
定向演化技术与蛋白质工程
——解读2018年诺贝尔化学奖
2019-10-31 08:35徐经纬
生物学通报 2019年1期
杨 冬 徐经纬
(北京师范大学生命科学学院 北京 100875)
2018年美国化学家Frances Arnold 获得了诺贝尔化学奖。其主要成就是在蛋白质的定向演化(directed evolution)上作出了开拓性的贡献。利用定向演化技术,科学家可提高酶的活性,改变酶的底物或产物范围,提高酶在特殊环境下的稳定性等,从而使酶在工业上有更广泛的应用潜力。不仅如此,定向演化实验还可以为研究蛋白质在自然环境下的演化历程提供数据,从而在进化论的理论研究上作出贡献。什么叫定向演化? 它又有什么用途?
蛋白质在生命活动的各个方面都发挥着重要作用,同时,其作为生物催化剂或新型大分子药物也具有重要的应用价值。因此,通过改造蛋白质分子以改进其生物活性在工业上具有重要意义。这一技术称为蛋白质工程。由于蛋白质的性质是由其氨基酸序列所决定的,因此,通过改变蛋白质的氨基酸序列,就可能改变其特性,从而设计出具有新型特征的蛋白质。然而,要成功改造一个蛋白质,需知道应在其氨基酸序列的哪些位点作出哪些改变。目前主要有2种策略。一种策略被称为理性设计(rational design),即利用已知的该蛋白质结构和功能的信息,进行定点突变,从而改变该蛋白质的特性,实现对其的改造。但是,在这一领域中,最大的困难是目前还不能精确地建立蛋白质结构和功能之间的关系[1]。因此,对于大部分体系而言,理性设计的效率并不高。另一个策略则是进行定向演化[1-2]。在这一思路下,研究者不需要针对蛋白质的工作机理作出假设,而是利用进化的原理,经过多轮筛选,从突变文库中选择出具有所需特性的蛋白质。
定向演化的整个过程可以总结如下(图1)。首先需要建立一个突变文库,然后表达出蛋白质,并针对蛋白质的特定功能对其进行筛选,从而获得符合要求的突变体进入下一轮。通常,这些突变体带有能表达出具有所需特性蛋白质的突变基因。之后,在所获得的突变体基础上进行进一步突变,构建新的突变文库,再进行下一轮筛选。这样,经过多轮选择或筛选的过程,就有可能得到设计者所期望的最终产物。本文将以Arnold 的工作为例,介绍这几年定向演化研究的具体成果。
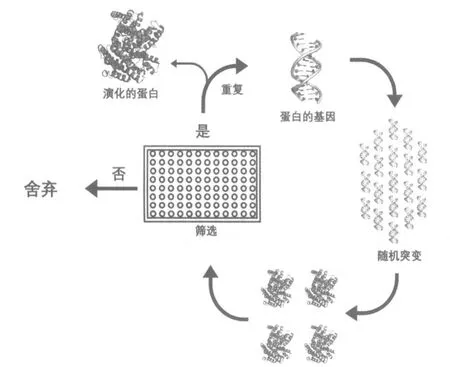
图1 定向演化的典型流程
1 改造枯草杆菌蛋白酶使之适应二甲基甲酰胺(DMF)环境
酶在有机溶剂环境下,有时可以催化其在水相环境中无法催化的反应。例如,丝氨酸蛋白酶在极性有机溶液环境下可以催化合成某些高分子化合物[3]。因此,通过改造酶的序列,增强其在某些有机溶剂中的催化能力,很可能具有工业上的应用价值。例如,可用于合成某些有经济价值的手性化合物。
Arnold 的研究组之前已获得了含有3位氨基酸替换的枯草杆菌蛋白酶突变体(3M)和含有4位氨基酸替换的突变体(4M),其在含有20%~40%的DMF 环境下较野生型具有更高的催化效率(即更高的kcat/Km值)(图2)[3]。在定向演化实验中,研究人员利用易错PCR 技术在4M 基础上引入了随机突变,构建突变文库并转化B.subtilis细胞,在含有酪蛋白和DMF 的固体培养基上进行培养。由于枯草杆菌蛋白酶是分泌蛋白,并可催化酪蛋白的水解反应,因此,其活性可通过观察培养基上的晕圈进行判断。通过挑选具有最大晕圈的克隆,即可获得本轮演化过程中的最佳突变体。然后对其再进行突变,构建突变文库,转化细胞进行下一轮筛选。经过几轮这样的筛选,最后就得到了能在高浓度DMF 环境下有效发挥催化作用的枯草杆菌蛋白酶突变体(PC3)[3]。
在40%的DMF 浓度下,PC3的催化效率是野生型的100倍以上,与4M 相比,也比其高2倍以上(图2)。不仅如此,PC3还可在更高的DMF 浓度下发挥催化作用。在60%和85%的DMF 条件下,其催化效率分别达到野生型的256倍和131倍。由于当初进行这一定向演化实验的主要目的是为了利用丝氨酸蛋白酶在极性有机溶剂条件下合成高分子化合物的能力,研究人员还特别对此进行了检测。结果发现,PC3在70%的DMF 条件下可催化合成约15mg 的多聚-L-甲硫氨酸,其聚合度达到4.7kD。而野生型蛋白质只能在60%的DMF 条件下合成聚合度约1.0~1.5kD 的多聚-L-甲硫氨酸[3]。

图2 枯草杆菌蛋白酶野生型蛋白质和各个突变体水解底物sAAPF-pna 的动力学参数(数据来自Chen,K[3])
2 逆转海因酶(hydantoinase)的对映体选择性(enantioselectivity)
海因酶的作用是水解5′-单取代海因(5-monostubstituted hydantoin),将其转变为N-氨甲酰基氨基酸(N-carbamoyl amino acid),而后者则可在L-氨甲酰水解酶(L-carbamoylase)的作用下转变成氨基酸。所以海因酶在工业界可被用于氨基酸的合成。但是,5′-单取代海因有L-和D-2种对映体(图3)[4]。在甲硫氨酸的合成过程中,海因酶偏好的底物是D-对映体,即D-5-(2-甲硫基乙基)-海因(D-MTEH),因此无法用于L-甲硫氨酸的合成。所以如果能将海因酶的底物选择性转变为L-对映体,即L-5-(2-甲硫基乙基)-海因(LMTEH),则将有重要的工业意义。
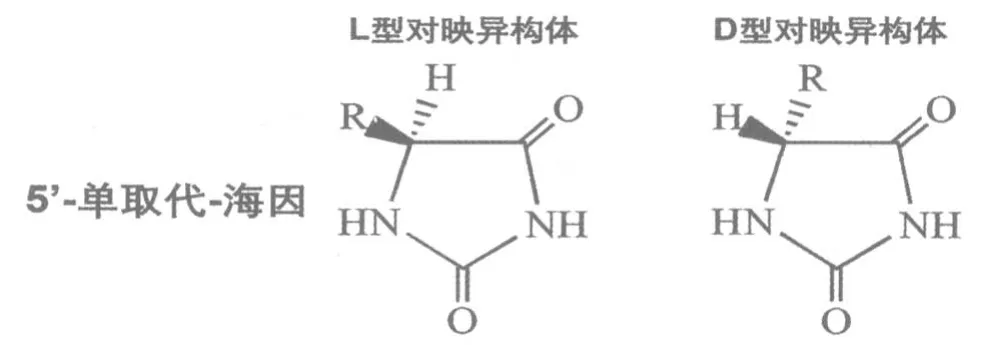
图3 5-单取代海因的L-、D-对映异构体[4]
在这项工作中,研究者通过易错PCR 技术在海因酶的序列中引入随机突变,构建了突变文库。之后将细胞培养在含有D-MTEH 或L-MTEH 的培养基上,并通过检测pH 的变化跟踪海因酶催化的反应,从而实现对突变体的筛选。通过筛选了约10000个克隆,研究者发现了突变体19AG11和11DH7。使用手性HPLC 技术对这2个突变酶进行研究表明,虽然它们还是偏向使用D-MTEH作为底物,但其对D-对映体的选择性显著变弱。由于这2个突变体都包含有I95L 突变,研究者进一步构建了含有I95L/Q251R 突变的突变体11DH7,并在此基础上引入了新的随机突变进行第2轮筛选,得到了22CG2。该突变体的对映体选择性与11DH7相同,但是其活性有所提高[4]。
由于继续使用易错PCR 技术似乎很难进一步提高酶对L-MTEH 的选择性,研究者打算另辟途径。他们在前期研究中发现I95可能在对映体的选择性上有重要作用,于是就在22CG2的基础上,对第95位氨基酸进行了饱和突变。结果发现,一个突变体Q2H4(含有I95F)具有明显的L-MTEH选择性,且其活性在22CG2的基础上还有进一步提高[4]。在E.coli中构建含有Q2H4的代谢通路后,经过发酵可明显观察到L-甲硫氨酸的积累[4]。这一结果显示了该突变体在氨基酸合成工业上的潜力。这项工作同时证明了先使用易错PCR 取样序列空间进行粗筛,找到关键氨基酸后,再利用饱和突变进行更加精细的筛选,从而实现酶的定向演化这一思路的可行性。
3 通过定向演化改造类胡萝卜素合成路线中的酶从而获得新型产物
萜类化合物是一大类具有极大多样性的天然产物,其构成单元为异戊二烯。在生物合成过程中,细胞利用不同的反应途径先合成异戊二烯焦磷酸(isoprenyl pyrophosphate,IPP)及其异构体二甲基丙烯焦磷酸(dimethylallyl pyrophosphate,DMAPP)。然后通过聚合反应,细胞可以合成包含有不同异戊二烯单元的线性分子,之后由不同的萜烯合成酶催化环化反应,再经过进一步的修饰(如羟化等)从而生成各类最终的萜类产物。类胡萝卜素属于四萜类化合物,即它们均含有8个异戊二烯单元。所有的类胡萝卜素的前体都是含有8个异戊二烯单元的八氢番茄红素(phytonene)。在该化合物的基础上,八氢番茄红素去饱和酶(phytonene desaturase)可以引入2-6个碳-碳双键(取决于该酶的物种来源),从而形成具有不同饱和程度的类胡萝卜素产物(图4)[5]。在通常情况下,八氢番茄红素去饱和酶引入的双键数量不超过5个。只有来源于E.uredovora的酶可以催化合成少量的完全共轭的3,4,3′,4′-脱氢番茄红素 (3,4,3′,4′-tetradehydrolycopene),即被引入6个双键的产物[5]。
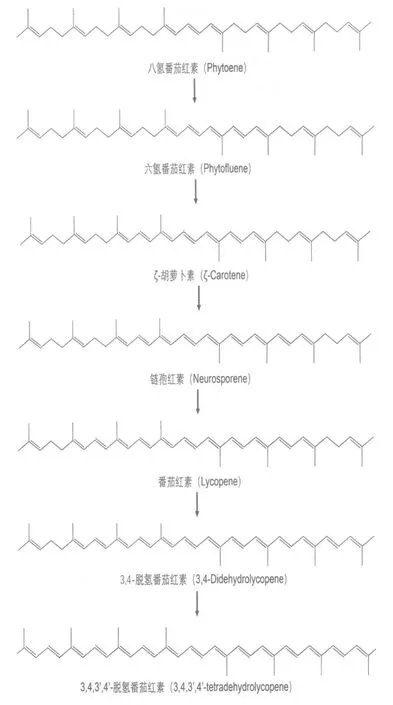
图4 八氢番茄红素去饱和酶催化从八氢番茄红素脱氢产生不同的类胡萝卜素产物
类胡萝卜素具有抗氧化活性,因此,其在抗肿瘤或慢性疾病的治疗方面具有潜在的应用价值。在这项研究中,Arnold 的团队试图探索通过改造类胡萝卜素合成通路上的酶,从而合成新型的类胡萝素的可能性[5]。在研究过程中,研究人员使用了DNA改组(DNA shuffling)技术,将来源于E.uredovora和E.herbicola的八氢番茄红素去饱和酶进行同源重组,从而获得了可供筛选的突变文库。之后,在E.coli中表达合成上游产物香叶基香叶基焦磷酸(geranylgeranyl pyrophosphate) 和八氢番茄红素的合成酶,以及这些八氢番茄红素的去饱和酶突变体。由于类胡萝卜素是有颜色的,因此可直接通过观察克隆的颜色进行筛选。通过筛选约10000个克隆,研究人员得到了突变体Ⅰ14[5]。Ⅰ14的颜色为粉色,说明其产物含有比番茄红素(lycopene)更多的双键。通过HPLC 最后鉴定该酶确实可催化3,4,3′,4′-脱氢番茄红素的合成。
在此基础上,研究者又同样使用DNA 改组技术,建立了番茄红素环化酶的突变文库。在表达香叶基香叶基焦磷酸和八氢番茄红素的合成酶,以及八氢番茄红素去饱和酶突变体Ⅰ14的E.coli细胞中,经过筛选4500个克隆,最后发现了一个鲜红色的克隆(Y2),其产物经过HPLC 分析被鉴定为包含有红酵母烯(torulene)[5]。该物质是3,4,3′,4′-脱氢番茄红素的前体3,4-脱氢番茄红素(3,4-didehydrolycopene)的环化产物。此项研究在历史上第1次鉴定出能通过直接环化3,4-脱氢番茄红素从而合成红酵母烯的酶[5]。
4 总结与展望
目前,通过定向演化技术对蛋白质分子进行改造可提高或改变其活性。这项技术具有很强的应用潜力,可以开发新型的酶用于化学合成工业,从而合成单纯使用化学合成方法难以经济性地实现其合成的产物。
此外,定向演化技术所获得的数据对于研究蛋白质的分子演化也有一定的价值。例如,Arnold组在研究细胞色素P450蛋白时,通过定向演化将原先催化长链脂肪酸羟化的P450BM3,改造成专一利用丙烷作为底物的P450PMOR2[6]。在这一定向演化实验中,研究人员进行了多轮筛选。在分析每一轮筛选所选中的突变体(即演化中间体)的催化性质后,发现随着演化的进行,其中间体对于丙烷的kcat逐渐增大,Km逐渐减小[7]。但是,蛋白质的热稳定性也随着演化的进程而逐步下降[7]。到突变体35E11时,蛋白质的稳定性达到最低值[7]。而之后的突变体ETS8的热稳定性则明显恢复,再之后的突变体19A12包含了一个重要的L188P 突变[7]。该突变本身对P450的稳定性有很大的削弱,但是此突变对于酶的专一性转向气相烷烃(比如丙烷)起着决定性的作用[7]。从19A12开始,该P450蛋白的最佳底物转变成了短链烷烃,其长度约3~4个碳原子,而之前的演化中间体则都有比较宽广的底物范围(包含3~9个碳原子的烷烃)[7]。
这一演化历程中,那些改变酶的底物特异性或提高其催化效率的突变在此演化体系中被认为是“有利”突变,是被选择的对象。然而,这些突变往往对蛋白质的稳定性具有破坏作用。因此,当蛋白质演化到某个阶段(即35E11)时,其稳定性已非常低,乃至不可能再积累这些“有利”突变。而同时另一些突变对酶的催化则没有直接影响,但是可提高酶的稳定性。在野生型酶或比较早期的演化中间体中,这些突变属于中性突变,因为此时酶的稳定性很高,再提高其稳定性并不会使之具有选择优势[2]。然而,当演化到ETS8的时候,因为具有增加蛋白质稳定性突变基因的突变体可容纳更多的“有利”突变,这将导致这些新一代的突变体能被选择出来。所以提高蛋白质稳定性的突变可提高蛋白质的可演化性(evolvability)[2]。在自然进化过程中,这些提高蛋白质稳定性的突变很可能是通过中性突变-随机漂变的方式在蛋白质中逐渐积累。因此,定向演化实验可能阐明在自然演化过程中中性突变对于蛋白质演化的影响作用。
猜你喜欢
——推动了照片保护与修复的研究
摄影之友(2022年11期)2022-11-08
井冈山大学学报(自然科学版)(2021年3期)2021-09-10
食品安全导刊(2021年20期)2021-08-30
中国食品学报(2020年7期)2020-08-03
日用化学工业(2019年1期)2019-01-30
中成药(2017年9期)2017-12-19
安徽农业科学(2015年13期)2015-12-18
医学研究杂志(2015年8期)2015-06-22
医药导报(2015年6期)2015-02-10
郑州大学学报(理学版)(2014年2期)2014-03-01
