
干旱区荒漠植物体内潜在水源差异及利用策略分析
2019-10-10 01:18李文静吕光辉张磊王恒方李周康王金龙马辉英刘志东
生态环境学报 2019年8期
李文静 ,吕光辉 *,张磊 ,王恒方 ,李周康 , 王金龙 ,马辉英 ,刘志东
1. 新疆大学资源与环境科学学院,新疆 乌鲁木齐 830046;2. 绿洲生态教育部重点实验室,新疆 乌鲁木齐 830046
在干旱区生境下,水作为植物生长发育过程中不可或缺的物质基础,是荒漠植物生长、繁殖等阶段的关键限制因子(Dai et al.,2015;付青云等,2019),因而,生态水文过程对土壤-植被体系的功能及演化方向起着决定性作用。研究发现,植物体所能利用的水源主要来自自然环境中的土壤水、降水、地下水以及地表径流(尹力等,2012)。植物体内各水源的比例及其在水文生态过程中的作用迥异,且随环境异质性发生变化。在水资源限制的背景下,研究植物对不同水资源的利用及分配规律的差异(李晖等,2006),有利于揭示干旱环境条件下植物共存机理及其对水资源的适应机制。
采用传统水文学方法很难研究植物体内水分来源、吸收利用过程(余绍文等,2011)。近年来,稳定性同位素技术由于具有较高的精确性和灵敏性,已成为示踪植物水分来源的有力工具(Cook et al.,2006;Vrzel et al.,2018)。关于植物对水源利用的研究,国内外学者采用氢氧稳定同位素示踪技术,将植物按照生活型及根系类型的差异分别展开相关分析。在不同生活型方面,Dodd et al.(1998)研究发现在植物群落中,乔灌草植物的水分利用方式存在差异,灌木和草本以地表降水为基础水源,而高大乔木所用基础水源为地下水。Villagra et al.(2011)研究了沙漠植物中典型的乔木、灌木及多年生的草本植物对水源的吸收利用策略,表明乔、灌木以深层土壤水作为第一水源,但草本植物依赖降雨维持其生存。以往的研究结果表明,不同生活型植物对各水源的利用存在一定差异,但乔木与高大灌木一般将稳定地下水作为首要水分来源,草本植物主要利用表层土壤水(Wu et al.,2016);在植物的不同根系分布范围方面,Hoekstra et al.(2014)发现与单一植被结构类型相比,浅根系与深根系物种的共存存在着垂直生态位互补现象。付青云等(2019)对3种林龄小叶锦鸡儿(Caragana microphylla)水分利用关系的探讨中,发现根系的分布决定着各个林龄小叶锦鸡儿对水分的利用范围及选择合适的用水策略。综上所述,利用稳定同位素技术对植物水分来源的研究有较全面认识(张光辉等,2006;林光辉,2013)。但对于干旱区内不同生活型植物对所需水源的吸收利用效率方面鲜见报道(刘保清等,2017),且以往研究未将植物以生活型、根系类型综合探讨它们对水分来源的影响。此外,以往研究大都关注于植物常见所需水源,而本研究位于干旱荒漠区,故考虑将凝结水作为植物体内所需潜在水源之一,进一步探析凝结水对植物生态水文过程的影响。
本文以艾比湖流域干旱荒漠区常见的12种植物为研究对象,通过分析其木质部不同潜在水源的δ18O值,再利用多源线性混合模型估算植物体内水分来源的比例关系,旨在回答如下3个科学问题,(1)植物因生活型差异对各潜在水源的利用效率是否存在差别?(2)不同根系类型植物之间是否存在水分利用的生态位垂直互补现象?以此揭示植物的生存策略。(3)凝结水作为干旱区一种重要的水源,对荒漠植物有何影响?解决上述3个科学问题,能够初步揭示荒漠植物对干旱环境的适应机制,为干旱荒漠区植物用水策略及生态建设提供科学依据和理论支持。
1 材料与方法
1.1 研究区概况
新疆艾比湖湿地自然保护区(44°30′-45°09′N,82°36′-83°50′E)于精河县西北,地处准噶尔盆地的西南方,总面积达2670.85 km2。该区域是准噶尔盆地西南缘的最低洼地和水盐汇集中心,是准噶尔盆地最低点。处于内陆干旱区和阿拉山口大风通道区,气候干旱,降雨稀少,属典型温带大陆性干旱气候。极端最高气温44 ℃,最低气温-34 ℃,年平均气温5 ℃,年平均降水量105.17 mm,年平均蒸发量约1315 mm(张雪妮等,2019),在这种干旱少雨的气候条件下,降水在没有入渗到土壤之前就已经被强蒸发消耗殆尽(马辉英等,2017),所以本文所测的潜在水源没有考虑大气降水。受地形地貌特征及气候条件的影响,艾比湖流域的植被分布同时受中亚和蒙古两个植物区系影响的过渡特征明显,是新疆境内荒漠植物种类最多的区域,以干旱荒漠植物为主,其中典型的乔木:胡杨(Populus euphratica)、梭梭(Haloxylon ammodendron);灌木:柽柳(Tamarix ramosissima)、盐爪爪(Kalidium foliatum)、白刺(Nitraria tangutorum)、骆驼刺(Alhagi sparsifolia)、罗布麻(Apocynum venetum);草本:芦苇、碱蓬(Suaeda microphylla)、猪毛菜(Salsola sinkiangensis)等(杨晓东等,2009;郄亚栋等,2018)。地下水埋深约1.8-2 m。
1.2 样地布设与样品采样
本试验区选在国家级自然保护区艾比湖湿地东大桥管护站附近(图1),环境背景较为一致的一个3×3 km2区域,所涉及的植被类型是研究区典型的乔-灌-草植物群,并按根系类型将其分为中深根系与浅根系植物类型(马辉英等,2017)。于2016年7月中旬,北京时间06:00-07:00进行采样(为更好收集凝结水,减少强蒸发对实验造成误差),采样期间无降水,并在其中选取乔木:胡杨、梭梭;灌木:柽柳、盐爪爪、骆驼刺、白刺、罗布麻;草本:芦苇、盐生草、碱蓬、猪毛菜、灰绿藜(Chenopodium glaucumL.),每个物种均选取生长旺盛、长势良好的植株作为采样对象。每个物种采取3株形态特征相似的植株,其中灌木和乔木均选择超过两年的茎,去除茎段的外皮和韧皮部,保留具有导水能力的木质部,而草本则采集根与茎相连的部分,迅速将植物木质部装入25 mL同位素样品收集瓶,盖好瓶塞,用parafilm封口膜封好瓶盖,并立刻将样品冷藏于液氮罐中。
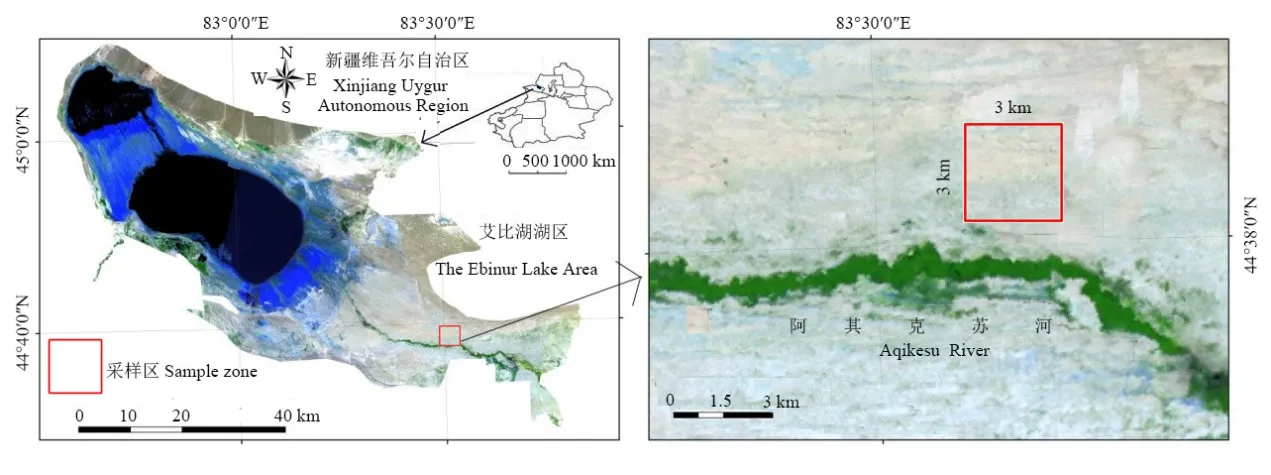
图1 研究区示意图 Fig. 1 The studied region
同时,利用对角线取样法采集土壤样本,各土壤样本分为(0-40、40-70、70-100、100-150 cm)共计4个土壤层。利用土钻取土用于土壤水的抽提和土壤含水量的测定,用铝盒将采集的各土层土壤样品封装并标记,用于后期土壤含水量的测定。采用研究点附近的3口井井水水位的深度代替地下水位;河水的收集样点设置为:研究区平行于阿其克苏河方向,分为3个点,间距为1 km,为了防止表层河水强蒸发而可能导致的分馏作用,故收集2.5 m处的深层河水。于植物冠层等高度随机放置3-5个培养皿用于收集冠层凝结水(当地时间21:00-22:00),每个培养皿中放置3-5片干布片,翌日(当地时间06:00-07:00)将润湿布片塞入25 mL同位素样品收集瓶。将河水和地下水的样品,装入25 mL同位素样品收集瓶,盖好瓶盖,并用parafilm封口膜封好瓶盖,放入便携式车载冰箱(-5 ℃)中保存,直至样品测定。
1.3 样品水分提取与分析
采用低温真空蒸馏法抽提植物可能吸收的潜在水源和木质部水分,随后将抽提出的液态水使用LGR液态水同位素分析仪(LWIA-V2 (DLT-100),LGR,American)测定稳定氢和氧同位素值:
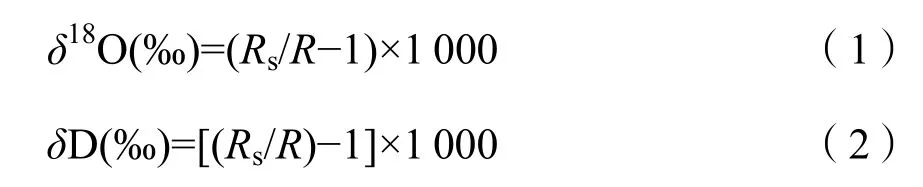
式中:δD和δ18O分别为对应样品的氢和氧同位素值;Rs和R分别为样品国际通用标准物中元素的重轻同位素丰度之比(如18O/16O);精确度(1δ)18O/16O优于0.1‰,D/H优于0.3‰;δD值的测试误差不超过1‰,δ18O值的测试误差不超过0.2‰。
1.4 模型选择
在利用稳定同位素技术测定植物水分来源时根据实际情况选择不同的方法,本文选择多源线性混合模型IsoSource法。其基本原理为:如果植物体内水分的来源方式有多种情况,则表征植物体内水的同位素值是各个潜在水源的同位素值的线性混合值,因此,线性混合模型如下:
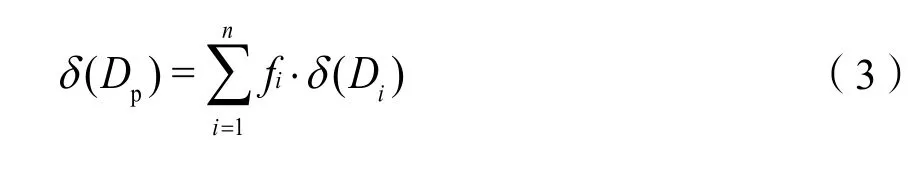

上式,δ(Dp)、δ(18Op)表示植物体内的水δD和δ18O值;而δ(Di)、δ(18Oi)分别是水源i的δD和δ18O值;fi是植物对水源i的吸收比例。潜在水源个数小于3的情况下,同时利用D和18O同位素,可求出fi的唯一解。在水源个数大于3时,Phillips et al.(2005)将确定各潜在水源贡献比率上、下限的IsoSource方法(增量定为1%,允许偏差定为0.01)。目前,相关研究表明,不论在单个生长季节内或是在年际的尺度上,植物体内液态水同位素组成的差异均能反映其吸收水源同位素的组成变化(段德玉等,2007)。
1.5 数据处理
采用IsoSource软件分析各个潜在水源对植物的贡献比例,不同深度土壤水分的δ18O值及不同生活型植物对水分利用率差异采用SPSS 17.0统计软件,用Origin 8.0作图。
2 结果分析
2.1 土壤含水量及δ18O在不同土层间的变化
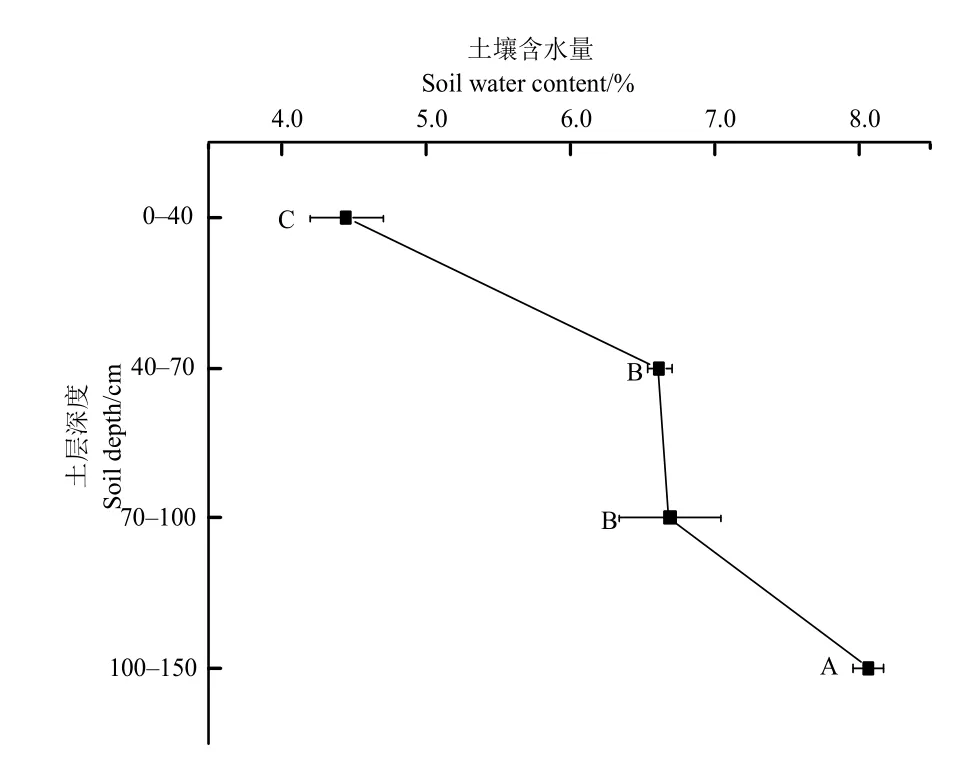
图2 不同土层的土壤含水量 Fig. 2 The soil water contents of the different soil layers
研究区土壤含水量随土壤垂直结构的变化趋势如图2所示。其中土壤含水量最大和最小值分别在土层100-150 cm和0-40 cm处,分别为(8.07%± 0.10%)、(4.45%±0.26%);土层40-70 cm和70-100 cm的土壤含水量大小分别为(6.62%±0.08%)、(6.69%±0.35%),二者没有显著性差异(P>0.05)。以上结果表明研究区土壤含水量随着土层深度的增加表现为逐渐增加趋势。
随土壤深度的递增,土壤水的δ18O值显著偏负,δ18O值在0-40 cm(-3.995‰±0.09‰)和40-70 cm(-5.756‰±0.07‰)表现出显著差异,与不同土层含水量表现出一致性差异,土壤水δ18O的变化范围为(-6.292‰--3.995‰)(图3)。
2.2 研究区常见的12种植物对各潜在水源的利用比例
研究区12种常见植物对各潜在水源利用率分析结果见表1,各潜在水源对植物的贡献率表现出明显的规律性,胡杨、梭梭、柽柳、盐爪爪、白刺、骆驼刺、罗布麻和芦苇均表现出将地下水作为最主要的水源(均大于58.0%),以各层土壤水及河水为辅,且较少利用冠层凝结水,其中柽柳对地下水的利用率最大为96.0%;而碱蓬、猪毛菜、灰绿藜表现出与之不同的水分利用策略,以土壤水作为第一大水源,特别是以表层土壤水为主,其中灰绿藜对表层土壤水利用率最大为91.0%,且这3种植物将冠层凝结水作为其第二大水源,对河水及地下水的利用率相对较低;盐生草则将地下水和各层土壤水作为主要水源,同时对河水、冠层凝结水的利用较均衡。
研究区优势种植物的δ18O值如表1所示,在物种水平上,胡杨、梭梭、柽柳的δ18O值较小,分别为-13.517‰、-13.302‰和-13.541‰,而碱蓬、猪毛菜、灰绿藜的δ18O值较大,为-4.351‰、-5.249‰、-4.296‰。
2.3 不同生活型植物体对各潜在水源的利用比例
按不同生活型将研究区植物划分为乔木、灌木和草本,而乔木、灌木在δ18O值之间未能表现出显著差异(P>0.05,图4),灌木与乔木植物仅对河水和100-150 cm的深层土壤水的利用存在显著差异(P<0.05),且地下水对乔木的贡献率均值为87%,表明乔木的生长主要依赖于稳定地下水资源,对水源利用较为单一,而灌木对河水和土壤水的利用相较于乔木偏高,表现出多面下注的策略。5种草本植物δ18O值变化范围为(-12.293‰--3.404‰),与乔灌木之间表现出显著差异(P<0.05),并将表层土壤水作为第一水分来源,地下水作为第二水源,总体表现为草本对地下水的平均利用率较大,这可能是由于中深根系植物芦苇、盐生草对地下水的利用率较高。中深根系的草本植物与灌木表现出相似的水分利用策略。
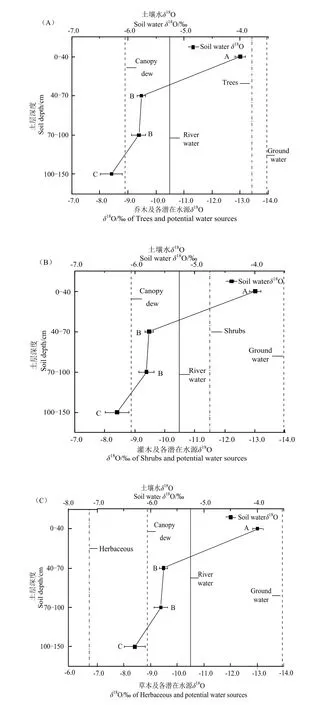
图3 δ18O在不同生活型植物和各潜在水源之间的变化 Fig. 3 δ18O the various of between the different vegetation types and the submerged water
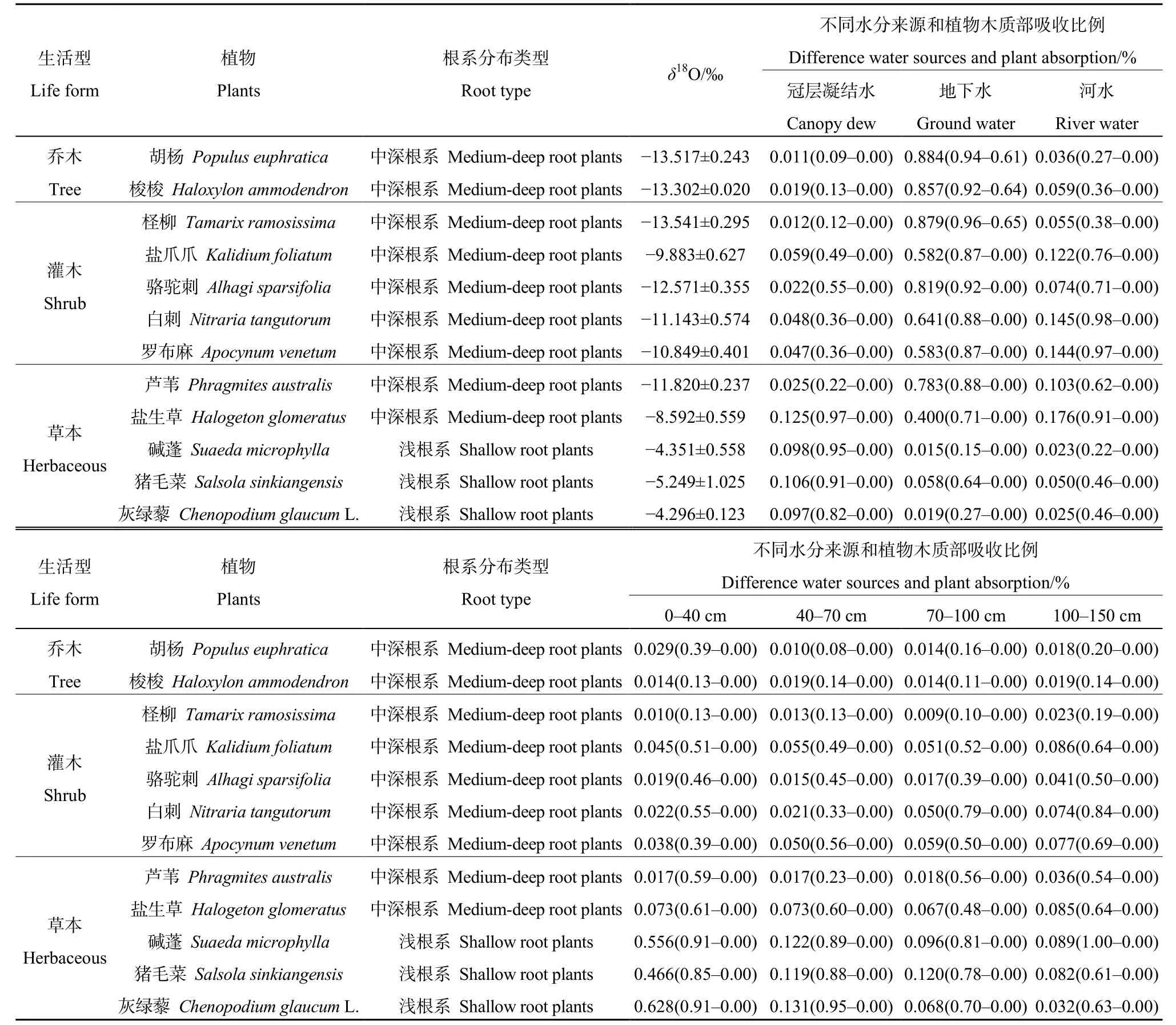
表1 研究区12种常见植物茎的氧同位素值及潜在水源贡献率 Table 1 The proportion of water sources and δ18O in 12 dominant families of the research area
对5种灌木,柽柳、骆驼刺、罗布麻、白刺和盐爪爪δ18O的均值为(-11.598‰±0.390‰),其值相对于冠层凝结水和河水偏负,而较为接近地下水-13.999‰(图5B)。两种乔木梭梭和胡杨对各潜在水源的利用,总体均值(-13.409‰±0.119‰)与灌木表现出类似的结果,对地下水-13.999‰更为接近(图5A)。
2.4 不同根系类型植物体对各潜在水源的利用比例
根据研究区植物根系的深浅将植物分为两类,中深根系与浅根系(图5)。除深层土壤水外,中深根系与浅根系植物在对其它各水源利用方面均表现出极显著差异(P<0.01)。特别是不同根系类型对地下水和表层土壤水的利用差异最大。中深根系植物对地下水的利用比例较高(均值为71.42%),而浅根系植物则将表层土壤水作为首要水源(均值为55.00%),同时将冠层凝结水作为第二大水源,由此可知,在中深根系与浅根系植物采用了空间垂直生态位互补策略,高效利用有限水资源。
由图6可知,中深根系植物的δ18O均值-11.691‰明显偏负于浅根系植物的δ18O均值-4.632‰。土壤水δ18O的变化范围为(-6.292‰--3.995‰),较为接近浅根系植物δ18O均值(-4.632‰±0.372‰),总体相对于冠层凝结水-8.890‰偏正,说明浅根系植物主要将土壤水和冠层凝结水作为主要水源,河水和地下水作为辅助水源(图6),而中深根系植物的δ18O均值-11.691‰较为接近地下水与河水的δ18O均值,表明该类型植物主要利用稳定的地下水及河水。中深根系植物胡杨、梭梭、柽柳的δ18O值最小,分别为-13.517‰、-13.302‰、-13.541‰,而浅根系碱蓬、猪毛菜、灰绿藜的δ18O值最大,为-4.351‰、-5.249‰、-4.296‰,由此发现根系越深δ18O值越偏负,根系越浅δ18O值则偏正,根系与δ18O值可能存在负相关。
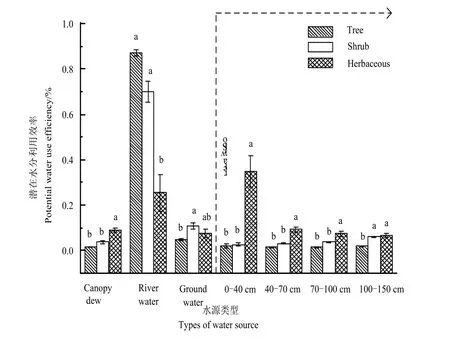
图4 不同生活型植物对各潜在水源利用率 Fig. 4 The utilization rate of different life types of plants to each potential water source
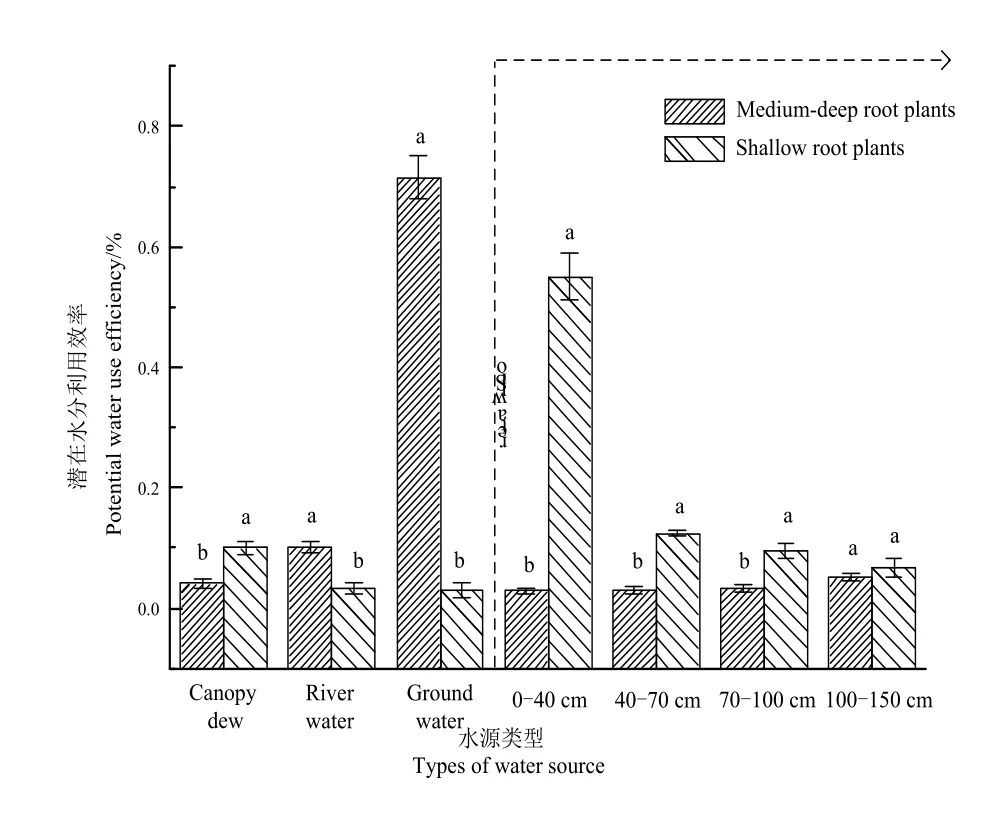
图5 不同根系类型植物对各潜在水源利用率 Fig. 5 The utilization rate of different root types of plants to each potential water source
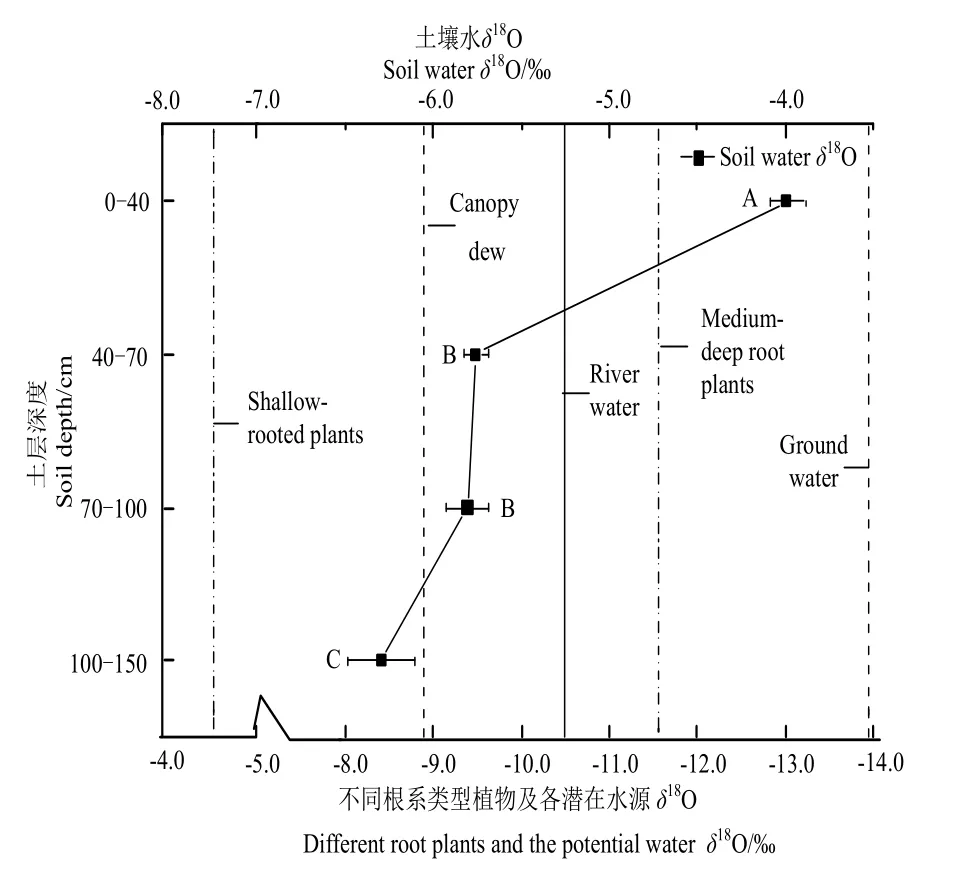
图6 δ18O在不同根系类型植物和各潜在水源之间的变化 Fig. 6 δ18O the various of between the different root types and the submerged water
3 讨论
本文中植被δ18O的变化范围在(-13.993‰--3.404‰)之间,这与以往对准噶尔盆地干旱荒漠植物体内的δ18O(-11.930‰--3.980‰)的研究结果较为接近(周海等,2013),表明干旱区荒漠植物体内δ18O的变化范围大致相同。研究区土壤水δ18O的显著变化层分别为40-70 cm和100-150 cm(P<0.05),这可能是与该层的土壤含水量有关,干旱区的土壤水分含水量,表现为表层最低,深层最高,因为表层强烈的蒸发,加上浅根系植物的利用,而深层则因为地下水补给,或者植物根系利用的偏少,以及渗漏补给等,蒸发也比较弱。本研究区地下水较浅,很容易通过毛细管作用补给深层土壤水(100-150 cm),使其相较于其它的土层深度,表现出更显著的偏负现象。越接近地表的土层深度,土壤水δ18O值逐渐偏正,土层深度从40-70 cm到0-40 cm,土壤水δ18O值发生了显著的偏正现象,说明在极端干旱的环境条件下,由于干旱区的强蒸发使得表层土壤水的氧同位素分馏速率加剧,最终导致氧的轻同位素16O显著蒸发,从而引起18O的富集,Asbjornsen et al.(2007)的研究也发现土壤水δ18O随着土层深度的加深而逐渐偏负。赵良菊等(2008)在关于土层深度对河岸林植物的研究中指出120 cm以上的土层发现了δ18O明显的偏正现象,这可能是由于该地区为受大陆性季风气候影响,土壤蒸发强度高所致。
艾比湖流域属于温带荒漠生态系统,其植被类型主要以沿河岸的杜加依林及旱生植物群为主。乔木、灌木木质部δ18O的均值分别为-13.409‰、-11.598‰,它们与地下水δ18O值-13.999‰更为接近。且乔木与灌木在水分利用方面,除灌木对河水利用显著大于乔木,剩余水源类别未能表现出显著差异。在对植物水源利用的前期研究中,Dawson et al.(1991)发现河岸植被并不直接利用河水,而是使用更为稳定的地下水,随着地表的干燥程度的增加,它们对地下水利用比例也上升,而在艾比湖荒漠林中,发现河水在乔灌中均有所贡献,且灌木对其利用率达10.8%,造成这种差异可能是因为研究区荒漠林地表较为干燥,乔木对水分的利用方式主要集中于地下水资源,灌木除较多利用地下水外,也多元利用其它水源;造成这种现象的原因可能为更好利用水资源,乔木对生长环境和水分利用效率上采取权衡策略,且乔木生活史较长,趋利避害,采取不断向下扎根的生存策略,而灌木与乔木相比,在对地下水的竞争方面处于劣势,为此,只能采用多面下注的生存策略,以此减少与乔木之间的生态位重叠,提高自身的生存、生长及繁殖适合度,通过协同进化作用与长期对环境的适应,从而达到其对生境中各潜在水源的综合利用能力。因此,地下水是乔灌木植物生长的第一水分驱动力。在干旱荒漠区,高大乔灌木的生长、繁殖及种群更替是由地下水作为主要驱动因子,荒漠区植物生长对地下水有很强的依赖性(樊自立等,2004;张江等,2018)。可知,地下水埋深是影响植物群落水分利用策略的主要因素。本研究中选取的乔木和灌木,对地下水的利用率平均达78.55%。这可能是由于阿其克苏河系的渗透和补给作用,使得距离河湖区较近的区域,地下水埋深较浅,土壤中盐分含量较高,在该区域中土壤盐分含量随着土壤剖面的增加而减小(张雪妮等,2019),植被为了减少盐渍化带来的胁迫,更多的吸收盐分含量相对较少的地下水资源,而很少吸取利用含盐量较高的表层土壤水。同时,乔灌木的光合产物对地下部的投入明显大于草本植物,而且灌木的对吸水层的选择性高于草本,对水分的利用可塑性更大。特别是受到水分胁迫时,乔灌木能够由土壤的水分利用性的变化而随之变化(Cipriotti et al.,2014;李巧燕等,2019),因而,在干旱、半干旱荒漠区,乔灌木较草本植物而言,对区域内水源更具有竞争优势。
另一控制植物的水分来源的关键因子是植物根系分布及根系活性。各植物之间的根系分布格局异质,致使植物的吸水模式和水分来源存在差异(Nie et al.,2012;陈小丽等,2014)。本研究发现,在艾比湖流域不同生活型荒漠植物的水源利用效率随土壤深度表现为:草本<灌木<乔木;草本植物的根系类型表现为:浅根系草本<中深根系草本,乔灌木的吸水范围大于草本,这也表明了建群种的生态适应较强,占有较宽的生态位。有研究表明(祁亚淑等,2015;Xu et al.,2017),中深根系的植物具有水力提升作用,可将深处的地下水、土壤水分运移到表层干燥土壤,以此为浅层植物的根系提供水分。根系是获取水分、养分的主要器官,同时还能感知逆境胁迫。根系能对土壤中分布不均的水资源进行再分配,它将水分含量多的区域吸收运送到亟需用水的部分,中深根系植物这种对水分的再分配,为伴生的浅根系植物提供新的水源。在地下水作用下,通过调整根系分布,促进浅层土壤水分的吸收,从而优化根系结构,对水分有效性的变化表现出较好的适应性。Hoekstra et al.(2014)发现与单一植被结构类型相比,浅根系与深根系物种存在着垂直生态位互补现象,对植物间的共存有着积极作用。在本研究中,浅根系草本猪毛菜、碱蓬及灰绿藜,它们根系无法达到地下水位,但地下水仍然对这些植物生长有贡献,这也证明了不同生态位存在合作关系,且这种促进关系能有效弥补干旱区的水分匮乏问题,使植物群落间充分利用水源,进而提高了该生态系统的水资源利用的效率和在逆境环境中的稳定性。众所周知,研究内容如若牵涉地下部分,其难度将会远远大于地上。因此,植物对水源的认识深度和范围具有相对局限性。本文对植物根系分类仅分为中深根系和浅根系两类,建议今后的研究应予以细化。
依照形成界面,可将凝结水划分为土壤凝结水与冠层凝结水,土壤凝结水很少能被根系吸收利用,植物主要在冠层界面上通过叶片吸收凝结水(龚雪伟等,2016)。本研究中发现,灌木与乔木在冠层凝结水利用模式上有明显差异,灌木对冠层凝结水利用率比乔木高。另外,不同生活型植物对冠层凝结水的利用从大到小依次表现为:草本(9.0%)>灌木(3.8%)>乔木(1.5%),草本植物对冠层凝结水的利用最高。这可能是因为高大的乔灌木根系深更容易触及地下水,而草本植物将所获得的水分更多投资于地上部,要依靠增加地上部分性状,改变植物的地上部与地下部生物量分配(Xu et al.,2017)。在水资源短缺,环境恶劣,竞争激烈的荒漠干旱区,草本植物更多采取机会主义生活史策略,尽可能多的抓住获得水资源的机会(龚雪伟等,2016)。本文发现浅根系草本植物对凝结水依赖程度高于中深根系植物,并将其作为第二大水源。可见,凝结水对干旱荒漠区的草本,尤其是浅根系草本的生存提供所需的水分保障,适当缓解植物对干旱的胁迫状况(郭占荣等,2002;方静等,2015)。
综上可看出,在干旱区,长期干旱、地下水埋深较深或者土壤水分不稳定的情况下,猪毛菜、灰绿藜、碱蓬等浅根系草本植物需要与胡杨、梭梭、柽柳等中深根系植物共存,借助中深根系植物的水分供给满足自身生长对水分的需求。浅根系植物和中深根系植物是恢复艾比湖流域脆弱生态环境的良好组合方式。在干旱荒漠区土壤水和地下水是多数荒漠植被生存所依赖的最为重要的水分来源,是决定荒漠区植被分布、生长、种群演替以及荒漠绿洲存亡的主导因子。因此,在艾比湖流域维持地下水水位和表层土壤水对维护荒漠植物的正常生长极为重要。了解荒漠植物的水分利用机理及适应极端干旱环境的策略,为恢复与重建受损的荒漠流域生态系统和维护荒漠植被的稳定及可持续发展提供理论依据。
4 结论
(1)不同潜在水源由生活型、植物根系类型的差异而对植物贡献率不同。①在生活型方面,乔木对水分的利用主要集中于单一地下水资源;灌木除较多利用地下水外,也多元利用其它水源;而草本植物因根系深浅不同,植物水分来源存在差异。②中深根系植物与浅根系植物对水源的利用在空间上存在垂直生态位互补现象,对物种共存及互利共生有着促进作用。中深根系草本植物利用地下水较多,浅根系草本植物主要利用表层土壤水。因此,在艾比湖荒漠区植物体水分来源和植物的生活型、根系分布范围相关,其中根系类型影响更为重要。
(2)凝结水在不同生活型体内利用大小为草本>灌木>乔木,且浅根系草本将冠层凝结水作为第二大水源,因此,冠层凝结水对浅根系草本的生长起着关键的促进作用。
(3)在干旱区,中深根系植物为浅根系植物提供基础的水分需求,浅根系植物为中深根系植物固着表层土壤水,因此,浅根系植物和中深根系植物的共存是恢复干旱荒漠区脆弱生态环境的良好组合方式,可提高荒漠植物对水分的综合利用效率。
猜你喜欢
农业工程学报(2022年1期)2022-03-25
科技创新导报(2021年33期)2021-04-17
中华建设(2019年12期)2019-12-31
全球定位系统(2019年4期)2019-08-30
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
乡村科技(2018年8期)2018-02-13
中国核电(2017年2期)2017-08-11
诗潮(2017年2期)2017-03-16
资源节约与环保(2014年12期)2014-02-01
植物营养与肥料学报(2010年3期)2010-11-16
