
GA3对Cu胁迫下菠菜幼苗矿质元素吸收、细胞超微结构的调控效应及其耐Cu机理研究
2019-11-04 06:38公勤李兆华王玲宋雅琦康群戴同威
生态环境学报 2019年9期
公勤 ,李兆华 ,王玲,宋雅琦,康群,戴同威
1. 湖北大学资源环境学院,湖北 武汉 430062;2. 新疆农业职业技术学院,新疆 昌吉 831100;3. 湖北省农村安全饮水工程技术研究中心,湖北 武汉 430062
Cu是自然界广泛存在的一种金属元素。近年来,由于铜矿肆意开采和冶炼,农业中含Cu农药、含Cu粪肥,以及污水灌溉的不合理使用,导致土壤中 Cu 含量超标(Upadhyay et al.,2010;Możdżeń et al.,2016)。过量的Cu会抑制植物生长,降低光合作用,破坏质膜完整性,扰乱矿质元素吸收以及改变细胞超微结构(Zhang et al.,2010)。植物应对Cu毒害已经进化出多种机制,如将Cu与细胞壁结合、与植物螯合素螯合、液泡分隔等(Zhu et al.,2013),以提高植物对Cu的耐受性。
赤霉素(GA3)是一种重要的植物生长调节剂,除了具有促进细胞分裂、种子萌发、植物生长等作用,还能参与植物生理代谢、诱导信号传导(Lovegrove et al.,2000;Zhu et al.,2012),缓解重金属对植物毒害(Piotrowska-Niczyporuk et al.,2012),减少重金属在植物体内积累(Abd et al.,2009),提高植物的抗氧化能力(Alonso-Ramirez et al.,2009)。目前,GA3调节重金属对植物毒害的研究内容主要集中在重金属 Cd中,如 GA3缓解 Cd对龙葵Solanum nigrum(Ji et al.,2015)、银胶菊Parthenium hysterophorus(Hadi et al.,2014)、向日葵 Helianthus annuus(Ouzounidou et al.,2005)、万寿菊 Tageteserecta(Sun et al.,2013)和拟南芥Arabidopsis thaliana(Zhu et al.,2012)等植物的毒害作用,而GA3对其他重金属元素毒性的调节作用研究较少。
菠菜(Spinacia oleracea L.)是石竹目藜科的一种草本植物,具有叶面积大、相对生长率高、重金属吸收率高等特点,近年来,在科研中它与同科目的其他植物常用于观察其生长对重金属污染和毒性的响应研究(Naz et al.,2013;Alia et al.,2015)。Liu et al.(2005)、Khan et al.(2008)、Monu et al.(2008)的研究结果已表明菠菜具有一定的耐Cu性,本课题组前期的研究也得到了同样结果(Gong et al.,2019),但是该植物的耐Cu机理仍不清楚。因此,本研究以菠菜幼苗为试验材料,比较正常生长、Cu胁迫以及GA3与Cu复合胁迫作用下,菠菜幼苗生物量、氧化应激反应、渗透调节物质、Cu2+积累、矿质元素吸收以及细胞超微结构的变化,从生长生理角度揭示菠菜的耐 Cu机理以及 GA3对其耐 Cu机理的调节效应,为Cu污染地区蔬菜栽培管理和重金属耐性品种筛选提供试验依据。
1 材料培养与试验处理
1.1 试验材料培养
本试验供试品种为日本大叶菠菜,种子生产商为河北省青县王镇店种子繁育站。选取颗粒饱满的种子,用0.1% NaClO消毒10 min,再用去离子水冲洗干净后,均匀置于内有滤纸的培养皿(直径9 cm)中,放入25 ℃培养箱中避光催芽48 h后,在塑料大棚内将露白的种子播于泥炭土中。待幼苗长至10 cm左右时,挑选生长一致的幼苗移栽至长53 cm、宽23 cm、高18 cm的塑料盆内,每盆10株。盆下用托盘衬底,盆内基质为泥炭土、珍珠岩(7꞉3)混合而成,每盆基质总质量为3 kg(干重为2.1 kg)。培养20 d,待长出旺盛根系后进行试验处理,培养期间视土壤湿度情况浇灌完全 Hoagland营养液。
1.2 试验处理
试验设置 9个处理浓度,即 C1(Cu2+0 mg·kg-1+GA30 mg·L-1,作为对照),C2(Cu2+700 mg·kg-1+GA30 mg·L-1),T1(Cu2+700 mg·kg-1+GA33 mg·L-1),T2(Cu2+700 mg·kg-1+GA35 mg·L-1),T3(Cu2+700 mg·kg-1+GA310 mg·L-1),T4(Cu2+700 mg·kg-1+GA320 mg·L-1),T5(Cu2+700 mg·kg-1+GA340 mg·L-1),T6(Cu2+700 mg·kg-1+GA360 mg·L-1),T7(Cu2+700 mg·kg-1+GA380 mg·L-1),每个处理3个重复。Cu2+以CuSO4(分析纯)的形式提供。用完全 Hoagland营养液配制 700 mg·kg-1Cu2+溶液,用 0.1 mmol·L-1NaOH 或 HCl调节溶液pH值至(6.5±0.3)。对照(C1)只浇灌400 mL完全Hoagland营养液,C2-T7处理每盆浇灌400 mLCu2+溶液,溢到托盘内的溶液再次倒入盆内,以确保盆内土壤Cu2+终浓度为700 mg·kg-1。
第二天早上,除对照(C1)外,其余各处理幼苗叶部喷施对应浓度的GA3溶液。用1 mL酒精溶解各处理中的 GA3试剂,用含有 0.1% Tween 20(C58H114O26,一种表面活性剂和分散剂)的去离子水稀释GA3溶液至各处理浓度。用喷施面积为1 cm3的雾状喷雾器向叶片正面喷3次处理液,至叶片有水珠滴下,每盆总喷施量为20 mL;C2处理只喷施20 mL含有0.1% Tween 20的去离子水。处理期间,根据温度的变化,每天以去离子水补充土壤所蒸发损失的水分,使其盆土总质量保持在(3.4±0.05) kg。试验幼苗的培养条件为温度22/15 ℃(白天/夜晚),相对湿度70%-80%,光照14 h、黑暗10 h,光照强度为8000 lx。处理7 d后取样测定生理指标。试验材料和试验所用处理液浓度由前期大量预试验所得。
2 测定指标及方法
2.1 生物量测定
将各处理组幼苗从盆中小心取出,先用自来水冲洗,再用去离子水反复冲洗3次,吸干水分后,将叶、茎、根分开,迅速称鲜质量(FM)。随后,将鲜样装入烘干袋中,放入烘箱于105 ℃下杀青10 min后,再降至80 ℃烘至质量恒定,称干质量(DM)。
2.3 脯氨酸、可溶性蛋白含量测定
采用酸性茚三酮显色法测定脯氨酸含量(Tewari et al.,2002)。称取叶部鲜样,加入磺基水杨酸,沸水中提取冷却后,吸取上清液,加入冰醋酸和酸性茚三酮,于沸水浴中加热 30分钟后冷却至室温,加入甲苯,充分振荡,静置分层后取上层溶液,测定520 nm波长下的吸光度值。以牛血清白蛋白为蛋白标准,采用考马斯亮蓝G-250方法测定酶提物的总可溶性蛋白含量。将上清液和染料置于分光光度计试管中,在 595 nm处测定吸光度(Banu Doğanlar,2015)。
2.4 矿质元素含量测定
将各处理盆内的菠菜幼苗取出,剪下根,在20 mmol·L-1Na2-EDTA溶液中浸泡 3 h除去表面吸附的 Cu2+,用去离子水反复冲洗干净后用吸水纸吸干,分别装入烘干袋中;将菠菜叶片用去离子水反复冲洗干净后用吸水纸吸干,装入烘干袋中。将叶片、根样品放入烘箱中 105 ℃杀青后,80 ℃烘干至恒量,研磨粉碎后过80目筛。采用硫酸-双氧水消解,半微量凯式定氮法测定植物N含量;采用硝酸-双氧水消解,ICP-AES测定植物中的P、K、Cu、Ca、Mg、Fe 含量(Dos Santos et al.,2014)。
2.5 透射电镜观测
随机选取各处理盆中幼嫩的菠菜叶片,每个处理选取3株幼苗,将其幼嫩叶片从中部切成连续的3段,每段约2 mm,迅速放入装有2.5%戊二醛(pH 7.4,0.1 mol·L-1的PBS缓冲液配制)的带塞瓶内,抽真空后于4 ℃下固定4 h,再转入1%锇酸(pH 7.4,0.1 mol·L-1的 PBS缓冲液配制)中于室温(20 ℃)下固定2 h,经乙醇系列脱水,SPURR树脂浸透、包埋、聚合,在 LKB型超薄切片机上切片,经醋酸铀-柠檬酸铅双重染色后,用透射电子显微镜(JEOL TEM-1230EX,Tokyo,Japan)观察并拍照(Gill et al.,2015)。
2.6 数据分析
运用Excel 2007和SPSS 17.0软件对数据进行统计分析。采用单因素(One-way ANOVA)和Duncan法进行方差分析和多重比较(α=0.05),用Pearson法对各指标进行相关分析。运用Origin Pro 9.0软件作图。图表中数据为平均值±标准差。
3 结果与分析
3.1 Cu胁迫及GA3对Cu胁迫下菠菜幼苗生物量的影响
图 1显示,与对照(C1)相比,Cu胁迫时菠菜幼苗总FW、总DW均为显著降低趋势(P<0.05),较对照(C1)相比分别降低了 31.9%、40.9%。喷施GA3后(T1-T7处理),与C2处理相比,菠菜总FW、总DW均呈增加趋势,其中T1、T2处理幼苗总FW、总DW的增幅最大,总FW较C2处理增加了25.5%和19.3%,总DW较C2处理增加了37.4%和26.7%。随GA3喷施浓度增加,幼苗总FW、总DW增幅逐渐减少,T6、T7处理幼苗的总FW、总 DW均降至最小值。此外,各处理下菠菜幼苗叶、茎、根部FW和DW的变化规律与总FW、总DW的变化规律一致。
3.2 Cu胁迫及GA3对Cu胁迫下菠菜幼苗MDA、ROS含量的影响

图1 Cu胁迫及GA3对Cu胁迫下菠菜幼苗鲜质量(a)、干质量(b)的影响Fig. 1 Effect of Cu stress and GA3 on fresh mass (a) and dry mass (b) of Spinacia oleracea seedlings under Cu stress

图2 Cu胁迫及GA3对Cu胁迫下菠菜幼苗MDA(a)、(b)、H2O2(c)含量的影响Fig. 2 Effects of Cu stress and GA3 on contents of MDA (a), (b) and H2O2 (c) in Spinacia oleracea seedlings under Cu stress
3.3 Cu胁迫及GA3对Cu胁迫下菠菜幼苗脯氨酸、可溶性蛋白含量的影响
图 3a显示,C2处理中菠菜幼苗体内脯氨酸、可溶性蛋白含量较对照(C1)分别增加了 55.4%、17.3%。喷施GA3后(T1-T7处理),T1-T3处理中脯氨酸含量较C2处理分别下降了74.4%、45.4%和40.0%,随后T4-T7处理中的脯氨酸含量逐渐增加,在T5处理时脯氨酸含量达到最大值。从图3b可以看出,T1、T2处理幼苗体内可溶性蛋白含量较C2分别增加了36.9%和49.2%,随后其含量随GA3浓度增加呈递减趋势,在T7处理时,可溶性蛋白含量降至最小值,较C2相比减少了15.7%。

图3 Cu 胁迫及GA3对Cu胁迫下菠菜幼苗脯氨酸(a)和可溶性蛋白(b)含量的影响Fig. 3 Effects of Cu stress and GA3 on proline (a) and soluble protein (b)contents in Spinacia oleracea seedlings under Cu stress
3.4 Cu胁迫及GA3对Cu胁迫下菠菜幼苗Cu吸收的影响
由表 1可知,C2处理中菠菜幼苗叶部、根部Cu含量较对照(C1)显著增加(P<0.05),且叶部Cu含量显著高于根部,其含量为根部的15.5倍。喷施GA3后(T1-T7处理),与C2处理相比,菠菜叶部Cu含量减少,而根部Cu含量增加,其中,T2处理叶部Cu含量降至最小值,较C2处理下降了82.5%,根部Cu含量增至最大值,为C2处理的1.0倍;T3-T7处理叶部Cu含量增加,根部Cu含量有不同程度的下降趋势。
3.5 Cu胁迫及GA3对Cu胁迫下菠菜幼苗矿质元素吸收的影响
由表2显示,与对照(C1)相比,C2处理菠菜幼苗叶部、根部N、P、K、Fe、Zn含量增加,Ca、Mg含量减少。喷施GA3后(T1-T7处理),T1、T2处理叶部N、K、Ca、Mg含量增幅最大,其平均值较C2分别增加了3.4%、84.4%、90.7%和130%,Zn含量降幅最小,其平均值较 C2相比减少了23.8%,而 P含量在 T2-T4处理时增幅最大,Fe含量仅在T1处理时增至最大值;T1、T2处理时,根部矿质元素的变化规律表现为P、K、Ca、Mg、Zn含量增幅最大,其平均值较 C2分别增加了46.4%、92.3%、17.8%、98.2%和39.9%;Fe含量降幅最小,其平均值较C2减少了19.5%;而N含量仅在T2处理增幅最大,较C2相比增加了6.2%。此外,随GA3喷施浓度增加,叶部、根部的7种元素均在T5-T7处理时降至最小值。

表1 Cu胁迫及GA3对Cu胁迫下菠菜幼苗Cu吸收的影响Table 1 Effects of Cu stress and GA3 on Cu absorption of Spinacia oleracea seedlings under Cu stress
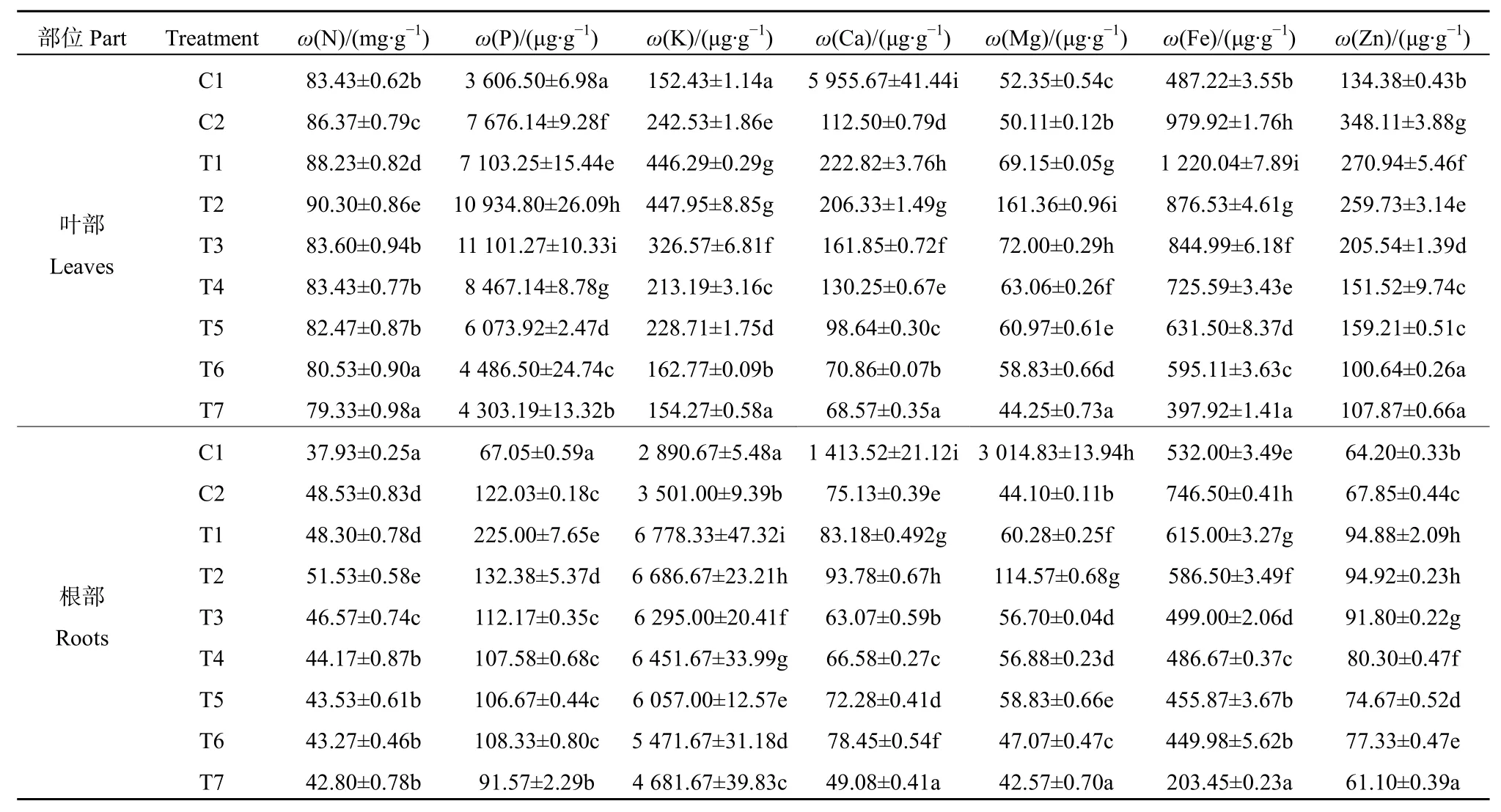
表2 Cu胁迫及GA3对Cu胁迫下菠菜幼苗叶部、根部矿质元素吸收的影响Table 2 Effects of Cu stress and GA3 on the uptake of mineral elements in leaves and roots of Spinacia oleracea seedlings under Cu stress
3.6 Cu胁迫及GA3对Cu胁迫下菠菜幼苗叶部细胞超微结构的影响
对照(C1)处理幼苗叶部细胞中细胞壁、叶绿体、线粒体、核膜、核仁、液泡清晰可见,各细胞器发育良好、结构完整(图4 C1-1和C1-2)。C2处理叶部细胞出现液泡面积扩大、叶绿体肿胀、基粒片层贴靠在叶绿体内膜周围,线粒体体积、数量减少(图 4 C2-1,C2-2)。T1、T3处理叶绿体肿胀状态基本消失,基粒片层堆叠紧密,均匀分布在叶绿体内部,液泡面积缩小,线粒体清晰可见,细胞核逐渐恢复(图4 T1-1-T3-2)。T4、T5处理细胞膜结构仍保持完整,叶绿体结构稍有肿胀,液泡面积逐渐变大,部分细胞出现细胞核解体,线粒体膜呈弥漫性,内脊变得模糊不清甚至断裂(图4 T4-1-T5-2)。T6-T7处理细胞形状变得狭长,叶绿体数量减少,基粒片层堆叠高度下降,细胞核膜表现为核固缩,染色质凝聚形成多核结构和小泡(图 4 T6-1-T7-2)。
4 讨论
4.1 Cu胁迫对菠菜幼苗生长生理特性的影响

图4 Cu胁迫及GA3对Cu胁迫下菠菜幼苗叶部细胞超微结构的影响Fig. 4 Effect of Cu stress and GA3 on ultrastructure of leaf cells of Spinacia oleracea seedlings under Cu stress
Cu是植物生长发育的必需微量元素,植物正常生长所需的Cu含量是5-25 mg·kg-1,当植物体内Cu含量超过30 mg·kg-1时,会对植物产生毒害作用(Krämer,2010;Zheng et al.,2019)。过量的 Cu会导致植物体内产生大量的活性氧(ROS)物质,如过氧化氢(H2O2)、超氧化物()和羟基自由基(-OH)等,这些ROS物质与植物体内的脂类、核酸、蛋白质快速反应形成膜脂过氧化反应,造成细胞膜损伤、酶活性失活,影响细胞的生存能力、抑制植物生长(Arora et al.,2012;Hussain et al.,2014)。MDA是膜质过氧化的重要产物,其含量反映了植物膜脂过氧化程度,与植株受害程度有密切的关系(Rombel-Bryzek et al.,2017)。本研究中,700 mg·kg-1Cu胁迫时(C2处理),菠菜幼苗体内的、H2O2、MDA含量显著增加,生物量明显下降,表明高浓度Cu胁迫能够引起植物产生大量的 ROS物质,扰乱植物对进出细胞物质的控制能力及内稳态的平衡,导致膜质过氧化伤害程度加剧,抑制植物生长(陈镔等,2019)。尽管高浓度Cu胁迫给菠菜幼苗带了如此大的伤害,但是植物体内的脯氨酸、可溶性蛋白含量仍表现出增加趋势,幼苗也处于存活状态,表明该植物具有较强的耐Cu性。但是不同植物的耐Cu性存在差异,有报道显示,铜矿指示性植物海州香薷 Elsholtzia splendens能在600-3000 mg·kg-1Cu的土壤中完成整个生命周期,该植物的根部是吸收和蓄积Cu的主要器官(Jiang et al.,2004;Liu et al.,2005);铜矿区(Cu含量 1158 mg·kg-1)的优势物种白茅 Imperata cylindrical,其根部的蓄Cu能力也强于叶部(Zheng et al.,2019)。本研究结果与上述研究结果不同,700 mg·kg-1Cu胁迫时(C2处理),菠菜幼苗叶部、根部Cu含量显著增加,且叶部的Cu含量显著高于根部,表明叶部是菠菜幼苗蓄Cu的主要器官,虽然本研究中菠菜生长环境中的Cu浓度不及上述两种植物高,但也能表现出较强的耐Cu性。
此外,Cu胁迫也会干扰植物对矿质元素的吸收和运输,导致植物出现养分不平衡、生理功能紊乱等毒害症状(Zhang et al.,2010)。本研究结果显示,与正常生长的幼苗相比(C1),700 mg·kg-1Cu胁迫时(C2处理),菠菜幼苗叶部、根部N、P、K、Fe、Zn含量均有增加趋势,这可能是由于高浓度Cu胁迫下,植物启动体内抗氧化防御系统,抵御胁迫环境,主动吸收、积累大量的矿质元素,维持其生命活动,这也是菠菜具有较强耐Cu性的一种表现。Ca、Mg是植物生长发育的必需元素,Ca具有保护细胞膜、促进植物生长的作用(Tuna et al.,2008),Mg是叶绿素分子的中心原子,也是蛋白质合成过程中核糖体亚单位聚集的重要元素(Shaul,2002;Souza et al.,2014)。本研究中,高浓度Cu胁迫导致菠菜体内Ca、Mg含量减少,表明这两种元素相对于其他矿质元素可能更容易受到Cu胁迫的影响,从而导致其吸收量减少,或是由于细胞膜损伤、膜透性增加造成离子外流,最终在植物体内表现出 Ca含量下降,对细胞膜的保护作用减弱;而Mg含量下降影响植物叶绿素的合成,植物光合作用受到抑制(Shaul,2002;Tuna et al.,2008)。
细胞是植物生命活动的基本单位。在重金属胁迫下,植物体内的超微结构也会因重金属的浓度和胁迫时间而发生改变(徐义昆等,2015)。本研究结果显示,正常生长情况下(C1),菠菜幼苗叶部细胞内细胞壁、细胞核、核膜、核仁清晰可见,叶绿体内外膜完整,基粒片层排列整齐,液泡居于细胞中部。700 mg·kg-1Cu胁迫时(C2处理),菠菜幼苗叶部细胞内细胞核消失,线粒体数量减少,液泡面积扩大、叶绿体肿胀、基粒片层贴靠在叶绿体内膜周围,其原因可能是高浓度Cu2+进入细胞,细胞膨压增大所致(Peck et al.,2010);其次,高浓度Cu也抑制了菠菜幼苗细胞内线粒体合成,导致其体积、数量减少。相同研究结果在同类试验中也出现,如Cu胁迫导致石花菜 Gelidium floridanum叶肉细胞壁厚度增加、叶绿体解体、线粒体肿胀(Dos Santos et al.,2014);可可Theobroma cacao植物叶部受到Cu胁迫后叶绿体膜破裂、类囊体肿胀(Souza et al.,2014)。
4.2 GA3对 Cu胁迫下菠菜幼苗生长生理特性的影响
重金属胁迫时,外源添加GA3能够减少植物吸收重金属,减缓重金属对植物的胁迫伤害。本研究结果显示,向Cu胁迫菠菜幼苗叶部喷施浓度低于5 mg·L-1的 GA3溶液时(T1、T2处理),幼苗体内的、H2O2含量降低,可溶性蛋白含量、生物量增加,表明浓度低于5 mg·L-1的GA3溶液,能降低Cu胁迫对菠菜幼苗的膜质过氧化伤害,抑制 ROS物质的产生,促进植物生长(Zhu et al.,2012;Ben Massoud et al.,2017)。但继续增加GA3浓度,菠菜体内的H2O2、MDA含量显著增加,脯氨酸含量也增加,而可溶性蛋白含量降低,菠菜幼苗鲜重、干重降低,表明高浓度的GA3会加剧Cu胁迫对菠菜幼苗的胁迫伤害,抑制可溶性蛋白的合成,降低其生长量,但是植物仍能增加脯氨酸含量抵御胁迫伤害(Wang et al.,2015;Muniandi et al.,2018)。此外,喷施浓度为5 mg·L-1GA3溶液时,菠菜叶部Cu含量降至最小值,而根部Cu含量增至最大值,表明5 mg·L-1GA3浓度能减少菠菜叶部吸收、积累Cu2+(Zhu et al.,2012),缓解Cu胁迫对幼苗叶部的毒害。类似结果在同类研究中也有出现,如1 mmol·L-1GA3能减轻 Cu 对豌豆 Pisum sativum 幼苗的伤害(Ben Massoud et al.,2017),100 mg·L-1GA3能减轻Cd2+和Pb2+对蚕豆Vicia faba和羽扇豆Lupinus植物的危害 (Abd et al.,2009)。但继续增加GA3浓度,菠菜叶部Cu含量降幅减少,根部Cu含量增幅也减少,表明GA3浓度增加促进了菠菜幼苗叶部对 Cu2+的吸收,而减弱了根部 Cu2+积累,其原因可能是高浓度 GA3对细胞分裂、加速生长的促进作用过强,导致细胞膜透性增加,Cu2+顺势进入根部细胞,植物为减轻大量 Cu2+聚集对根部细胞的伤害,主动将 Cu2+迁移至叶部,从而导致叶部Cu含量多于根部(Hadi et al.,2014;Ji et al.,2015;邵君伟,2015)。
矿质元素是参与植物生理代谢的重要物质(Zhang et al.,2010)。非生物胁迫条件下,GA3能够改变植物对矿质元素吸收和运输(Tuna et al.,2008)。在本研究中,喷施浓度低于 5 mg·L-1的GA3时(T1、T2处理)时,与Cu胁迫(C2处理)相比,菠菜幼苗叶部N、K、Ca、Mg含量增幅最大,Zn含量降幅最小,根部P、K、Ca、Mg、Zn含量增幅最大,Fe含量降幅最小,表明Cu胁迫下,低于5 mg·L-1的GA3处理能促进菠菜幼苗吸收矿质元素,这可能是GA3降低了Cu胁迫对植物细胞膜的氧化损伤,细胞膜的完整性得到了恢复,植物吸收的矿质元素能够保存在细胞内,也可能是GA3调节改变了植物对矿质元素的吸收通道,缓解了Cu2+对等价矿质元素吸收的拮抗效应(Ashraf et al.,2002)。但随 GA3浓度继续增加,菠菜幼苗叶部、根部吸收矿质元素含量下降,其原因可能是GA3浓度过高导致菠菜幼苗体内的氧化伤害加剧,植物体内产生了大量的ROS物质,造成细胞膜透性增加,矿质元素外流(Hadi et al.,2014;Ji et al.,2015)。
植物叶片组织和细胞器结构会因植物生长环境和外界刺激而发生变化(Du et al.,2017),叶绿体是植物进行光合作用的重要细胞器,其结构的完整性是植物进行高效光合的前提(黄晓霞等,2017)。本试验结果显示,与 Cu胁迫相比,喷施浓度低于5 mg·L-1的GA3时(T1、T2处理),菠菜幼苗叶部细胞叶绿体肿胀状态基本消失,基粒片层堆叠紧密,均匀分布在叶绿体内部,液泡面积缩小,线粒体清晰可见,细胞核逐渐恢复,表明低浓度的GA3溶液能缓解Cu胁迫对植物细胞伤害,促使细胞结构恢复正常,从而使细胞器较正常地发挥生理功能。但继续增加GA3浓度,菠菜幼苗细胞中的叶绿体、线粒体、细胞核数量出现不同程度的下降,结构模糊不清等,这可能是GA3添加浓度过高逆转了其对Cu胁迫菠菜幼苗的缓解作用,导致胁迫伤害加重(Muniandi et al.,2018)。
5 结论
700 mg·kg-1Cu胁迫时,菠菜幼苗体内Cu含量显著增加、氧化伤害加剧,生长量和Ca、Mg含量均受到抑制,细胞器结构损伤,但脯氨酸和可溶性蛋白含量增加,N、P、K、Fe、Zn含量升高,植物也处于存活状态,能够表现出较强的耐Cu性。喷施低于5 mg·L-1的GA3能减少Cu胁迫下菠菜幼苗叶部Cu含量,缓解氧化伤害,增加可溶性蛋白含量,促进矿质元素吸收,增加植物生物量,同时,叶绿体肿胀消失、液泡面积缩小,线粒体清晰可见,细胞核逐渐恢复。而继续增加GA3浓度时,幼苗叶部Cu含量增加、氧化伤害加剧,可溶性蛋白和矿质元素含量减少,植物生长受到抑制,叶绿体、线粒体、细胞核数量减少,结构模糊不清。因此,可以推断低于5 mg·L-1的GA3能缓解Cu胁迫对菠菜幼苗的毒害,继续增加GA3浓度会导致Cu胁迫伤害加剧。
猜你喜欢
中国甜菜糖业(2022年4期)2023-01-21
农业工程学报(2022年6期)2022-06-27
中国烟草科学(2022年2期)2022-05-27
少儿科技(2019年9期)2019-09-10
湖北农业科学(2018年18期)2018-12-11
中成药(2018年11期)2018-11-24
现代园艺(2017年21期)2018-01-03
中国糖料(2016年1期)2016-12-01
植物营养与肥料学报(2012年1期)2012-10-26
中国烟草学报(2012年2期)2012-04-09
