. [Li X,Yin X,Yang S H,Yang Y Q,Hu X Y,Yang Y P. 2014. Effects of temperatures on seed germination characteristics of alpine plant Stipa purpurea(Poaceae)[J]. Plant Diversity and Resources,36(6):698-706.]
劉晶,张鹤婷,殷悦,陈惠萍. 2017. 外源硫化氢对干旱胁迫下萌发水稻种子抗氧化代谢的影响[J]. 南方农业学报,48(1): 31-37. [Liu J,Zhang H T,Yin Y,Chen H P. 2017. Effects of exogenous hydrogen sulfide on antioxidant metabolism of rice seed germinated under drought stress[J]. Journal of Southern Agriculture,48(1): 31-37.]
劉润. 2016. 温度和DHAP对天山云杉种子萌发和幼苗生长过程中抗氧化酶活性的影响[D]. 乌鲁木齐:新疆农业大学. [Liu R. 2016. Effects of DHAP and temperature on the antioxidant enzyme activity during seed germination and seeding growth of Picea schrenkiana[D]. Urumqi: Xinjiang Agricultural University.]
刘小金,徐大平,杨曾奖,张宁南,陈美红. 2014. 温度对越南黄花梨种子萌发的影响[J]. 林业科学研究,27(5):707-709. [Liu X J,Xu D P,Yang Z J,Zhang N N,Chen M H. 2014. Effects of temperature on seed germination of Dalbergia tonkinensis[J]. Forest Research,27(5):707-709.]
龙汉利,罗建勋,辜云杰,孙雁霞,邬晓勇. 2013. 桢楠种子萌发过程中抗氧化酶及贮藏物质的变化[J]. 西南农业学报,26(3): 978-981. [Long H L,Luo J X,Gu Y J,Sun Y X,Wu X Y. 2013. Changes of antioxidant enzymes and storage subatance during phoebe seed germination[J]. Southwest China Journal of Agricultural Sciences,26(3):978-981.]
鲁小名,魏永胜,梁宗锁. 2012. 黄芪种子萌发的基础温度与积温需要——一种模型方法[J]. 西北农业学报,21(7): 127-133. [Lu X M,Wei Y S,Liang Z S. 2012. Basal temperature and thermaltime requirement for the germination of Astragalus membranaceus seeds: A modeling app-roach[J]. Acta Agriculturae Boreali-occidentalis Sinica,21(7): 127-133.]
Parisa Eshraghi,田郎. 2011. 2个伊朗椰枣品种的体细胞胚胎发生[J]. 世界热带农业信息,(2): 25-26. [Eshraghi P,Tian L. 2011. Somatic embryogenesis of two Iranian Date Palm varieties[J]. World Tropical Agriculture Information,(2): 25-26.]
钱长江,蔡辉明,张红艳,杜勇,张文香,王宏伟,陈志萍. 2017. 不同温度和赤霉素浓度对百脉根种子萌发及植株萌蘖生长的影响[J]. 南方农业学报,48(6):997-1002. [Qian C J,Cai H M,Zhang H Y,Du Y,Zhang W X,Wang H W,Chen Z P. 2017. Effects of different temperatures and gibberellin with different concentrations on seed germination and sprout tiller growth of Lotus corniculatus L.[J]. Journal of Southern Agriculture,48(6):997-1002.]
尚瑞广,王兵益,徐珑峰. 2014. 温度、水分和光照对玛咖种子萌发的影响[J]. 西南农业学报,27(6): 2564-2568. [Shang R G,Wang B Y,Xu L F. 2014. Effects of temperature,moisture stress and light on seed germination of Maca (Lepidium meyenii Walp.)[J]. Southwest China Journal of Agricultural Sciences,27(6): 2564-2568.]
宋兆伟,郝丽珍,黄振英,李娜,赵清岩. 2009. 光照和温度对沙芥和斧翅沙芥植物种子萌发的影响[J]. 生态学报,30(10):2562-2568. [Song Z W,Hao L Z,Huang Z Y,Li N,Zhao Q Y. 2009. Effects of light and temperature on the germination of Pugionium cornutum(L.) Gaertn. and Pugionium dolabratum Maxim. seeds[J]. Acta Ecologica Sinica,30(10):2562-2568.]
唐安军,龙春林,刀志灵,宋松泉,田美华. 2005. 棕榈种子萌发特性及其贮藏行为的研究[J]. 云南植物研究,27(6): 657-662. [Tang A J,Long C L,Dao Z L,Song S Q,Tian M H. 2005. Studies on germination characteristics and storage behavior of Trachycarpus fortunei seeds[J]. Acta Botanica Yunnanica,27(6): 657-662.]
王鸿彬,刘雄盛,李志辉,肖玉菲,蒋燚,黄荣林. 2018. 温度对江南油杉种子萌发特性的影响[J]. 种子,37(1): 46-51. [Wang H B,Liu X S,Li Z H,Xiao Y F,Jiang Y,Huang R L. 2018. Effect of temperature on seed germination characteristics of Keteleeria fortuei var. cyclolepis[J]. Seed,37(1): 46-51.]
汪建军,麻安卫,汪治刚,蔡宇,旷宇,田沛. 2016. 不同温度和PEG处理对中华羊茅种子萌发的影响[J]. 草业学报,25(4):73-80. [Wang J J,Ma A W,Wang Z G,Cai Y,Kuang Y,Tian P. 2016. Effects of different temperature and moisture conditions on seed germination of Festuca sinensis[J]. Acta Prataculturae Sinica,25(4): 73-80.]
辛成齐. 2015. 椰枣microRNA鉴定及其在果实发育过程中的表达谱研究[D]. 北京:中国科学院大学. [Xin C Q. 2015. Identification of date palm(Phoenix dactylifera L.) microRNAs and their expression profiling during multi-staged fruit development[D]. Beijing: University of Chinese Academy of Science.]
徐恒恒,黎妮,刘树君,王伟青,王伟平,张红,程红焱,宋松泉. 2014. 种子萌发及其调控的研究进展[J]. 作物学报,40(7):1141-1156. [Xu H H,Li N,Liu S J,Wang W Q,Wang W P,Zhang H,Cheng H Y,Song S Q. 2014. Research progress in seed germination and its control[J]. Acta Agronomica Sinica. 40(7):1141-1156.]
张晓艳,杨忠仁,郝丽珍,张凤兰,郑清岭,郭霏,霍学婧. 2017. 温度及盐胁迫对地梢瓜种子萌发及抗氧化酶活性的影响[J]. 西北植物学报,37(6): 1166-1174. [Zhang X Y,Yang Z R,Hao L Z,Zhang F L,Zheng Q L,Guo F,Huo X J. 2017. Effect of temperature and salt stress on seed germination and antioxidant enzyme activities of Cynanchum thesioides(Freyn). K. schum[J]. Acta Agriculturae Boreali-occidentalis Sinica,37(6): 1166-1174.]
Asomaning J M,Sacande M,Olympio N S. 2011. Germination responses of Terminalia superba Engl. and Diels seeds on the 2-Way Grant’s thermogradient plate[J]. Research Journal of Seed Science,4(1): 28-39.
Daws M I,Burslem D F R P,Crabtree L M,Kirkman P,Mul-lins C E,Dalling J. 2002. Differences in seed germination responses may promote coexistence of four sympatric Piper species[J]. Functional Ecology,16(2): 258-267.
Jusaitis M,Polomka L,Sorensen B. 2004. Habitat specificity,seed germination and experimental translocation of the endangered herb Brachycome muelleri(Asteraceae)[J]. Biological Conservation,116(2): 251-266.
Kohli A,Sreenivasulu N,Lakshmanan P,Kumar P P. 2013. The phytohormone crosstalk paradigm takes center stage in understanding how plants respond to abiotic stresses[J]. Plant Cell Reports,32(7): 945-957.
Leverenz J W,Bruhn D,Saxe H,2010. Responses of two provenances of Fagus sylvatica seedlings to a combination of four temperature and two CO2 treatments during their first growing season: Gas exchange of leaves and roots[J]. New Phytologist,144(3): 437-454.
Meulebrouck K,Ameloot E,van Assche J A,Verheyen K,Hermy M,Baskin C. 2008. Germination ecology of the holoparasite cuscuta epithymum[J]. Seed Science Research,18(1): 25-34.
Orozco-Almanza M S,León-Garcı́a L P,Grether R,Garcı́a-Moya E. 2003. Germination of four species of the genus Mimosa(Leguminosae) in a semi-arid zone of Central Mexico[J]. Journal of Arid Environments,55(1): 75-92.
Qu X X,Baskin J M,Wang L,Huang Z Y. 2008. Effects of cold stratification,temperature,light and salinity on seed germination and radicle growth of the desert halophyte shrub,Kalidium caspicum(Chenopodiaceae)[J]. Plant Growth Regulation,54(3):241-248.
Wolters H,Jürgens G. 2009. Survival of the flexible: Hormonal growth control and adaptation in plant development[J]. Nature Reviews Genetics,10(5): 305-317.
(責任编辑 邓慧灵)
猜你喜欢
小天使·一年级语数英综合(2019年10期)2019-11-10
儿童时代·幸福宝宝(2019年9期)2019-10-28
智族GQ(2019年8期)2019-08-30
小天使·二年级语数英综合(2018年7期)2018-09-11
文学港(2018年1期)2018-01-25
莫愁·家教与成才(2017年7期)2017-07-11
知识就是力量(2016年9期)2016-11-02
小学阅读指南·低年级版(2016年9期)2016-05-14
少儿科学周刊·儿童版(2016年1期)2016-03-14
