
六倍体披碱草属的核型分析
2019-09-17 09:21杨财容张海琴刘松青沙莉娜周永红
四川农业大学学报 2019年4期
杨财容,张海琴,刘松青,沙莉娜,周永红*
(1.成都师范学院化学与生命科学学院,成都 611130;2.四川农业大学小麦研究所,成都 611130)
染色体是物种基因的载体。物种进化过程中的染色体结构与数量变化带来基因的重新排列、基因数量的增加或减少,染色体结构与数量变化也是造成群体间生殖隔离的一个重要原因,进而形成新物种[1]。G.A.Levitsky[2]首次将核型定义为体细胞染色体的表型特征,即核型是指某种真核生物个体或某一分类群(种、亚种或变种、居群)的染色体数目、长度、臂比、着丝粒位置、有无随体等形态特征的总和。染色体核型分析是细胞遗传学研究中的一个基本方法,即通过观察物种染色体形态特征,并按照一定的标准,人为地对物种染色体进行配对、编号和分组的过程。它易操作、经济实惠的特点使其得到了广泛使用,对研究物种起源与演化、物种分类、物种之间亲缘关系以及植物远缘杂交育种中的染色体鉴别发挥了重要作用[3-5]。
披碱草属是小麦族中的一个大属,包含了从四倍体到十倍体的物种,但是很多物种还缺乏生物系统学的详细资料。刘玉红、蔡联炳和冯海生等[6-7]发现 E.atratus、E.breviaristatus、E.cylindricus 等物种的核型,但是结果显示部分物种的核型不一致。六倍体披碱草属物种分布广泛,物种不同居群的核型可能存在变异。本研究分析了9 个六倍体披碱草属物种的核型,探讨物种不同居群的核型变异,为其后续理论研究和育种利用提供基础。
1 材料和方法
1.1 供试材料
本研究涉及六倍体披碱草属物种9 个。所有材料的物种名称、编号及来源列于表1。PI 编号材料由美国国家植物种质资源库American National Plant Germplasm System(Pullman,Washington,USA)提供,其余材料由四川农业大学小麦研究所采集。所有材料均种植于四川农业大学小麦研究所多年生种质圃,凭证标本藏于四川农业大学小麦研究所标本室(SAUTI)。
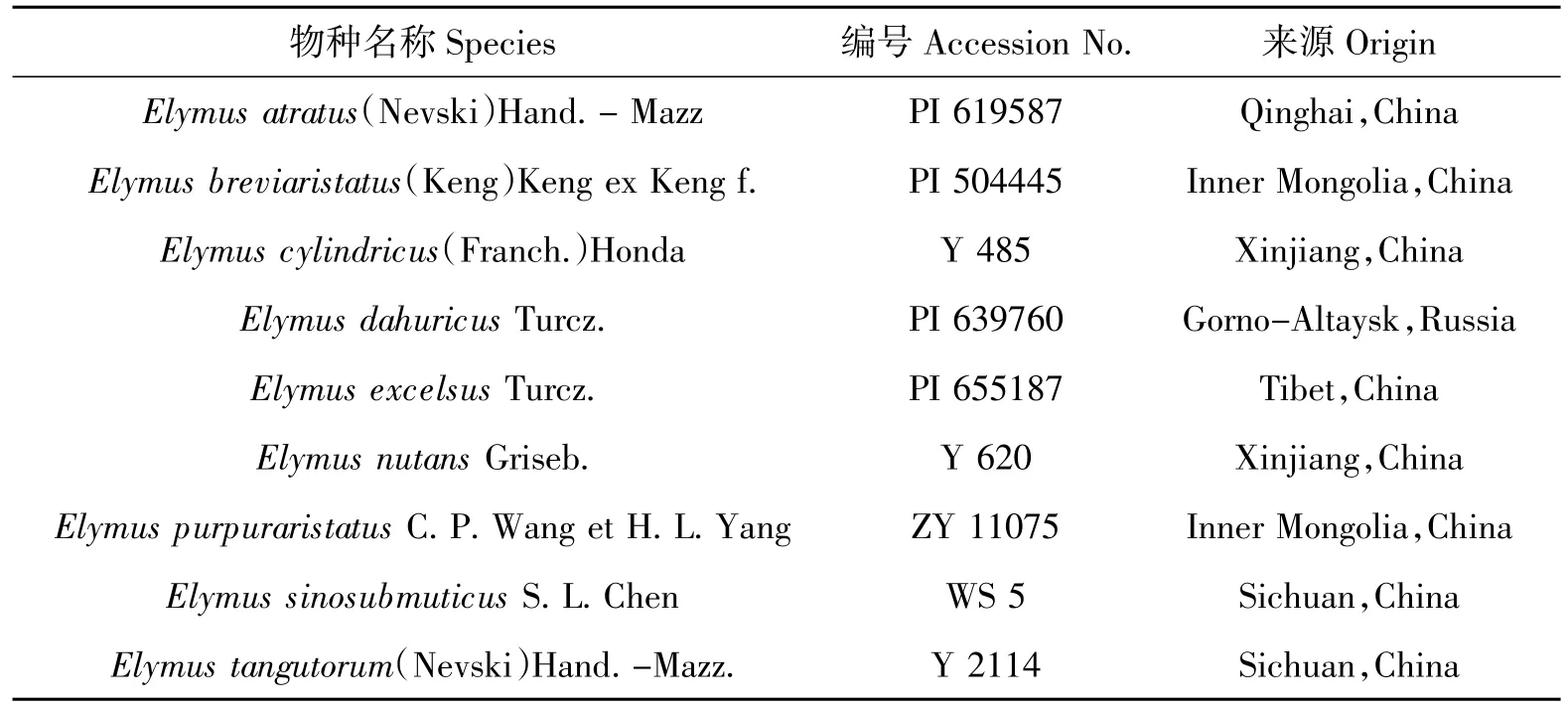
表1 供试材料Table 1 Materials used in the study
1.2 试验方法
1.2.1 根尖获取
取饱满的种子置于带有湿润滤纸的培养皿上,4 ℃解除休眠后, 置于25 ℃恒温培养箱培养发芽,根长到一定长度后移栽于盆中。待种子根尖长到1~2 cm 时于上午或下午的合适时期剪取,将剪取的根尖置于空的离心管内置于N2O 中预处理2 h 左右,取出后用90%冰醋酸固定5 min 后,于70%酒精中保存。
1.2.2 滴片法制片
将于70%酒精中保存的3~5 个根尖用蒸馏水清洗3~5 次,用刀片切取分生区,将切下的部分转入装有10 μL 酶解液(含纤维素酶和果胶酶)的0.1 mL的EP 管中,再将其置于37 ℃的恒温水浴锅中酶解14 min 左右(酶解时间取决于根尖分生区的大小、老嫩等)。酶解结束后弃掉大部分的酶解液,然后用70%的酒精清洗两次,再用解剖针轻轻捣碎组织,离心后弃掉液体,加入约30 μL 的100%冰醋酸,轻轻混匀后滴片。待载玻片自然干燥后滴加改良苯酚品红染液染色,盖上盖玻片在Olympus BX-51(Olympus)下观察并拍照。
酶解液的制备方法为:
①0.01 mol/L 柠檬酸缓冲液(Citrate buffer,0.01 mol/L,pH 值 4.5)的配制:分别称取 1.47 g 二水合柠檬酸钠(Na3C6H5O7·2H2O)和水合柠檬酸(C6H8O7·H2O)1.05 g 溶解于 500 mL ddH2O 中;
②10 mL的0.01mol/L 柠檬酸缓冲液中加入400mg纤维素酶(cellulase)和 200 mg 果胶酶(pectinase),充分溶解后,分装并保存在-20 ℃冰箱中。
1.2.3 染色体核型分析
核型分析采用A.Levan 等[8]的两点四区系统法确定着丝点类型,核型分类依照G.L.Stebbins[9]的分类标准进行,同时参照李懋学等[10]的植物核型分析标准,核型不对称系数按照H.Arano[11]的方法计算。每个物种选取5 个分裂良好的细胞,利用Nuc-Type 2.0(四川大学)进行染色体长度测量,利用该软件可直接输出染色体参数以及核型模式图。基于得到的染色体参数利用Excel 软件对核型数据进行统计,并对同源染色体进行配对。最后,利用Photoshop 软件,按照染色体的相对长度由长至短(短臂在上,长臂在下)的顺序排列,着丝粒对齐排列于同一直线上,绘制核型图。
2 结果与分析
9 个六倍体披碱草属物种的染色体形态、 核型图和核型模式图见图1,核型分析结果见表2。
E.atratus:核型公式为 2n=6x=42=38m+4sm。42条染色体中有4 条染色体为近中部着丝粒染色体,38 条为中部着丝粒染色体,无随体染色体。染色体相对长度介于3.44%~6.33%之间,染色体长度比为1.95。染色体臂比范围为 1.02~2.07 之间,平均臂比为1.28,臂比大于2 的比例为4.76%。染色体不对称系数为56.06%,核型为2A 型。
E.breviaristatus:核型公式为 2n=6x=42=34m+8sm。42 条染色体中8 条染色体为近中部着丝粒染色体,34 条中部着丝粒染色体,无随体染色体。染色体相对长度为3.58%~6.15%,染色体长度比为1.74。染色体臂比范围为1.02~2.29, 平均臂比为1.35,臂比大于2 的比例为7.14%。染色体不对称系数为57.52%,核型为2A 型。
E.cylindricus:核型公式为 2n=6x=42=36m+6sm。42 条染色体中6 条染色体为近中部着丝粒染色体,36 条中部着丝粒染色体,无随体染色体。染色体相对长度为3.76%~5.85%,染色体长度比为1.70。染色体臂比范围为1.01~2.27, 平均臂比为1.29,臂比大于2 的比例为4.76%。染色体不对称系数为56.31%,核型为2A 型。

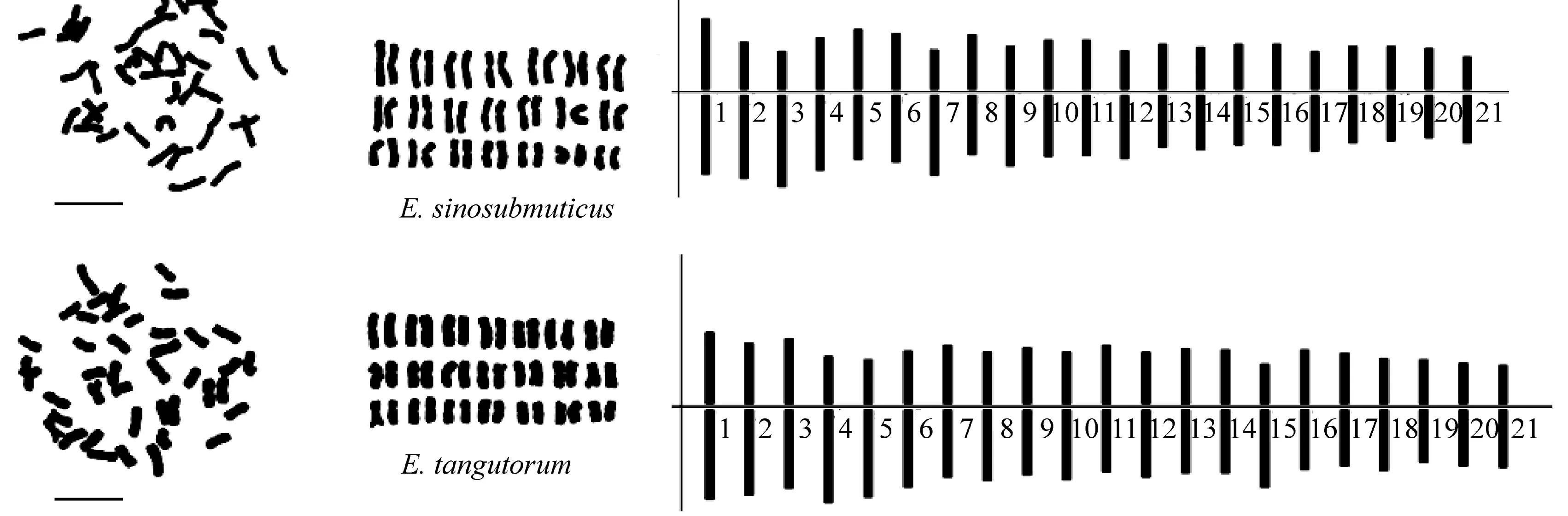
图1 六倍体披碱草属物种的染色体形态、核型图和核型模式图Figure 1 Mitotic metaphase chromosomes, karyograms and ideograms of the hexaploid Elymus species.scale bar=10 μm.

表2 披碱草属物种核型参数Table 2 Parameters of karyotype of Elymus species
E.dahuricus:核型公式为 2n=6x=42=36m+6sm。42 条染色体中6 条染色体为近中部着丝粒染色体,36 条中部着丝粒染色体,无随体染色体。染色体相对长度为3.91%~5.80%,染色体长度比为1.62。染色体臂比范围为1.05~1.93, 平均臂比为1.30,臂比大于2 的比例为2.38%。染色体不对称系数为56.61%,核型为2A 型。
E.excelsus:核型公式为 2n=6x=42=34m+8sm。42 条染色体中8 条染色体为近中部着丝粒染色体,34 条中部着丝粒染色体,无随体染色体。染色体相对长度为3.94%~5.78%,染色体长度比为1.63。染色体臂比范围为1.02~2.35,平均臂比为1.36,臂比大于2 的比例为11.9%。染色体不对称系数为57.67%,核型为2A 型。
E.nutans:核型公式为 2n=6x=42=38m+4sm。42条染色体中4 条染色体为近中部着丝粒染色体,38条中部着丝粒染色体,无随体染色体。染色体相对长度为3.46%~5.79%,染色体长度比为1.78。染色体臂比范围为1.03~2.86,平均臂比为1.37,臂比大于 2 的比例为 7.14%。染色体不对称系数为57.82%,核型为2A 型。
E.purpuraristatus:核型公式为2n=6x=42=36m+6sm。42 条染色体中6 条染色体为近中部着丝粒染色体,36 条中部着丝粒染色体,无随体染色体。染色体相对长度为3.49%~6.18%,染色体长度比为1.87。染色体臂比范围为1.03~2.35, 平均臂比为1.32,臂比大于2 的比例为7.14%。染色体不对称系数为56.97%,核型为2A 型。
E.sinosubmuticus:核型公式为2n=6x=42=36m+6sm。42 条染色体中6 条染色体为近中部着丝粒染色体,36 条中部着丝粒染色体,无随体染色体。染色体相对长度为3.48%~6.44%,染色体长度比为2.03。染色体臂比范围为1.05~2.39, 平均臂比为1.33,臂比大于2 的比例为7.14%。染色体不对称系数为57.17%,核型为2B 型。
E.tangutorum:核型公式为2n=6x=42=36m+6sm。42 条染色体中6 条染色体为近中部着丝粒染色体,36 条中部着丝粒染色体,无随体染色体。染色体相对长度为3.78%~6.26%,染色体长度比为1.75。染色体臂比范围为1.06~1.95,平均臂比为1.33,臂比大于2的比例为7.14%。染色体不对称系数为57.02%,核型为2A 型。
3 讨论与结论
刘玉红[6]于 1985年报道 E.atratus、E.breviaristatus、E.cylindricus、E.dahuricus、E.nutans 和 E.tangutorum 的核型类型为1A 型,在本文中以上物种的核型类型均为2A 型,其报道E.excelsus、E.purpuraristatus 的核型类型为2A 型,与本文结果一致。蔡联炳和冯海生等[7]于1997年报道E.breviaristatus和E.sinosubmuticus 的核型为 2A 型,E.breviaristatus的核型类型与本文一致, 本文发现E.sinosubmuticus 的核型为2B 型, 其报道的染色体相对长度、随体染色体条数等染色体参数与本文报道结果不一致,我们认为造成上述差异的原因可能有①材料居群间的核型分化。六倍体披碱草属物种分布广泛,所选择材料的来源不一样,染色体核型可能存在分化,从而表现出差异性;②染色体核型的研究主要基于染色体长度的测量, 人工测量采用的方法,以及测量的角度均不同,都将对结果造成误差;③随体作为染色体的一个重要特征,可能受到制片过程的影响,制片过程中预处理时间太长会造成染色体太短,随体不容易分辨,另外传统压片法以及最新的酶解滴片法对染色体的物理结构影响也不同。
本研究的六倍体披碱草属物种的染色体主要以中部着丝粒染色体为主,有少量亚中部着丝粒染色体。各个物种间的核型参数(染色体相对长度,染色体长度比,染色体臂比,平均臂比,臂比大于2 的比例)等均存在差异。但是从核型公式来看,E.dahuricus 复合群内的物种 E.cylindricus,E.dahuricus,E.purpuraristatus、E.tangutorum 的核型一致,均为 2n=6x=42=36m+6sm, 复合群内另一个物种E.excelsus的核型不同,为2n=6x=42=34m+8sm,但是复合群内的物种核型类型均为2A 型, 说明亲缘关系较近的复合群内物种核型特征也具有相似性。T.Oinuma[12]认为物种核型和物种染色体组存在平行进化关系,前人研究表明复合群内物种E.dahuricus 染色体组组成为StYH[13],可能复合群内其他物种也具有相同的染色体组组成。E.breviaristatus 和E.sinosubmuticus 的核型存在差异,分别为2A 和2B,究竟二者亲缘关系如何还有待进一步研究。
猜你喜欢
美文(2022年18期)2022-10-20
生物技术进展(2022年1期)2022-02-11
临床检验杂志(2021年10期)2021-11-24
川北医学院学报(2021年6期)2021-07-13
西藏农业科技(2021年4期)2021-04-18
中国科学院大学学报(2021年2期)2021-03-18
三农资讯半月报(2020年16期)2020-09-14
植物研究(2019年6期)2019-11-15
科学之谜(2018年9期)2018-12-17
中小学教学研究(2018年4期)2018-06-15
