
解淀粉芽胞杆菌FJAT-8754产纤维素酶和淀粉酶发酵动力学模型的构建
2019-09-10 11:25葛慈斌蓝江林刘波王凯郑雪芳
福建农业学报 2019年6期
葛慈斌 蓝江林 刘波 王凯 郑雪芳




摘 要:【目的】解淀粉芽胞杆菌菌株FJAT-8754是一株高产纤维素酶、淀粉酶的芽胞杆菌,具有在复合饲料酶制剂生产中的应用潜力。为深入了解该菌株的发酵进程,以便实现对发酵过程进行最优化控制。【方法】采用50 L发酵罐发酵菌株FJAT-8754,分析发酵过程中菌体数量、pH值、底物浓度、纤维素酶和淀粉酶等的动态变化,并选用Logistic方程和Luedeking-Piret方程构建发酵动力学模型。【结果】根据菌株FJAT-8754发酵过程中各参数变化动态建立了模拟菌体生长、纤维素酶与淀粉酶产生和底物消耗的动力学模型,并应用SPSS 16.0软件对数据进行计算与分析,Origin 9.0软件经非线性拟合与优化,获得了最佳模型参数值。【结论】所构建的动力学模型的计算值与实验值拟合良好,较好地反映了菌株FJAT-8754发酵过程中菌体生长、底物消耗和酶合成的动力学特征。菌株FJAT-8754发酵过程中产物的合成属于生长部分偶联型。
关键词:解淀粉芽胞杆菌;发酵;动力学模型;纤维素酶;淀粉酶
中图分类号:Q 938文献标识码:A文章编号:1008-0384(2019)06-697-08
Abstract: 【Objective】To construct a kinetic model for the optimized fermentation of Bacillus amyloliquefaciens FJAT-8754, which was capable of effectively producing cellulase and amylase. 【Method】A 50 L fermenter experiment was conducted to study the cellular growth,pH value,enzyme production and carbohydrate consumption of the fermentation and for subsequent process optimization. A kinetic model was established using the logistic and Luedeking-Piret equations, and experimental data calculated and analyzed with SPSS 16.0 software. 【Result】 The optimal fermentation conditions fitted well in the non-linear model as shown by Origin 9.0 software. 【Conclusion】The kinetic model obtained accurately reflected the performance of the fermentation. The cellulase and amylase were products of FJAT-8754 metabolism in the culture.
Key words:Bacillus amyloliquefaciens; fermentation; kinetic model; cellulase; amylase
0 引言
【研究意义】解淀粉芽胞杆菌 Bacillus myloliquefaciens是芽胞杆菌科Bacillaceae芽胞杆菌属Bacillus的一种革兰氏阳性杆菌,由于其能产生液化淀粉酶分解淀粉,而被定为该名[1]。解淀粉芽胞杆菌在自然界广泛存在,许多菌株可以产生一系列能够抑制真菌和细菌生长的代谢产物及多种水解酶,因此具有防治植物病害、促进动植物生长、水果保鲜等功能[2-4]。有较多学者对从自然界分离到的解淀粉芽胞杆菌进行了发酵方面的研究,期望将其应用于植病生防、动物饲养、环境保护等领域。王卉等[5]用正交试验法优化了具有植物促生长效果的解淀粉芽胞杆菌L-S60的固体发酵工艺,可以使发酵产物中的菌体数达到2.42×1010 cfu·g-1;郦金龙等通过响应面分析法,确定了解淀粉芽胞杆菌菌株11568液体发酵产柚苷酶的最优培养基成分和发酵条件 [6];王全等[7]对解淀粉芽胞杆菌菌株12-7产抗菌蛋白的发酵条件进行了优化,为将该菌株应用于棉花黄萎病生物农药的研制开发奠定了基础。
【前人研究进展】发酵动力学研究微生物发酵过程中菌株生长速率、培养基消耗速率和产品形成速率间的相互作用和随时间变化的规律,是对微生物生长和产物合成过程的定量描述。目前,关于解淀粉芽胞杆菌产纤维素酶和淀粉酶发酵动力学的研究未见报道,但其他芽胞杆菌发酵动力学研究有过许多的报道。欧志敏等阐明了蜡样芽胞杆菌Bacillus cereus菌株CGMCC No.12336分批发酵过程的动力学模型,并拟合得到了动力学参数 [8];于晓萌等[9]优化了巨大芽胞杆菌Bacillus megaterium菌株GJT321生产Levan果聚糖的发酵条件,并研究了该菌株发酵生产Levan果聚糖的数字化模型;田雪等[10]基于Logistic方程和Luedeking-Piret等方程建立了地衣芽胞桿菌Bacillus licheniformis产β-甘露聚糖酶分批发酵动力学模型;孙宏等[11]优化了枯草芽胞杆菌Bacillus subtilis菌株Z-4产纤维素酶的培养条件,探索了该菌株发酵产酶过程中细胞生长和纤维素酶产物积累的规律,建立了相关数学模型;马永强等[12]建立了地衣芽胞杆菌菌株2709产碱性蛋白酶的发酵动力学模型;刘朝辉等[13]构建了能够较好表征枯草芽胞杆菌TJ-200603分批发酵过程中菌体细胞生长、产物β-甘露聚糖酶合成以及基质总糖消耗的3个动力学方程,这些都为各菌株发酵过程的在线控制和预测提供了理论基础。
【本研究切入点】解淀粉芽胞杆菌菌株FJAT-8754是本研究所筛选到的具有高产纤维素酶和淀粉酶复合酶的菌株,为了通过发酵获得大量的菌体及高活力的复合酶制剂产品,王凯等前期对菌株的发酵条件进行了优化[14]。【擬解决的关键问题】为了从数学层面去分析、揭示菌株发酵过程中菌体数量、底物消耗、酶产生的动态变化,为该菌株的中试扩大培养以及工业化生产提供理论基础,作者进行了菌株的发酵动力学研究。
1 材料与方法
1.1 菌株与培养基
菌株:解淀粉芽胞杆菌菌株FJAT-8754,保存于福建省农业科学院农业生物资源研究所菌种库;培养基:菌株FJAT-8754的一级种子液培养基为Luria-Bertani(LB)培养基,二级种子液培养基的组成为玉米淀粉10 g、豆粕粉10 g、NaCl 5.0 g、MgSO4·7H2O 0.5 g、KH2PO4·2H2O 0.5 g、K2HPO4·2H2O 0.5 g、CaCl2 0.2 g,蒸馏水1 000 mL,pH值6.2,上罐发酵培养基同二级种子液培养基。
1.2 菌株FJAT-8754的发酵培养
活化菌株FJAT-8754,并分别制备一级、二级种子液。应用50 L发酵罐(GUJS-50L型,镇江东方生物工程设备技术公司),参照葛慈斌等[15]的方法,配制发酵培养基30 L、灭菌、接种,设定罐温35℃、通气量30 L·min-1、转速200 r·min-1,进行发酵培养,并每隔4 h 记录发酵罐中发酵液的pH值、DO值,并取样300 mL分装成3份,用于发酵液中活菌数、纤维素酶与淀粉酶活力、总糖、还原糖等状态参数的测定。共进行3次发酵试验,取上述状态参数平均值进行发酵动力学分析。
1.3 发酵液的活菌数测定
采用稀释涂布平板法测定发酵液的活菌数,取×10-4、×10-5、×10-6等3个梯度的稀释液100 μL,均匀涂布在LB培养基平板(每个浓度重复3次),放置在30℃恒温培养箱内培养,3 d后统计平板上的菌落数,计算各发酵液的菌体数。
1.4 纤维素酶、淀粉酶活力的测定
取不同时段的发酵液5 mL,6 000 r·min-1离心10 min,上清液即为粗酶液,置于4℃冰箱备用;参照王凯等[14]的方法,用3,5-二硝基水杨酸(3,5-dinitrosalicylic acid,DNS)显色法测定发酵液中纤维素酶、淀粉酶的活力。
1.5 发酵液总糖、还原糖含量的测定
还原糖含量测定:采样3,5-二硝基水杨酸法[16]测定,取1 mL发酵液于50 mL容量瓶中并用蒸馏水定容,取定容后的l mL稀释液于20 mL的比色管中,加入1.5 mL DNS溶液,混匀后沸水浴10 min,冷却至室温后定容至20 mL,于520 nm波长处测吸光度,计算出发酵液中的还原糖含量。总糖含量测定:取5 mL发酵液于100 mL锥形瓶中,加蒸馏水15 mL和6 mol·L-1盐酸15 mL,混匀,在沸水浴中加热30 min后,用碘化钾-碘溶液检测水解程度,至完全水解;冷却后,加入甲基红指示液两滴,用10% NaOH溶液中和至红色消失,过滤并定容至100 mL;取定容后的溶液1 mL,按照还原糖的测定方法测定发酵液中的总糖含量。
1.6 动力学模型的构建
1.6.1 菌株FJAT-8754的生长动力学模型
在描述发酵过程中菌体细胞生长时,最常用的是Logistic方程(阻滞方程),该方程能较好描述菌体的生长规律。Logistic方程为:
式中:μm为最大比生长速率,h-1;X为菌体浓度,×107 cfu ·mL-1;X0为初始菌体浓度,×107 cfu ·mL-1;Xm为最大菌体浓度,×107 cfu ·mL-1;t为时间,h。对方程(1)积分可得菌体浓度X的时间函数为:
以方程(2)为自定义函数,利用Origin 9.0软件对菌株FJAT-8754的生长进行非线性拟合,即可得到该菌株生长的动力学模型。
1.6.2 菌株FJAT-8754的纤维素酶、淀粉酶合成动力学模型
微生物发酵过程中除产生菌体以外,还会形成次级代谢产物。Elmer等[17]把代谢产物的形成方式分为三类,即生长相关型(也叫生长偶联型)、部分生长相关型(部分偶联型)和非生长相关型(非偶联型)。Luedeking-Piert方程常用于描述发酵产物合成动力学,当发酵产物的产生同时存在非生长相关型和生长相关型时,用下列公式表示:
式中:P为纤维素酶或淀粉酶的酶活,U ·mL-1;α为与生长相关联的产物合成系数;β为与生长无关的产物合成比速率。
将公式(2)带入公式(3)并积分,整理得到公式(4):
将已知X0、Xmax、μmax拟合值代入公式(4),应用Origin 9.0软件进行非线性自定义函数拟合,使用Levenberg-Marguardt参数拟合工具箱,分别进行拟合求参数α、β,将参数带入公式(4),即得解淀粉芽胞杆菌FJAT-纤维素酶、淀粉酶合成动力学模型:
(1)纤维素酶合成动力学模型
(2)淀粉酶合成动力学模型
1.6.3 菌株FJAT-8754的碳源消耗动力学模型
在菌株FJAT-8754发酵过程中,玉米淀粉作为唯一碳源,主要用于产生新的菌体、维持菌体基本生命活动和代谢产物的生成这3个方面;但在发酵过程中,维持菌体基本生命活动所需消耗的底物所占比例相对较小,经常被忽略不计,为方便建模,可将该菌株发酵过程中玉米淀粉的消耗分为菌体生长消耗和产物合成消耗2部分,因此底物消耗方程式可用类似于Luedeking-Piert模型,具体公式如下:
式中:S为碳源(玉米淀粉)的质量浓度,mg ·mL-1;YX/S为细胞对碳源的得率常数;YP/S为酶活对碳源的得率常数。
将公式(2)、(3)代入公式(7),并进行积分处理得:
将纤维素酶、淀粉酶合成动力学公式(5)、(6)代入(8)整理得:
其中:α1為纤维素酶合成系数,β1为产物合成比速率(对纤维素酶),Y1为纤维素酶活产物与基质的得率常数;α2为淀粉酶合成系数,β2为产物合成比速率(对淀粉酶);Y2为淀粉酶酶活产物与基质的得率常数;S0为培养初期的初始基质浓度,μg ·mL-1。
利用Origin 9.0软件,采用Levenberg-Marguardt参数拟合工具箱,对菌株FJAT-8754发酵过程中基质消耗进行拟合,可得底物(总糖)消耗的动力学模型。
2 结果与分析
2.1 解淀粉芽胞杆菌FJAT-8754的发酵特征
解淀粉芽胞杆菌菌株FJAT-8754在50 L发酵罐培养过程中,随着菌株的生长,发酵液中的DO值、pH值、菌体浓度、还原糖浓度等参数也相应发生着一系列的动态变化(图1)。从图1可以看出,菌株FJAT-8754接种4 h后进入对数生长期,并在24 h时进入稳定期,28 h时菌体浓度达到最大值1.30×109 cfu ·mL-1,此后菌体浓度缓慢下降;纤维素酶、淀粉酶的酶活在刚开始发酵时极低,随着菌株进入对数生长期,酶活也随之迅速升高,并在32 h时(略迟于菌体浓度达到最大值的时间)达到最大酶活,酶活分别为125.8、1 267.7 U ·mL-1。
菌株FJAT-8754发酵液的溶氧量(DO)在开始培养后便快速下降,在16 h时达到最低、只有18.2,此后随着底物的消耗和菌株生长进入稳定期、衰亡期,发酵液的DO值逐渐上升。发酵液的pH在发酵2 h后开始快速下降,至发酵12 h时达到最低点5.22,这可能是因为菌体快速生长大量消耗葡萄糖产酸的缘故;之后pH值开始缓慢上升,至发酵36 h时升至6.17。
发酵液的总糖含量在0~4 h几乎没变化,之后随着菌株进入对数生长期,菌体快速生长需要消耗大量的碳源而开始迅速降低,至发酵20 h后由于菌体增长速度放缓,总糖消耗变小,浓度缓慢降低至基本保持稳定。还原糖浓度在0~4 h变化不大,发酵4 h后随着大量总糖被水解,还原糖浓度开始较快上升,并于16 h时达到峰值2.53 mg ·mL-1,之后由于总糖水解的减少,发酵液中的还原糖浓度又开始下降,至发酵36 h时降至1.92 mg ·mL-1。
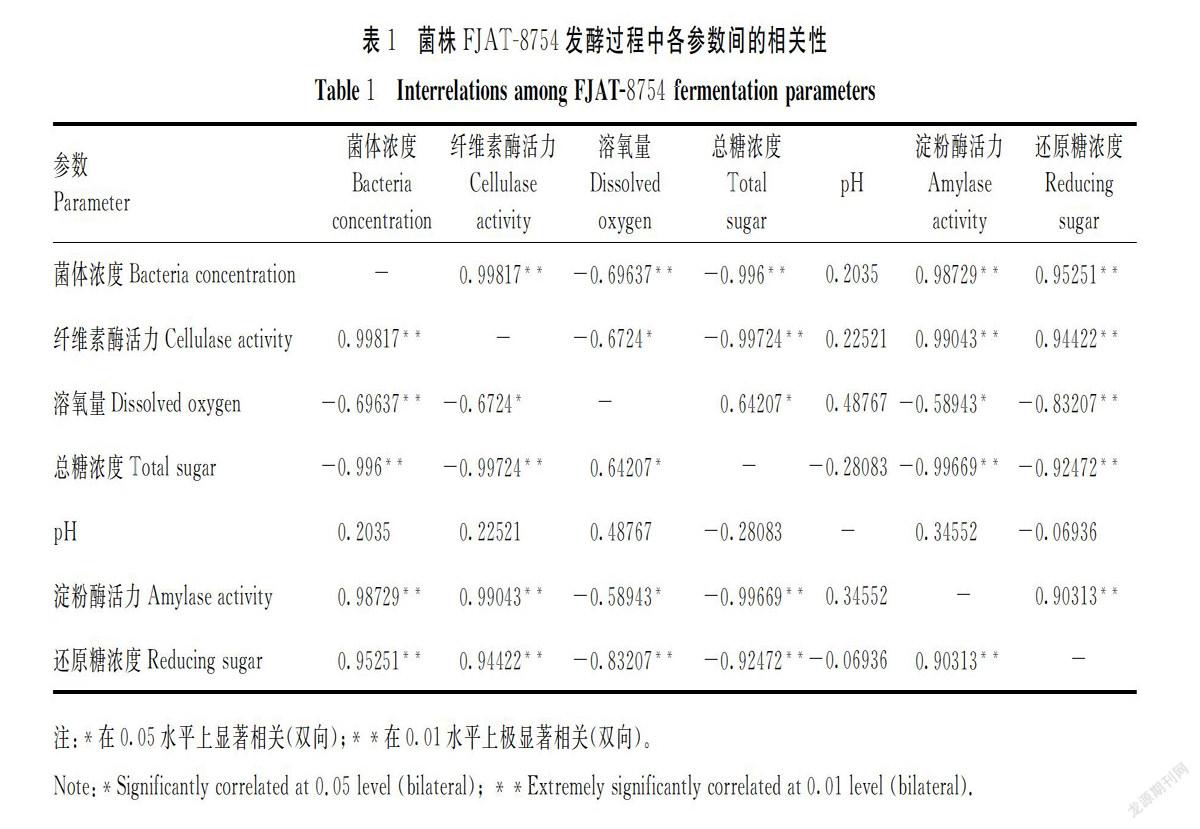
分析菌株FJAT-8754发酵过程中,发酵液的菌体浓度、DO值、pH值、纤维素酶活力、淀粉酶活力、总糖浓度和还原糖浓度等7个参数之间的相关性(表1),可以看出,pH值与其他6个参数之间的相关性均不显著;菌体浓度与除pH值之外的另5个参数之间存在着极显著的相关性,其中与DO、总糖浓度之间是负相关;纤维素酶活力与总糖浓度、淀粉酶活力及还原糖浓度之间的相关性也极显著,与DO之间的相关性显著;淀粉酶活力与总糖浓度、还原糖浓度、DO之间的相关性与纤维素酶活性基本一致;总糖浓度与DO之间显著相关,与还原糖浓度之间则极显著负相关;还原糖浓度与DO之间也存在着极显著的负相关关系。
2.2 解淀粉芽胞杆菌FJAT-8754生长动力学
解淀粉芽胞杆菌菌株FJAT-8754在50 L发酵罐的培养过程中,菌体细胞在经过短暂的延迟期后,进入对数生长期,之后趋于稳定(图1)。用Logistic方程描述菌株生长过程,用origin软件对试验所得的数据进行非线性拟合,得到了该菌株生长的动力学模型(10),拟合模型曲线(图2)R2=0.99766,且试验值都在95%的置信区间内,模型预测值与实测值能较好的拟合。
2.3 解淀粉芽胞杆菌FJAT-8754的纤维素酶、淀粉酶合成动力学
由菌株FJAT-8754发酵过程参数测定(图1)可以看出,纤维素酶、淀粉酶的活性随菌体的增长而增加,达到稳定期后这两种酶才达到最高酶活,表明这两种酶的形成方式为部分偶联型。运用Luedeking-Piert公式计算得到的菌株FJAT-8754产纤维素酶、淀粉酶合成动力学模型分别为公式(5)和(6);产纤维素酶的拟合模型曲线见图3,R2=0.99657;产淀粉酶的拟合模型曲线见图4,R2=0.99298。
2.4 菌株FJAT-8754的底物(总糖)消耗动力学模型
用类似于Luedeking-Piert模型计算得到的菌株FJAT-8754发酵过程中底物(总糖)消耗的动力学模型见公式(11),拟合模型曲线见图5,R2=0.98905。
3 讨论与结论
菌株FJAT-8754发酵过程中,菌体生长、纤维素酶与淀粉酶合成、底物消耗互为联系,密不可分。菌株FJAT-8754纤维素酶、淀粉酶活力的增长趋势滞后于菌体细胞生长,即产酶峰值滞后于菌体浓度峰值,这与郑毅等[18]、徐莹等[19]的研究结果较相似,即菌体细胞生长进入稳定期后,发酵体系中纤维素酶、淀粉酶的生产量仍在增长,酶活力继续提高,由此认为菌株FJAT-8754发酵生产纤维素酶、淀粉酶分为菌体生长和产物合成两个阶段,产物合成属于部分偶联型。
发酵动力学是微生物发酵过程优化和控制的重要基础,研究发酵动力学有助于深入了解微生物的代谢规律,从而有利于建立合理的发酵工艺,或是优化已有的发酵过程 [20];Monod、Aiba、Logistic和Luedeking-Piret等函数常用于发酵动力学模型的构建[21-22]。李浩等[23]对生防菌吡咯伯克霍尔德氏菌菌株JK-SH007液体分批发酵产生抑菌活性的动力学进行了研究,运用非线性拟合的方法确立了该菌株分批发酵动力学模型。Zhang等[24]建立了酿酒酵母菌株D254发酵过程中酵母菌生长、乙醇产生和底物消耗的动力学模型,有助于及时了解发酵液中乙醇及残糖的浓度。赵健烽等[25]以Logistic、Luedeking-Piret等方程为基础,建立了铜绿假单胞菌菌株M14808发酵过程中菌体生长、鼠李糖脂合成和碳源消耗的动力学模型,拟合度较好;周有彩等[26]构建了50 L发酵罐小球藻分批培养的Logic和Ludeking-Piret动力学模型,可用于描述小球藻分批培养细胞生长、蛋白质合成和葡萄糖消耗的过程;潘丹阳等[27]通过Logistic方程构建了层生镰孢菌细胞生长、几丁质脱乙酰酶(CDA)合成及糖基质消耗的非结构动力学模型;聂国兴等[28]基于Logistic和Luedeking-Piret,根据分批发酵过程中菌体生长、总糖消耗、反应体系pH值变化规律等构建枯草芽胞杆菌E79发酵过程中菌体细胞生长、产物合成及基质消耗的动力学模型,并应用Oirgin7.5软件经非线性拟合与优化,获得了最佳模型参数值,能较好反映菌株E79发酵的动力学特征。
本研究使用Origin 9.0软件对解淀粉芽胞杆菌菌株FJAT-8754产纤维素酶、淀粉酶复合酶发酵过程进行分析处理,建立了该菌株发酵过程中菌体生长、纤维素酶与淀粉酶合成以及底物消耗的动力学模型。经过模型拟合,发现这些模型与实验结果拟合较好,R2都在0.989以上,说明这些模型可较准确地描述发酵过程中菌株FJAT-8754菌体生长、纤维素酶与淀粉酶合成、基质消耗的变化规律。
参考文献:
[1]LIU W X, WANG X B, WU L H, et al. Isolation, identification and characterization of Bacillus amyloliquefacuens BZ-6, abacterial isolated for enhancing oil recovery from oily sludge[J]. Chemosphere, 2012, 87(10): 1105-1110.
[2]胡忠亮, 郑催云, 田兴一, 等. 解淀粉芽胞杆菌在环境保护和农业生产中的应用[J]. 农药, 2016, 55(4): 241-245.
HU Z L, ZHENG C Y, TIAN X Y, et al. Application of Bacillus amyloliquefaciens in environmental protection and agriculture production[J]. Agrochemicals, 2016, 55(4): 241-245.(in Chinese)
[3]栾素军, 邢焕, 孙永波, 等. 解淀粉芽胞杆菌在畜禽养殖中的应用研究进展[J]. 中国畜牧兽医, 2016, 43(10): 2615-2620.
LUAN S J, XING H, SUN Y B, et al. Research progress on Bacillus amyloliquefaciens in animal breeding industry[J]. China Animal Husbandry and Veterinary Medicine, 2016, 43(10): 2615-2620.(in Chinese)
[4]陆燕, 李澄, 陈志德, 等. 解淀粉芽胞杆菌41B-1对花生白绢病的生防效果[J]. 中国油料作物学报, 2016, 38(4): 487-494.
LU Y, LI C, CHEN Z D, et al. Biological control activities of Bacillus amyloliquefaciens 41B-1 against Sclerotium rolfsii[J]. Chinese Journal of Oil Crop Sciences, 2016, 38(4): 487-494.(in Chinese)
[5]王卉, 游成真, 秦宇轩, 等. 解淀粉芽胞杆菌L-S60生物学特性及其固态发酵工艺研究[J]. 中国农业大学学报, 2016, 21(9): 133-142.
WANG H, YOU C Z, QIN Y X, et al. The biological characteristics and solid-state fermentation of Bacillus amyloliquefaciens L-S60[J]. Journal of China Agricultural University, 2016, 21(9): 133-142.(in Chinese)
[6]郦金龙, 朱运平, 腾超, 等. 解淀粉芽胞杆菌11568产柚苷酶发酵条件的优化[J]. 中国食品学报, 2016, 16(7): 80-89.
LI J L, ZHU Y P, TENG C, et Optimization of fermentation conditions for naringinase by Bacillus amyloliquefaciens 11568[J]. Jouanal of Chinese Institute of Food Science and Technology, 2016, 16(7): 80-89.(in Chinese)
[7]王全, 王占利, 高同國, 等. 响应面法对解淀粉芽胞杆菌( amyloliquefaciens)12-7产抗菌蛋白条件的优化[J]. 棉花学报, 2016,28(3): 283-290.
WANG Q, WANG Z L, GAO T G, et al. Antagonistic protein producing condition optimization of antagonistic strain Bacillus amyloliquefaciens 12-7 against Verticillium dahlia Kieb. using response surface methodology[J]. Cotton Science, 2016,28(3): 283-290.(in Chinese)
[8]欧志敏, 马兰. 蜡样芽胞杆菌发酵产脂肪酶的分批发酵动力学[J]. 发酵科技通讯, 2017, 46(1): 1-5.
OU Z M, MA L. Kinetics models of batch fermentation of Bacillus cereus CGMCC No. 12336 for lipase production[J]. Bulletin of Fermentation Science and Technology, 2017, 46(1): 1-5.(in Chinese)
[9]于晓萌, 张京良, 孙永超, 等. 基于发酵优化和动力学建立Levan果聚糖生产的数字化模型[J]. 食品与发酵工业, 2016,42 (12): 20-25.
YU X M, ZHANG J L, SUN Y C, et al. Establishment of digital model for levan production based on fermentation optimization and kinetics[J]. Food and Fermentation Industries, 2016,42 (12): 20-25.(in Chinese)
[10]田雪, 解鑫, 周晓杭, 等. 地衣芽胞杆菌产β-甘露聚糖酶分批发酵动力学模型的建立[J]. 中国农学通报, 2013, 29(18): 193-199.
TIAN X, XIE X, ZHOU X H, et al. Construction of kinetics model for β-mannase fermentation process using Bacillus licheniformis HDYM-04[J]. Chinese Agricultural Science Bulletin, 2013, 29(18): 193-199.(in Chinese)
[11]孙宏, 姚晓红, 吴逸飞, 等. 产纤维素酶枯草芽胞杆菌的培养条件优化及发酵模型构建[J]. 中国食品学报, 2012, 12(5): 76-81.
SUN H, YAO X H, WU Y F, et al. Optimization of growth conditions of a cellulose-producing Bacillus subtilis atrain and its growth model construction[J]. Journal of Chinese Institute of Food Science and Technology, 2012, 12(5): 76-81.(in Chinese)
[12]馬永强, 尹永智, 杨春华, 等. 地衣芽胞杆菌2709产碱性蛋白酶的发酵动力学研究[J]. 食品工业科技, 2010, 31(6): 159-161, 164.
MA Y Q, YIN Y Z, YANG C H, et al. Study on fermentation kinetics of production of alkaline protease by B. licheniformis 2709[J]. Science and Technology of Food Industry, 2010, 31(6): 159-161, 164.(in Chinese)
[13]刘朝辉, 陈云, 齐崴, 等. 中性β-甘露聚糖酶分批发酵动力学研究[J]. 化学工程, 2008, 36(10):66-70.
LIU Z H, CHEN Y, QI W, et al. Kinetics of neutralβ-mannanase batch fermentation[J]. Chemical Engineering (China), 2008, 36(10):66-70.(in Chinese)
[14]王凯, 蓝江林, 刘波, 等. 解淀粉芽胞杆菌FJAT-8754产纤维素酶和淀粉酶特性及发酵条件优化[J]. 福建农业学报, 2014, 29(4): 357-363.
WANG K, LAN J L, LIU B, et al. The characteristics of cellulose and amylase from B. amyloliquefaciens FJAT-8754 and optimization of fermentation conditions[J]. Fujian Journal of Agricultural Sciences, 2014, 29(4): 357-363.(in Chinese)
[15]葛慈斌, 刘波, 肖荣凤, 等. 生防菌哈茨木霉FJAT-9040深层发酵的生长适合度分析[J]. 福建农业学报, 2013, 28(10): 1032-1038.
GE C B, LIU B, XIAO R F, et al. Submerged fermentation of Trichoderma harzianum, FJAT-9040[J]. Fujian Journal of Agricultural Sciences, 2013, 28(10): 1032-1038.(in Chinese)
[16]黄翊鹏, 祁静, 郑亚军, 等. 3,5-二硝基水杨酸法测定椰子纳塔发酵液中总糖含量研究[J]. 广东农业科学, 2009, 36(12): 171-174.
HUANG Y P, QI J, ZHENG Y J, et al. Study on colorimetric determination of water-soluble total sugar in Nata de coco fermentation broth with 3,5-dinitrosalicylic acid[J]. Guangdong Agricultural Sciences, 2009, 36(12): 171-174.(in Chinese)
[17]ELMER L, GADEN J R. Fermentation process kinetics[J]. Journal of Biochemical and Microbiological Technology and Engineering, 1959, 7(4): 413-429.
[18]郑毅, 张志国, 关雄. 苏云金芽胞杆菌蛋白酶发酵动力学模型的构建[J]. 生物数学学报, 2008, 23(4): 727-734.
ZHENG Y, ZHANG Z G, GUAN X. Kinetics model for protease of Bacillus thuringiensis by liquid fermentation[J]. Journal of Biomathematics, 2008, 23(4): 727-734.(in Chinese)
[19]徐莹, 何国庆. 弹性蛋白酶分批发酵动力学模型的建立[J]. 农业生物技术学报, 2007, 15(1): 138-141.
XU Y, HE G Q. Dynamic models for elastase batch fermentation process[J]. Journal of Agricultural Biotechnology, 2007, 15(1): 138-141.(in Chinese)
[20]许晓鹏, 郑志永, 朱莉, 等. Sphingomonas sp. 生产威兰胶分批发酵动力学[J]. 食品与生物技术学报, 2015, 34(5): 507-511.
XU X P, ZHENG Z Y, ZHU L, et al. Study on welan gum fermentation dynamics of Sphingomonas sp. [J]. Journal of Food Science and Biotechnology, 2015, 34(5): 507-511.(in Chinese)
[21]KSSBAWATI, RUSNI S, SULFAHRI, et al. Determining an appropriate unstructured kinetic model for batch ethanol fermentation data using a direct search method[J]. Biotechnology & Biotechnological Equipment, 2018,32(5): 1167-1173.
[22]WANG S, LIN H, RU B, et al. Kinetic modeling of biomass components pyrolysis using a sequential and coupling method[J]. Fuel, 2016, 185: 763-771.
[23]李浩, 任嘉紅, 叶建仁, 等. 吡咯伯克霍尔德氏菌JK-SH007分批发酵动力学模型的构建[J]. 南京林业大学学报(自然科学版), 2016, 40(2): 173-178.
LI H, REN J H, YE J R, et al. Construction of batch fermentation kinetics model of Burkholderia pyrrocinia strain JK-SH007[J]. Journal of Nanjing Forestry University (Natural Sciences Edition), 2016, 40(2): 173-178.(in Chinese)
[24]ZHANG G F, LI X L, CHEN W X, et al. Organic acid content, antioxidant capacity, and fermentation kinetics of matured coconut (Cocos nucifera) water fermented by Saccharomyces cerevisiae D254[J]. International Journal of Food Engineering, 2018. DOI: 10.1515/ijfe-2017-0331.
[25]赵健烽, 辛兴, 卫培培, 等. 铜绿假单胞菌M14808产鼠李糖分批发酵动力学研究[J]. 南京理工大学学报, 2013, 37(2): 309-313.
ZHAO J F, XIN X, WEI P P, et al. Kinetics of batch fermentation of rhamnolipid by Pseudomonas aeruginosa M14808[J]. Journal of Nanjing University of Science and Technology, 2013, 37(2): 309-313.(in Chinese)
[26]周有彩, 何勇锦, 李林声, 等. 基于发酵动力学模型的小球藻高密度发酵培养[J]. 过程工程学报, 2018, 18(3): 624-631.
ZHOU Y C, HE Y J, LI L S, et al. High cell density fermentation of Chelorella based on kinetics model[J]. The Chinese Journal of Process Engineering, 2018, 18(3): 624-631.(in Chinese)
[27]潘丹阳, 刘帅, 万芳芳, 等. 层生镰孢菌产甲壳素脱乙酰酶发酵动力学[J]. 菌物学报, 2018, 37(9): 1207-1214.
PAN D Y, LIU S, WAN F F, et al. Fermentation kinetics of chitin deacetylase production by Fusarium proliferatum[J]. Mycosystema, 2018, 37(9): 1207-1214.(in Chinese)
[28]聂国兴, 明红, 宋东蓥, 等. 枯草芽胞杆菌E79分批发酵动力学[J]. 化学工程, 2011, 39(11): 6-10.
NIE G X, MING H, SONG D Y. Kinetics of xylanase batch fermentation by Bacillus subtilis E79[J]. Chemical Engineering (China), 2011, 39(11): 6-10.(in Chinese)
(责任编辑:林海清)
猜你喜欢
家庭医药(2022年7期)2022-07-20
人人健康(2017年11期)2017-06-02
科技创新导报(2016年21期)2016-12-17
湖北农业科学(2016年18期)2016-12-08
科教导刊·电子版(2016年26期)2016-11-21
科技视界(2016年11期)2016-05-23
科技视界(2015年25期)2015-09-01
科技传播(2009年17期)2009-10-28
现代农业科技(2009年19期)2009-03-20
中学生物学(2008年8期)2008-10-27
