
青藏高原东缘高原鼢鼠种群抗药性评估
2019-12-12 07:31谭宇尘韩天虎许国成魏彦明蔡志远姚宝辉郭怀亮苏军虎
草业科学 2019年11期
谭宇尘,韩天虎,许国成,魏彦明,蔡志远,王 缠,姚宝辉,郭怀亮,苏军虎
(1. 甘肃农业大学生命科学技术学院,甘肃 兰州 730070;2. 甘肃省草原技术推广总站,甘肃 兰州 730020;3. 甘肃农业大学动物医学院,甘肃 兰州 730070;4. 甘肃农业大学草业学院,草业生态系统教育部重点实验室,中美草地畜牧业可持续发展研究中心,甘肃 兰州 730070)
抗药性是指药物在治疗疾病或病症时药物治疗效果降低,是生物对周围所处环境的变化和化学药品影响而做出自身适应的结果[1-2]。生物会形成获得性耐药性,即在药物的使用过程中,由于药物反复投喂,用药量不足或滥用药物导致其对药物产生了抵抗[2]。化学方法防治啮齿动物过程中,通常使用急性杀鼠剂、生物类蛋白杀鼠剂和抗凝血剂类杀鼠剂[3]。抗凝血剂类杀鼠剂是典型的慢性杀鼠剂,对家畜和鸟类毒害作用小,是最为广泛应用的一类杀鼠剂[4-5]。长期使用抗凝血剂类药物对啮齿动物进行防治会使其产生拒食、代谢应对以及基因突变等行为、生理和遗传等抗性机制[6-8]。Boyle和Mary[9]在瑞典检测出有部分鼠类在抗凝血剂类药物投喂过程中有抗药性产生,此后西欧和北美也检测到抗性种群的产生。19世纪50年代起,我国开始使用抗凝血剂类药物控制啮齿动物,在抗凝血剂类药物对害鼠的防治过程中多次检测到害鼠产生抗药性[10]。
抗凝血剂类药物的作用方式是通过抑制维生素K环氧化物复合物还原酶亚基1 (vitamin K epoxide reductase complex subunit 1,Vkorc1)来抑制血液的凝固,增加血管的通透性,使啮齿类动物内出血而死亡[11-12]。Vkorc1基因突变中已有5种特异性氨基酸突变使啮齿类动物对抗凝血剂类药物有抗药性,从而构成了啮齿动物的抗性个体,Vkorc1基因突变使不同地区的啮齿动物种群出现不同程度的抗性[13]。了解Vkorc1基因序列变异及对其编码蛋白质的结构突变,是研究啮齿动物是否产生抗药性的方法之一。研究发现,具有遗传基础的抗药性与Vkorc1基因的外显子突变有关[14],已检测出的啮齿动物体内Vkorc1氨基酸的多态性Ser56Pro、Trp59Arg、Phe63Cys、Leu120Gln和Tyr139Cys会增加对抗凝血剂类药物的抗药性[15]。
高原鼢鼠(Eospalax baileyi)隶属于啮齿目、鼹形鼠科、鼢鼠亚科、鼢鼠属,是青藏高原特有的物种之一,在繁殖活动时期大量啃食作物根茎、树皮草根等,采食植物的地下根系,也常将植物地上部分的茎叶拖入洞道内食用或作巢内铺垫之用,对农作物和牧区造成严重的影响[16]。长期以来人们对高原鼢鼠多采用药物防控和人工捕捉的方式,防控药物多采用溴敌隆(鼢鼠灵)等抗凝血剂类药物[11],但有关高原鼢鼠抗药性监测的研究缺乏。科学有效掌握高原鼢鼠的抗药性情况,对高原鼢鼠的管理具有重要意义。本研究在青藏高原东缘的天祝、甘南等地采集了有抗凝血药物投放历史的870只高原鼢鼠样本,检测高原鼢鼠Vkorc1基因第一外显子、第二外显子和第三外显子,分析了其核苷酸变异情况,并结合已发现的抗药性变异位点结合分析其抗药性,评估青藏高原地区高原鼢鼠种群抗性水平,旨在为青藏高原地区高原鼢鼠的科学有效管理提供参考[10]。
1 材料与方法
1.1 高原鼢鼠样本采集
于2017-2018年,在青藏高原东缘的武威市天祝藏族自治县、甘南藏族自治州的碌曲、合作、临潭和卓尼县和定西市岷县等有抗凝血药物投放10年以上的地区,弓形夹捕捉高原鼢鼠,称重后按体重来鉴别年龄信息[17],选择全部为成年以上的个体,总的比例雌雄各半,采集尾端样本(尾巴末端3~4 cm的部位)经烘干后室温保存。所有高原鼢鼠参考相关文献进行经物种鉴别后,再进行Vkorc1基因测序[18],具体样地信息如表1所列。
1.2 引物设计试验方法
1.2.1 引物设计
通过将高原鼢鼠的DNA序列与在NCBI上已发表的褐家鼠序列(NM_203335.2)和亲缘关系较近的同科物种裸鼹鼠(Heterocephalus glaber)序列(XM_004856224.3)的Vkorc1基因序列进行序列对比分析。用Primer3Input (Version 0.4.0) 软件对高原鼢鼠Vkorc1基因外显子可能存在SNP的片段设计引物(表2)。引物序列由西安擎科泽西生物科技有限有限公司合成。

表 2 高原鼢鼠3个外显子部位引物序列Table 2 Primers sequence of 3 exons in plateau zokor
1.2.2 PCR扩增产物处理及基因型分析
PCR扩增产物初步经聚丙烯酰胺凝胶电泳SSCP-PAGE检测样本基因型的SNP。利用已成功扩出的Vkorc1基因的3个外显子进行扩增。反应包含10 × buffer缓冲液,0.5 μL的dNTP,0.3 μL的Taq DNA聚合酶,1 μL的引物和1 μL的模板DNA,反应体系为25 μL。
1.3 数据分析
高原鼢鼠Vkorc1基因外显子PCR结果使用琼脂糖凝胶电泳进行检测,PCR产物经非变性聚丙烯酰胺凝胶电泳SSCP-PAGE筛选过后送至西安擎科泽西生物技术有限公司进行测序。所得高原鼢鼠Vkorc1基因外显子测序结果用OLIGO7.0进行编辑比较。
高原鼢鼠Vkorc1氨基酸突变位点使用MEGA7.0进行比对分析,所得突变的高原鼢鼠氨基酸三维模型使用SWISS-MODEL进行预测细化和评价,并使用GOR IV对蛋白质二级结构检查比较[19]。
2 结果
2.1 高原鼢鼠Vkorc1基因外显子扩增
高原鼢鼠Vkorc1基因外显子PCR结果使用琼脂糖凝胶电泳进行检测,得到高原鼢鼠3个外显子基因序列,第一外显子为147 bp,第二外显子为106 bp,第三外显子为157 bp (图1)。所得到合适的高原鼢鼠Vkorc1外显子使用非变性聚丙烯酰胺凝胶电泳进行检测,得到了高原鼢鼠3个外显子的SNP(图2)。

图 1 图1高原鼢鼠外显子PCR产物琼脂糖凝胶电泳结果Figure 1 Agarose gel electrophoresis results of Vkorc1 exons 1-3 PCR products in plateau zokor

图 2 高原鼢鼠3个外显子聚丙烯酰胺凝胶电泳SNP结果Figure 2 SNP results of polyacrylamide gel electrophoresis in three exons of plateau zokor Vkorc1
2.2 高原鼢鼠Vkorc1基因外显子多态性SNP分析
在全部870个高原鼢鼠样本的Vkorc1基因外显子多态性结合测序图谱分析,得出各基因型突变位点及突变频率(表3),以及高原鼢鼠不同地区外显子基因型数量分布(表4)。

表 3 高原鼢鼠Vkorc1基因外显子多态性突变位点及突变频率Table 3 Polymorphic mutation sites and frequency of Vkorc1 gene exons 1-3 in plateau zokor
2.3 高原鼢鼠Vkorc1氨基酸序列检测及抗药性
使用SWISS-MODEL进行分析发现,在870个高原鼢鼠个体中,有197个个体发现Vkorc1氨基酸序列与野生型有差异(图3)。
Vkorc1基因3个外显子中第一外显子BB基因型,第二外显子AB基因型,第三外显子BB基因型这3种最常见的基因型在每一个受样地的样本中占很大比例。天祝、碌曲、临潭和卓尼地区检出了全部的基因型,其中包括导致抗性发生的Phe63Cys氨基酸突变。这种Vkorc1氨基酸的突变的三维模型预测到Phe63Cys的侧链冲突而导致蛋白质空间构象的改变。Vkorc1基因第三外显子BB基因型中对比褐家鼠有氨基酸突变Ala143Val,这种基因型的突变也能导致蛋白质空间构象的改变(图4)。合作地区检测出了Gly19Glu和Leu114Ile的氨基酸突变,但缺少Arg61Pro氨基酸突变的基因型和能致抗性发生的Phe63Cys氨基酸突变基因型(表5)。
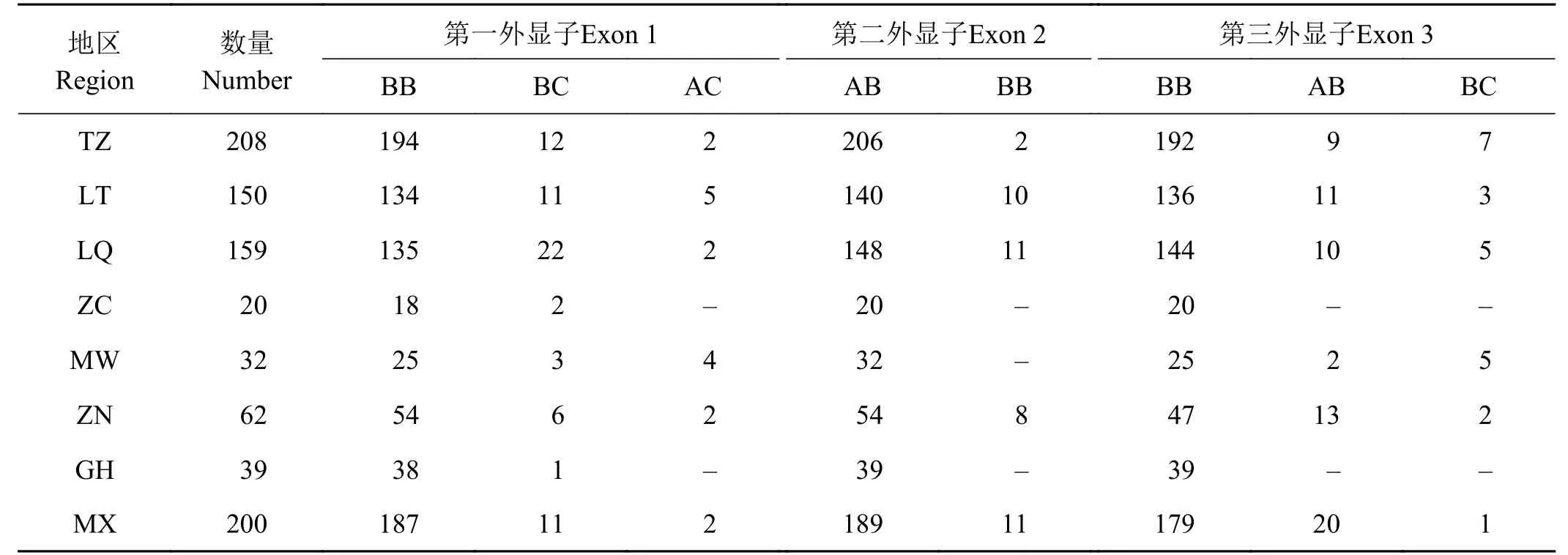
表 4 高原鼢鼠不同地区外显子基因型数量分布Table 4 SNP results of polyacrylamide gel electrophoresis in three exons of plateau zokor Vkorc1

图 3 高原鼢鼠Vkorc1基因4个单核苷酸多态性位点Figure 3 Four single nucleotide polymorphic loci of Vkorc1 gene in plateau zokor
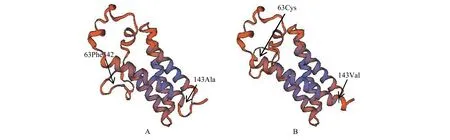
图 4 高原鼢鼠的维生素K环氧还原酶的三维模型Figure 4 Three-dimensional model of vitamin K epoxide reductase in plateau zokor

表 5 高原鼢鼠检测到的4个氨基酸变异的概况Table 5 Overview of the four Vkorc1 amino acid variants detected in plateau zokor
在氨基酸突变的基因型遗传偶联上发现,在碌曲和合作样地的样本中共有52只互为亲缘关系的高原鼢鼠,并发现其中有11只的第一外显子杂合子AC基因型的交换组合,以及有7只第一外显子的杂合子BC基因型的Gly19Glu氨基酸突变。第一外显子的杂合子BC基因型在与其他氨基酸序列交换时仅在2只高原鼢鼠中检测到,第三外显子的杂合子AB基因型在与其他氨基酸序列交换时在8只高原鼢鼠中检测到。
结合DNA-PCR产物测序和PCR-SSCP方法,经过MAGA7.0软件比对分析结果发现,在高原鼢鼠目标群体中VKORC1基因外显子翻译区序列有4个种间突变位点,对比褐家鼠序列,高原鼢鼠有18个错义突变位点,第一外显子有8个氨基酸突变位点(Ser7Asn、Arg10Trp、Leu11Val、Leu17Val、Leu20Phe、Asn36Asp和Glu37Lys);第二外显子有3个氨基酸突变位点(His68Pro、Ala72Met和Ile90Leu);第三外显子有8个氨基酸突变位点(Gly99Val、Ile107Val、Leu108Val、Val114Leu、Leu118Val、Ala143Val、Met146Thr和Leu147Trp)。在记录的其他4种能导致啮齿动物产生抗药性基因型的氨基酸突变(Ser56Pro、Trp59Arg、Leu120Gln、Tyr139Cys),在高原鼢鼠中未检测到(图5)。870只高原鼢鼠中大多数都匹配裸鼹鼠Vkorc1基因的外显子序列,然而89.2%的高原鼢鼠在第一外显子具有Pro8Pro、Gly9Gly、Arg12Arg和Gly46Gly同义SNP,95.2%高原鼢鼠在第二外显子具有Gly64Gly、Trp66Trp和Glu67Glu同义SNP,88.2%高原鼢鼠在第3外显子具有Arg99Arg同义SNP。
有41只高原鼢鼠有与Phe63Cys突变相关的核苷酸置换,显示出抗药性发生情况,占全部高原鼢鼠的4.8%,其中天祝有2只,抗性发生率为1.0%(n = 208),碌曲种群检测到10只,抗性发生率为6.3% (n = 159),临潭种群有10只,抗性发生率为6.7% (n = 150),卓尼种群有8只,抗性发生率为12.9% (n = 62),岷县种群有11只,抗性发生率为5.5% (n = 200) (表4)。

图 5 高原鼢鼠对比褐家鼠氨基酸错义突变位点Figure 5 Comparison of amino acid missense mutation sites between plateau zokor and Rattus norvegicus
3 讨论
本研究发现,青藏高原东缘高原鼢鼠Vkorc1基因3个外显子具有多态性位点,以及Vkorc1氨基酸序列的发生的突变,根据氨基酸序列分析,结合已知突变位点和抗药性的相关性,说明Vkorc1基因介导的维生素K还原氧化酶的改变使高原鼢鼠对抗凝血剂类药物具有抗药性,并初步发现了4个抗性(地理)种群。
3.1 高原鼢鼠Vkorc1基因 Phe63Cys基因型的突变影响抗药性
研究发现啮齿类动物具有5个产生抗药性的氨基酸位点,关键位点的氨基酸取代可能会干扰底物或成为Vkorc1酶的抑制剂,对抗凝血剂类药物产生活性的改变。而蛋白表达分析Vkor酶更能准确地反映高原鼢鼠体内检测到的抗性条件,从而发现这些Vkorc1基因突变与物种是否无关。对高原鼢鼠的这五个氨基酸位点进行检测发现,在天祝、碌曲、临潭、卓尼和合作美吾捕获的高原鼢鼠中都检测出了Vkorc1氨基酸的Gly19Glu、Arg61Pro、Phe63Cys和Leu114Ile的错义突变位点。在天祝、碌曲、临潭和卓尼地区的539只高原鼢鼠中部分样本的Vkorc1基因携带纯合子Phe63Cys基因型。研究发现影响挪威鼠(Rattus norvegicus)对抗凝血剂类药物产生抗性相应突变的个体中显示,携带纯合子Phe63Cys的啮齿类动物对抗凝血剂类药物有较强的抗性[19]。且Phe63Cys突变在屋顶鼠(Rattus rattus)体内表达比较大时,使Vkorc1酶活性低于正常屋顶鼠(野生型)体内酶的表达水平,并导致其抗药能力提高6倍[20]。
在其他啮齿动物中已有报道,并在高原鼢鼠Vkorc1基因序列中发现的单核苷酸突变(Phe63Cys),已证明与抗凝血类药物的抗药性有关,但所检测出的抗性高原鼢鼠数量在所有检测样本中占4.8%,说明已出现抗性个体但尚未形成抗性种群。在美吾、则岔、尕海和果芒等地高原鼢鼠与褐家鼠种间比较发现氨基酸不同,在其他啮齿类中未见报道。在高原鼢鼠中发现的5种氨基酸突变中,但这5种氨基酸突变与抗凝血类药物的抗药性没有关系。研究发现,啮齿动物虽会产生同义替换,但不会对抗凝血剂类药物产生抗药性。此外,Phe63Cys的突变比例并不高[21]。本研究发现,只有41只高原鼢鼠有与Phe63Cys突变相关的核苷酸置换,占全部高原鼢鼠的4.8%,41只高原鼢鼠中有2只分布在天祝,10只分布在碌曲,10只分布在临潭,8只分布在卓尼,11只分布在岷县。这对于当地高原鼢鼠种群数量的控制是一种挑战,在有抗性高原鼢鼠个体产生的情况下,当地应使用多种不同的药物来控制高原鼢鼠。或是从已检测到相关氨基酸Phe63Cys突变的地方捕获活的高原鼢鼠,通过凝血反应测试来验证高原鼢鼠的实际抗性情况[22]。
3.2 高原鼢鼠Vkorc1基因其他基因型的突变影响抗药性
全球许多国家都有关于啮齿类动物对抗凝血剂类药物产生抗药性的研究。通常抗凝血剂类药物诱饵的广泛应用和重复使用,以及诱捕实践的效率低下,被认为是产生广泛抗药性的原因之一。Vkorc1基因突变是啮齿类动物对抗凝血剂类药物产生抗药性的主要原因之一,这种基因突变可与维生素K依赖性凝血因子缺乏相关[23]。一种方法是对啮齿类动物的Vkorc1基因进行变异位点分析,从而发现是否有其基因在抗凝血类药物的使用中出现了突变,从而解析出啮齿动物对抗凝血剂类药物是否产生抗药性[24-25]。另一种方法则是用啮齿类动物Vkorc1基因序列介导翻译的氨基酸在毕赤酵母中的表达和抑制常数,来测定啮齿类动物对抗凝血剂类药物的抗药性[26]。
研究表明,Vkorc1基因中其他许多突变也可使啮齿类动物对抗凝血剂类药物产生抗药性。如Buckle等[27]发现,在褐家鼠Vkorc1基因上的第139位酪氨酸产生变异的个体均会对抗凝血剂类药物产生不同程度的抗药性。Buckle等[28]发现Vkorc1第一外显子基因序列在第74个碱基上发生A >T碱基替换的变异也会导致氨基酸的突变,这种突变可能通过改变蛋白质的螺旋结构造成蛋白质部分功能的缺失,从而导致啮齿类动物对各种抗凝血剂类药物产生抗药性。本研究中,在临潭和天祝所捕获的高原鼢鼠样本中均未有这种突变发生。在其他研究中,尚未发现Vkorc1基因序列和Vkorc1表达的氨基酸序列中与抗凝血抗药性相关的其他基因型突变[29]。
3.3 高原鼢鼠抗药性分子检测及其治理
啮齿类抗药性传统的检测多用敏感性测定,该方法需要进行捕捉、饲养和适口性分析等[2],很难进行大规模的种群抗性评估,而生理生化酶活性的测定方法也需要进行采集血样等进行分析,亦具有过程复杂等特点。随着分子生物学技术的发展,获得啮齿动物DNA的技术和手段日趋完善和简便,这为分子检测提供了极大的便利。本研究在前期研究的基础上,候选溴敌隆的抗性基因,并结合已知的变异及其抗药相关性,首次对青藏高原东缘8个地理种群近900只个体进行了检测,较全面的掌握了高原鼢鼠的抗性情况。但对来自新型Vkorc1基因变异群体的啮齿类动物进行血凝反应测试阐明其抗药性状况,还需要进一步研究以确定其抗药性机制。同样,其他Vkorc1基因型的改变也可能在其他啮齿动物对抗凝血剂类药物产生抗药性中发挥作用。在分子抗性水平上的其他研究发现,细胞色素氧化酶P450 (Cytochrome P450, P450)基因的突变也可导致啮齿类动物的抗药性增加。抗凝血剂类药物主要由细胞色素P450基因编码的酶参与的新陈代谢被代谢出体外。截至目前,细胞色素P450 CYP2C9 (Cytochrome P450 2C9)基因中有30多个等位基因的等位变异与酶的活性有关,因此具有CYP2C9野生型等位基因的个体更易对抗凝血剂类药物敏感[30]。同样,常见的携带有CYP2C9纯合基因型的抗凝血剂类药物给予剂量要高于CYP2C92和CYP2C93等其他基因型[31]。通过对细胞色素P450基因进行扩增和测序,可以确定高原鼢鼠是否有因P450基因上的表达异常而对药物产生抗药性。
一旦停止对抗凝血剂类药物的广泛使用,对溴敌隆等其他抗凝血剂类药物有抗药性的个体就可以从种群中逐步淘汰出来。在对挪威鼠的抗药性检测中有证据表明,当停止使用抗凝血剂类药物时,啮齿类动物种群的抗性基因频率会逐年下降[32],携带抗药性基因的啮齿动物的增速度下降,从而影响到它们的大规模繁殖。然而,另一项研究发现,对抗凝血剂类药物抵抗能力强的个体比对药物敏感强性的的个体体重更重,在没有药物选择压力的情况下,携带抗药性基因的啮齿类动物种群数量并没有发生太大的变化[33]。使用交叉使用类药物或是间断使用抗凝血剂类药物,都可以预防或减轻高原鼢鼠对抗凝血剂类药物的抗药性,这将为抗药性的解决提供理论依据。因此,针对抗药性种群的出现,首先进行必要的抗药性监测,在认识抗性种群结构和抗性水平的基础上采用药物轮换使用,或使用生物防治如加大采用C型、D型肉毒素的防治或引入狐狸(Vulpes)、鹰(Accipiter)天敌等其他防治方法,都能有效控制高原鼢鼠的种群数量。其次从环境治理方面,结合草地管理措施开展(清理高原鼢鼠的活动繁殖场所,或者通过草地改良补播等)基础综合控制和防治重点场所设施的完善,或者开展新药和新剂型的发展[34],都可以对高原鼢鼠的种群数量进行有效的控制。
猜你喜欢
电子科技大学学报(2022年5期)2022-10-29
Zoological Research(2022年3期)2022-06-07
南方农业(2021年30期)2021-12-25
中国生殖健康(2020年4期)2021-01-18
兽医导刊(2020年3期)2020-12-30
农家科技中旬版(2020年5期)2020-06-15
世界农药(2019年3期)2019-09-10
农药科学与管理(2019年10期)2019-04-20
中国生殖健康(2018年4期)2018-11-06
新农业(2017年2期)2017-11-06
