
猪链球菌2型通过诱导IFN-γ促进脑微血管内皮细胞自噬破坏血脑屏障完整性
2020-02-24 08:21刘嘉楠姜合祥雷连成
中国兽医学报 2020年12期
刘嘉楠,贾 丽,姜合祥,吴 桐,李 扬,雷连成,3*
(1.吉林大学 动物医学学院, 吉林 长春 130062;2.吉林大学 护理学院, 吉林 长春 130062;3.长江大学 动物科学学院,湖北 荆州 434023 )
由猪链球菌2型(Streptococcussuistype 2,SS2)引起的脑膜炎可导致高死亡率和严重后遗症,这对养猪业和公共环境卫生管理构成了巨大威胁[1]。然而,目前仍然没有有效的治疗方法。SS2诱导脑膜炎的关键步骤是黏附和通过血脑屏障(BBB)的侵袭,引发中枢神经系统感染和炎症反应[2]。因此,进一步探索其发病机制将有助于对猪链球菌型脑膜炎进行有效的靶向治疗。
脑微血管内皮细胞/星形胶质细胞(BMEC/AC)共培养BBB模型广泛用于脑膜炎机制研究[3]。与单个BMEC模型相比,BMEC/AC共培养BBB模型更好地模拟了BBB的实际环境[4]。通过内皮电阻(TEER)评价所构建屏障模型的紧密性,高TEER值意味着单层细胞融合良好[5]。
当SS2通过血液循环到达BBB时,它可刺激BMEC产生细胞因子并改变BBB的完整性,并进一步穿过BBB侵入神经系统。据报道,SS2于刺激猪脑微血管内皮细胞(PBMEC)产生炎症前细胞因子。一般而言,细胞因子具有许多功能,如调节先天免疫和适应性免疫[6],甚至在抗菌和抗病毒感染中发挥重要作用。然而,过量的细胞因子,被称为“细胞因子风暴”,会给机体造成更大的危害[7]。SS2感染后产生的细胞因子类别及其功能尚不清楚。如果可以及时监测细胞因子的反应,可以更好地选择药物并确定药物治疗时间,从而实现更好的治疗效果。
IFN-γ是由T细胞和NK细胞分泌的多效细胞因子,可参与许多炎症性疾病的发病机制。最近的报道表明,IFN-γ通过激活免疫系统发挥抗肿瘤作用。然而,在某些条件下,IFN-γ的作用是有限的,甚至会促进疾病的发生和发展。最近的一项研究表明,IFN-γ诱导牛乳腺上皮细胞的自噬和精氨酸耗尽,精氨酸的加入可以逆转IFN-γ诱导的乳腺上皮细胞的恶性转化[8]。在SS2感染过程中,IFN-γ的表达也显著增加。因此推测IFN-γ可在SS2感染下引起BMECs自噬。
在本研究中,先以猪脑切片为模型,检测SS2感染后引起的损伤以及炎性细胞因子的变化情况,后针对炎性细胞因子对细胞造成的损伤进行进一步研究。并且成功分离了猪原代BMEC和ACs,构建BMEC/AC共培养BBB模型,便于更好地模拟体内BBB生理学状态。使用ELISA和RT-PCR方法鉴定SS2感染后BBB模型细胞因子的变化。并研究在IFN-γ处理下猪脑切片,BBB模型和PBMEC的反应,证明IFN-γ可诱导PBMECs自噬并破坏BBB完整性。添加精氨酸可拮抗细胞自噬和IFN-γ诱导的BBB损伤,为SS2脑膜炎的预防和治疗提供新的见解及药物选择,构建的BBB共培养模型也为SS2脑膜炎和BBB治疗提供了有效研究基础。
1 材料与方法
1.1 主要试验材料猪链球菌CVCC606购自中国微生物菌种保藏中心;Brain Heart Infusion(BHI)培养基购自海博生物;PBMEC/ACs分离自2周龄健康仔猪;DMEM/F12培养基、双抗(青链霉素)购自Biological Industries;胎牛血清(FBS)购自Clark;RIPA buffer 购自碧云天;GAPDH抗体、LC3B抗体、HRP标记的山羊抗小鼠IgG、山羊抗兔IgG均购自Protein tech;猪重组IFN-γ蛋白购自GENWAY。
1.2 PBMEC/AC分离培养与BBB模型建立健康仔猪麻醉安乐致死后将头分开并消毒。从颅骨中取出脑组织,并用无菌手术刀将其切成2 mm2厚的组织片。将组织转移到50 mL试管中,用预冷的CM (98% DMEM/F-12+2% FBS+1% penicillin/streptomycin) 洗涤2次,重悬后以4℃、400 r/min离心5 min。弃去上清液,将细胞沉淀物用等体积的25%牛血清蛋白(BSA)重悬,4℃、1 000 r/min离心15 min。离心后,收集沉淀,加入25% BSA重悬,进一步清洁,再次离心后,细胞培养基重悬。培养60 min 后,将含有悬浮细胞的细胞培养基移至另一个细胞培养瓶中进行培养即为AC,并将先前的细胞瓶添加到新的5 mL细胞培养基中以进行PBMEC培养。
共培养的BBB Transwell模型用分离的原代PBMEC和AC构成。首先,将Transwell放入培养皿中,加入细胞培养基润湿Transwell。将第3次传代后的AC以2×104/cm2的细胞浓度接入Transwell,使AC黏附3 h,然后将Transwell倒置插入并置于12孔细胞培养皿中。将0.5 mL细胞培养基添加到底部腔室中,并将1.5 mL细胞培养基添加到基底外侧腔室中。3 d后,将PBMEC以2×104/cm2的浓度接入Transwell的上部。每2~3 d更换细胞培养基。共培养BBB模型的试验在两种细胞接种后的7~8 d内进行。对照组包括单个PBMEC构建的BBB模型和DMEM/F-12组(无细胞对照组)。
1.3 Real-time qPCRTRIzol(Invitrogen)裂解PBMEC/AC共培养BBB模型或PBMECs,氯仿提取RNA (Fisher Scientific)。使用Prime ScriptTMRT Master Mix Kit (TaKaRa)反转录,SYBR Green(TaKaRa)进行RT-PCR检测。用2-△△Ct法测定靶基因的相对转录水平,基因引物列见表1。
1.4 Western blot常规方法制备样品,电泳,将凝胶上的蛋白条带转印至甲醇激活的PVDF膜,5%脱脂乳室温封闭1 h,加入相应的一抗4℃孵育过夜,TBST洗膜,加入相应二抗室温下孵育1 h,TBST洗膜,滴加ECL化学发光试剂,Tanon成像系统显影。
1.5 引物设计和合成本研究中用到的荧光定量PCR引物如表1所示,均由吉林省库美生物科技有限公司合成。
1.6 SS2培养与感染将SS2 606株在BHI中培养至D600= 0.6(CFU=1×108/mL),收集1 mL菌液,离心并用PBS洗涤2次,感染脑组织切片。
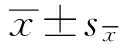
2 结果
2.1 SS2感染诱导IFN-γ高表达为探究SS2感染后,脑组织损伤情况以及所引起的炎症反应,首先选用脑组织切片,检测SS2感染后炎性细胞因子的变化情况。切取仔猪脑组织,制备脑切片培养,并对其活性进行评价。随着培养时间的延长,脑组织切片逐渐清晰透明,切片边缘有明显增生(图1A);并对培养至5 d的脑切片进行PI染色,发现脑切片并无红染(图1B);LDH试剂盒检测发现脑切片的LDH在1 d释放量较高,从3 d起逐渐降低并趋于稳定(图1C),以上结果表明脑切片活性良好。

表1 Real-time qPCR引物及序列
选取培养3 d后的切片,用SS2感染脑组织切片24 h,模拟SS2感染过程。荧光定量PCR检测感染后炎性细胞因子表达情况。感染后的脑组织切片模型中IFN-γ、iNOS和IL-10这3种炎性细胞因子变化显著,iNOS、IFN-γ的表达量与对照组相比明显上升,而IL-10的表达量则出现了一定下降,ELISA结果也显示了相似的结果(图1D),由此可以认为这3种炎性细胞因子与SS2感染猪脑组织有关。
2.2 IFN-γ能诱导BBB通透性增强为了评估IFN-γ在SS2感染后对BBB的影响,检测IFN-γ处理对BBB完整性的变化。首先分离仔猪原代BMEC和ACs,构建了PBMEC/AC共培养BBB模型。并在Transwell中培养仔猪原代BMECs和ACs,期间持续监测共培养BBB模型的TEER值,模型在4 d后TEER值达到稳定状态。统计分析显示,BMEC/AC共培养BBB模型的TEER值显著高于仅由PBMECs构建的BBB模型(图2A)。并通过HRP渗透性检测BBB模型的通透性(图2B),结果显示BMEC/AC共培养BBB模型的基底外侧HRP浓度明显低于由PBMEC构建的单一BBB模型(图2B),表明BMEC/AC共培养BBB模型的完整性更好。除此之外,还对模型中外排转运蛋白进行评估。BMEC/AC共培养BBB模型中外排转运蛋白在转录水平显著高于由PBMEC构建的单一BBB模型(图2C)。以上结果表明,成功建立了BMEC/AC共培养BBB模型。

图1 SS2感染对脑组织的影响 A.脑组织切片随培养时间的变化情况;B.PI染色检测切片活性;C.不同培养条件下脑切片LDH释放情况;D.SS2感染脑切片后RT-PCR和ELISA检测细胞因子释放情况。*.表示P<0.05;**.表示P<0.01。下同
用IFN-γ(40 μg/L)刺激成功构建的BBB共培养模型,与对照组相比,24 h后BBB共培养模型的TEER值显着降低(图2D),表明IFN-γ显著损伤BBB共培养模型,IFN-γ可能破坏BBB的完整性。
2.3 IFN-γ能诱导PBMEC发生自噬以40 μg/L 的IFN-γ刺激脑切片,在转录水平检测自噬标志分子LC3的表达。结果显示与对照组相比,IFN-γ处理组的LC3Ⅱ/LC3Ⅰ的比值明显升高(图3A),所以可以推测SS2刺激后通过IFN-γ介导脑组织在转录水平发生了自噬。同时LDH检测发现感染后脑切片的LDH释放量显著升高(图3B)。以上结果表明IFN-γ会诱导自噬进而引起脑组织损伤。
通过之前的试验结果发现IFN-γ会破坏BBB引起脑组织中LC3Ⅱ的高表达,为判断究竟是由哪种细胞引起的自噬,检测IFN-γ是否会引起仔猪原代BMEC发生自噬。在细胞培养基中加入40 μg/L IFN-γ,细胞内LDH释放量随刺激时间的增加而升高(图3C)。在刺激12或24 h后,Western blot检测,结果表明分离的仔猪原代BMECs可表达高水平的LC3Ⅱ(图3D)。此外,通过RT-PCR检测自噬信号通路中的几个关键因子,包括ULK1,Becline和LC3Ⅱ,均被上调(图3E)。该结果表明IFN-γ可诱导PBMEC自噬并破坏BBB的完整性。
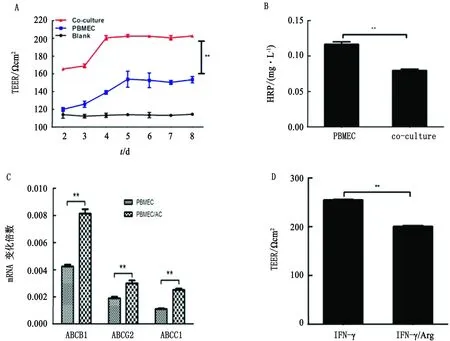
图2 BBB模型的建立及IFN-γ对BBB通透性的影响 A.BBB模型电阻值随培养时间变化情况;B.HRP渗透量评估BBB模型完整性;C.外排转运蛋白表达评估BBB模型完整性;D.IFN-γ引起BBB共培养模型电阻值显著下降
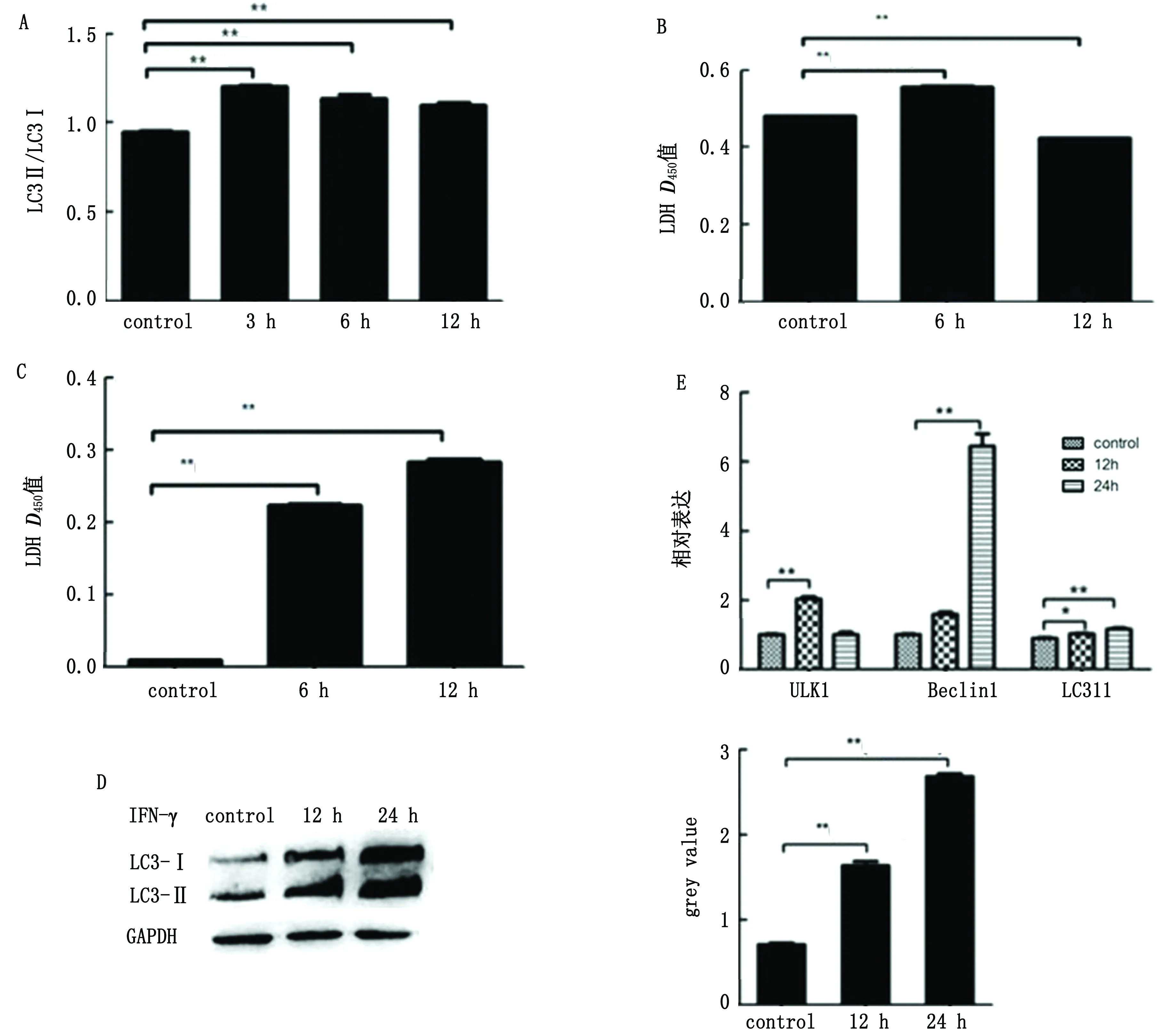
图3 IFN-γ诱导PBMEC发生自噬 A.IFN-γ引起脑切片中LC3高表达;B.IFN-γ引起脑切片中LDH释放量增加;C.IFN-γ诱导PBMEC释放LDH;D.IFN-γ可诱导PBMEC表达LC3;E.IFN-γ诱导自噬相关分子转录水平表达上调
2.4 精氨酸可以减轻IFN-γ诱导的自噬并保护BBB模型的完整性据报道,精氨酸的补充可以缓解由IFN-γ诱导的上皮细胞自噬,所以探究补充精氨酸是否可以减轻IFN-γ对PBMECs自噬的影响。与只添加IFN-γ相比,添加精氨酸(10 mmol/L)后 LC3Ⅱ/LC3Ⅰ值显著降低,表明补充精氨酸可以减轻由IFN-γ引起的PBMEC自噬(图4A)。通过检测电阻值以判断添加精氨酸 (10 mmol/L)治疗后对BBB完整性的影响。结果显示,与仅添加IFN-γ组相比,添加精氨酸治疗后能显著降低IFN-γ对BBB完整性的破坏(图4B)。

图4 精氨酸可减轻IFN-γ对PBMEC的损伤 A.精氨酸可以减轻由IFN-γ引起的PBMEC自噬;B.补充精氨酸可显著降低IFN-γ对BBB完整性的破坏
3 讨论
在本研究中,利用猪脑组织切片,以及体外构建PBMEC、AC共培养模型和猪原代BMEC分3个层次逐步证明在SS2感染下IFN-γ的表达会显著增加。并通过添加IFN-γ发现高水平的IFN-γ可显着破坏BMEC/AC共培养BBB模型的完整性,这说明SS2感染后可能通过激活自噬相关因子诱导PBMEC自噬,进而破坏BBB的完整性。添加精氨酸治疗后,发现可显著降低细胞自噬和IFN-γ诱导的BBB损伤,表明其可拮抗IFN-γ对猪脑部的损伤。
IFN-γ是一种具有化学活性的可溶性细胞因子,已被证实在免疫和病理反应中起多种作用。一方面,已确定IFN-γ在各种免疫细胞的功能和成熟中起着重要作用[9]。另一方面,IFN-γ可通过激活巨噬细胞而促进自噬体的成熟。越来越多的研究支持上皮细胞自噬的发展与IFN-γ密切相关[10]。LC3Ⅱ是哺乳动物自噬体膜蛋白的标志物,反映了自噬的变化[11]。本研究证实IFN-γ可促进PBMECs自噬的发展,自噬信号通路的关键分子ULK1[12]和 Beclin1[13]等均显着上调。因此,IFN-γ诱导PBMECs自噬的信号途径可能有更多的相关分子参与,此过程还需进一步研究。但本研究说明高水平的IFN-γ可以促进SS2脑膜炎的发展。
精氨酸可通过促进淋巴细胞生长、巨噬细胞活化以及细胞因子的分泌等方式参与机体的免疫调节。在小鼠饲喂过程中添加精氨酸可通过促进IFN-γ的分泌增强Th1免疫反应,同时可增加IL-10水平调节免疫作用。本研究中,补充精氨酸可显著拮抗IFN-γ对BMEC的破坏作用,抵御由IFN-γ引起的自噬对机体损伤,为精氨酸参与机体免疫调节提供了又一证据。
在研究中还发现SS2感染后iNOS的表达显着增加。iNOS作为同工酶,可以由内皮细胞、神经细胞和巨噬细胞产生。它可以帮助巨噬细胞抵抗免疫系统中的病原体[14]。因此,推测iNOS可能被脑组织释放以抵抗SS2的侵袭。IL-10在抑制巨噬细胞和树突状细胞功能中起重要作用,有助于控制和启动免疫反应[15]。IL-10的过量产生通常会导致无法控制微生物的感染。本研究发现SS2感染后IL-10的表达显着下降,而较低的IL-10可能有助于控制SS2感染,但该机制需要进一步验证。
本试验成功构建原代PBMEC/AC共培养BBB模型,为研究BBB功能提供了很好的模型,例如鉴定可通过BBB进入的细胞因子以及筛选BBB治疗剂。此外,SS2感染产生的高水平IFN-γ可以通过诱导PBMEC的自噬而严重破坏BBB的完整性,而精氨酸的补充可以对抗这种现象。精氨酸与IFN-γ之间呈负相关,可以作为诊断治疗SS2脑膜炎的综合指标,本研究为进一步研究SS2脑膜炎奠定了良好的基础。
猜你喜欢
云南化工(2021年9期)2021-12-21
宁夏医学杂志(2020年3期)2021-01-21
天然产物研究与开发(2018年4期)2018-05-07
中国音乐教育(2017年4期)2017-05-20
中国男科学杂志(2016年9期)2016-03-20
中外医疗(2015年11期)2016-01-04
中国当代医药(2015年30期)2015-03-01
中国医疗美容(2015年5期)2015-02-03
河南科技(2014年18期)2014-02-27
中国医学科学院学报(2013年6期)2013-03-11
