
宽体沙鳅脑组织学研究
2020-03-04 03:16史晋绒张青莲沙小宇王永明谢碧文
水生生物学报 2020年1期
史晋绒 张青莲 沙小宇 王永明 谢碧文
(内江师范学院生命科学学院, 长江上游鱼类资源保护与利用四川省重点实验室, 内江 641199)
鱼体器官和系统的正常生理、生化过程是在神经系统的主导下实现的[1]。作为脊椎动物中种类最多的类群, 不同鱼类对外界环境中声音、光线、气味、温度等变化都形成了较固定的条件反射[1]。由于鱼类缺乏大脑皮层, 脑的机能活动主要表现在脑各部分之间的联系上[1]。因此, 对鱼类神经系统的研究有利于分析鱼类对环境变化的适应性及中枢神经系统的相关功能。
宽体沙鳅(Siniotia reevesaeChang), 隶属鲤形目(Cypriniformes), 鳅科(Cobitidae), 沙鳅亚科(Botiinae), 沙鳅属(Sinibotia), 主要分布于长江干流、岷江、金沙江、雅砻江、沱江等水系的下游, 为长江上游特有及重要经济鱼类[2]。近年来由于水利工程建设、过度捕捞及环境污染等因素的影响, 其野生资源量急剧下降。目前, 关于宽体沙鳅的研究主要集中在胚胎发育[3,4]、繁殖生物学[5,6]、消化系统[7,8]、染色体核型分析与DNA含量测定[9]等方面, 尚无神经系统的相关研究。本文通过对宽体沙鳅脑组织结构进行连续观察, 了解其脑组织结构特征, 旨在完善该鱼的基础生物学资料, 丰富鱼类中枢神经系统组织学研究内容, 为脊椎动物比较解剖学和神经生物学的研究提供理论参考。
1 材料与方法
1.1 实验材料
2017年12月, 从长江上游鱼类资源保护与利用四川省重点实验室选取体质健壮、健康无伤的性成熟宽体沙鳅10尾, 体重(14.143±4.384) g, 体长(8.740±0.838) mm。实验鱼均在自然水温、自然光周期、鱼缸循环潜水泵冲水供氧条件下养殖, 养殖水体溶氧>5 mg/L以上, 氨氮<0.02 mg/L。
1.2 实验方法
新鲜活鱼经MS-222麻醉后断头取脑于10%福尔马林溶液中固定24h。将已固定的脑依次经梯度酒精脱水, 二甲苯透明及石蜡包埋后进行连续矢状和冠状切片(厚度5 μm), HE及Nissl染色, Nikon SM1500体式显微镜及Motic BA 400光学显微镜观察, Motic Images Plus 2.0显微图像分析软件测量,结果用平均数±标准差(Mean±SD)表示,n=30。
2 结果
宽体沙鳅脑分为5个部分: 端脑(Telencephalon)、间脑(Diencephalon)、中脑(Mesencephalon)、小脑(Cerebellum)和延脑(Myelencephalon), 呈中间宽两端细的梭形结构(图1)。
2.1 端脑
端脑位于脑最前端, 由前方的嗅囊(Rhinencephalon)、嗅叶(Lobi olfactorius)和后方的大脑(Cerebrum)组成。嗅叶呈半球状, 分化为嗅球(Olfactory bulbs)和嗅束(Olfactory tractus)。嗅球紧贴嗅囊, 和大脑之间通过嗅束相连接(图1)。大脑呈长柱状, 其结构复杂, 分区明显(图1)。
嗅球由外层的脑膜(Meninges)和中央区域的小细胞层(Small cell layer)组成(图版 Ⅰ、图版 Ⅱ-1)。脑膜由外层的上皮组织及内层结缔组织组成。外层上皮细胞为单层扁平上皮; 内层由外向内依次为神经纤维外层(Outer neural fiber layer)、小球层(The layer of glomerule)和神经纤维内层(Inner neural fiber layer), 三者分界不明显(图版 Ⅰ、图版Ⅱ-1)。神经纤维外层细胞较少, 呈椭圆形, 细胞小,染色较深, 呈斑块状。嗅神经末梢、血管和僧帽状细胞形成小球层。僧帽细胞(Mitral cell)多呈椭圆形, 长径(7.625±1.549) μm, 短径(5.572±1.003) μm,主要集中在小球层, 神经纤维外层和内层尚有分布(图版 Ⅲ-1)。神经纤维内层细胞分布较紧密, 细胞呈圆形, HE染色均匀, 颜色稍浅。嗅球中央区域的小细胞层中神经细胞多为圆形和椭圆形, 数量较多,分布紧密, 染色较深, 细胞直径(3.699±0.923) μm(图版 Ⅰ、图版 Ⅱ-1)。
嗅束又称为嗅茎(Olfactory stalk), 神经纤维排列稀疏, 有圆形小细胞散布其中, 直径(2.436±0.689) μm。近脑皮处有丰富血管围绕(图版Ⅰ、 图版Ⅱ-2)。嗅束呈中空的圆筒状, 其中心空腔为始于嗅球后部的嗅茎腔(Phinocoel, 图版Ⅰ, 图版Ⅱ-2)。
大脑背面中央有一纵沟称中央纵沟(Central longitudinal groove), 它将大脑分隔为左右对称的两个半球(图1a)。大脑半球的腹面有一纵行浅沟即内嗅沟(Internal olfactory sulcus, 图1b)。左右半球背面有一横行的浅沟, 横沟前面是额叶(Frontal lobe), 额叶后紧跟两纵行小叶, 距离中央纵沟近的称为背叶(Tergal lobe), 侧面的是侧叶(Lateral lobe),侧叶的后方是梨状叶(Pyriform lobe), 二者之间为梨状沟(Pyriform sulcus, 图1c)。侧叶和背叶之间有一呈Y形的明显小沟, 称为席氏沟(Schiff’s ditch), 它又叫Y状沟(图1a)。
在大脑横切面上, 包围大脑的薄而透明的膜状结构为不含神经组织的大脑皮, 属古脑皮(图版Ⅰ、图版Ⅱ-3)。大脑内致密结构为纹状体(Corpus striatum), 分为左右两基叶(图版Ⅰ、图版Ⅱ-3)。大脑中部为中央嗅区(Central olfactory area), 背叶、侧叶、梨状叶分别对应背嗅区(Dorsal olfactory area)、侧嗅区(Lateral olfactory area)和梨状区(Pyriform area)。各个分区周缘围绕大部分神经细胞, 分别形成对应的神经核。大脑两半球在前部相互分离, 在中后部腹面通过前连合(Commissura anterior)相联接, 前连合表现为横行的神经纤维(图版Ⅱ-4)。
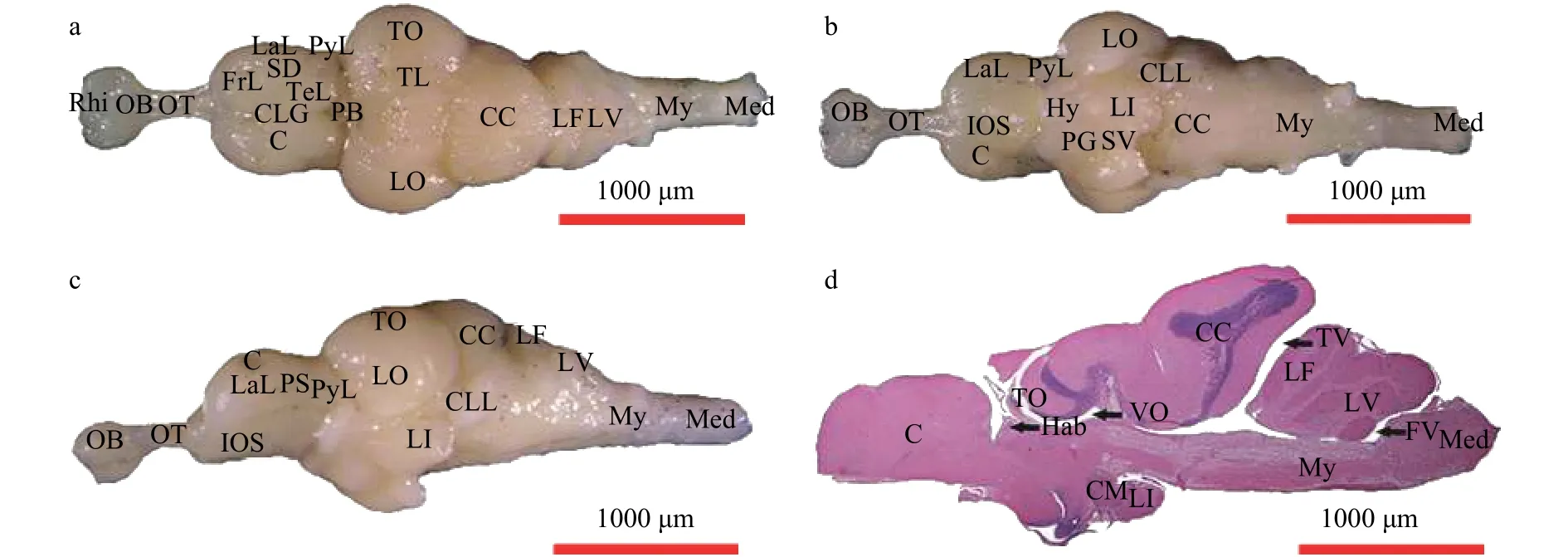
图1 宽体沙鳅脑背面观(a)、腹面观(b)、侧面观(c)及纵切图(d, HE染色)Fig. 1 Brain of the Sinibotia reevesae shown in dorsal view (a), venture view (b), lateral view (c) and vertical section (d, HE stain)
公共脑室中间两侧为连前核(Precommissure nucleus, 图版 Ⅰ、图版 Ⅱ-3), 该区域富含神经纤维, 细胞排列相对紧密。在大脑前中部, 背侧群(Dosal precommissure nucleus)细胞排列相对稀疏,细胞呈圆形, 直径(3.866±0.642) μm。腹侧群区域(Vetral precommissure nucleus)细胞排列较背侧群紧密, 直径(3.644±0.671) μm (图版 Ⅰ-3、图版 Ⅱ-3、图版 Ⅲ-2); 在大脑中后部背侧群、腹侧群逐渐缩小、排列更紧密, 显微镜下可对其进行明显进行区分: 背侧群细胞排列相对稀疏, 细胞直径(3.444±0.719) μm。腹侧群排列较背侧群紧密, 数量较背侧群多, 直径(3.201±0.806) μm, 腹侧群尼氏体含量多, Nissl染色较深(图版Ⅰ-4、图版 Ⅱ-4)。在大脑尾部仅见腹侧群细胞(图版 Ⅱ-5)。
Nissl染色可见大脑分区显著(图版 Ⅲ-2)。背嗅区细胞在大脑皮周边密布, 细胞小且圆, 直径(3.277±0.684) μm。侧嗅区细胞排列较背嗅区稀疏,呈颗粒状, 细胞大且圆, 直径(5.016±0.801) μm (图版 Ⅲ-2)。背嗅区有一神经核即背嗅核(The nucleus dorsal olfactory), 侧嗅区即为侧嗅核(The nucleus lateral olfactory, 图版 Ⅲ-2)。中央嗅区染色稍浅,被连前核、背嗅区、侧嗅区包围, 其神经细胞较大,呈圆形、椭圆形、多角形或不规则梭形, 大小(8.852±1.657) μm, 可以清晰地看到轴突, 神经细胞分布较其他区稀疏(图版 Ⅲ-2)。
大脑中后部, 前连合上方, 公共脑室腹部两侧为呈索状的视前核(Preoptic nucleus, 图版 Ⅱ-4、图版 Ⅲ-3)。视前核细胞列纵行, 纤维交错其中, 细胞多呈圆形, 直径(3.705±0.879) μm。
在大脑后部, 侧嗅核尾部出现呈梨状的梨状区,其神经纤维疏松, 内分布有梨状核(Pyriform nucleus, 图版 Ⅰ-4、图版 Ⅱ-5、图版 Ⅲ-4)。该区细胞呈圆形, 直径(1.837±0.586) μm, 后部常分布有索状排列的小细胞群(图版 Ⅲ-5)。梨状核始发处侧上方靠近侧嗅核方向的端脑基部, 有脚内核(Entopeduncular nucleus), 其细胞小且密, 呈圆形, 直径(2.751±0.425) μm(图版 Ⅱ-4、图版 Ⅲ-3)。梨状核腹侧, 近内嗅沟处带核(Tenae nucleus)圆形细胞大量聚集, 细胞直径(2.475±0.468) μm (图版Ⅱ-5、图版 Ⅲ-4)。
2.2 间脑
端脑后方逐渐延伸为结构复杂的间脑。间脑由上丘脑(Epithalamus)、背侧丘脑(Thalamus)及下丘脑(Hypothalamus)构成(图版 Ⅰ-5、图版 Ⅱ-6)。
上丘脑由背囊(Dorsal sac)、松果体(Pineal body)、松果缰(Habenula)、缰连合(Habenular commissure)及后连合(Posterior commissur)五部分组成,其中松果缰及缰连合为神经组织。松果体是在间脑与端脑相接处的间脑顶壁向前方突出的一囊状结构, 又称脑上腺(Epiphysis), 被背囊包裹, 属于内分泌腺(图版 I-5、图版 Ⅱ-6)。松果缰是一对位于上丘脑中央空隙侧壁顶部的长梭形隆起, 其内为缰核(Habenual nucleus, 图版 Ⅰ-5、图版 Ⅱ-6、图版Ⅲ-6)。缰核细胞小, 呈圆形或椭圆形, 细胞直径(2.448±0.606) μm, 密布神经纤维外周(图版 Ⅲ-7)。缰核的纤维不断延伸, 将两侧连接起来即缰连合(图版 Ⅰ-5、图版 Ⅱ-6、图版 Ⅲ-6)。缰连合上方出现横行的纤维即后连合, 其上方为中脑纵枕(Torus longitudinalis, 图版 Ⅰ-5、图版 Ⅱ-6)。后连合为中脑与间脑所共有, 其大部分纤维进入中脑, 可作为区分中脑与间脑的标准(图版 Ⅱ-6)。后连合尾部视叶腔内各有一个明显三角凸起, 称为副错位核(Paracommissual nucleus), 该区域细胞小、染色深且紧密围绕神经纤维外周(图版 Ⅰ-5、图版 Ⅱ-6)。
背侧丘脑又称丘脑, 分布在间脑横切面中部,细胞群散布于中央, 侧壁增厚形成视丘(Optic thalamus), 第3脑室(Third ventricle)位于中间(图版 Ⅰ-5、图版 Ⅱ-6)。第三脑室顶部有可产生脑脊液的脉络膜丛(Choroid plexus, 图版 Ⅲ-6)。
下丘脑位于间脑腹面, 可分为漏斗(Infundibulum)、垂体(Pituitary gland)、血管囊(Saccus vasculosus)和下叶(Inferior lobe, 图1b)。在间脑纵切面上, 可见漏斗体位于间脑腹面稍后方, 是垂体与下叶形成的空腔, 其脑壁向后方外突(图版 Ⅰ-6、图版 Ⅱ-7)。漏斗后方伸出单个血管囊(图版 Ⅰ-6、图版 Ⅱ-7、图版 Ⅲ-8)。血管囊背侧为视前隐窝(Recessus preopticus), 呈弯月状, 左右各一(图版 Ⅲ-8)。漏斗体下方为垂体(图1d、图版 Ⅰ-6)。下叶位于下丘脑中后部血管囊两侧, 左右各一, 其内有下叶腔(Ventricle of inferior lobes)(图版 Ⅰ-7、图版Ⅱ-8), 第三脑室扩张伸入其间。在间脑后部, 下叶腔逐渐扩展, 并伸入漏斗(图版 Ⅰ-8、图版 Ⅱ-9)。漏斗体腹侧, 视前隐窝两侧各有一个被下叶包裹的乳头体(Corpus mamillare), 其内有乳头体核(Mamillary nucleus, 图版 Ⅲ-8)。乳头体核内细胞小且密,与纤维交织排列。下丘脑前部为视神经(Optic nerve)形成的视神经交叉(Optic chiasma), 交叉后为视束(Ophthalmic tract)入间脑。
2.3 中脑
中脑位于间脑上方, 由一对圆球状体的视叶(Lobus opticus)构成(图1)。视叶由视盖(Tectum opticum)、被盖(Tegmentum)和视叶腔(Ventriculus opticus)组成。视叶腔极大, 其内有小脑瓣(Valvula cerebelli)伸入(图版 Ⅰ-7、图版 Ⅱ-8)。
中脑背部为视盖, 由外向内分为边缘层(Marginal stratum)、视神经层(Opticun stratum)、表面纤维层(Stratum fibrosum et griseum superficiale)、中央细胞层(Central griseum stratum)、中央纤维层(Stratum album central)和围脑室层(Stratum periventriculare, 图版 Ⅲ-9、10)。边缘层纤维较厚且紧密分布, 未见有细胞分布, 与视神经层间有明显分离, 有神经纤维伸入视神经层; 视神经层血管丰富,细胞多呈卵圆形且散布于排列较稀疏的神经纤维中, 可见来自中心及双侧视束的视神经纤维; 表面纤维层最厚, 含有丰富的来自视神经的前枝纤维,纤维排列较密, 有较大细胞散乱分布其中, 细胞呈圆形、椭圆形或不规则形; 中央细胞层与表面纤维层分界明显, 薄, 细胞分散排布, 呈圆形或椭圆形,神经纤维稀疏, 联络视盖; 中央纤维层相对较薄, 细胞少, 由传出神经纤维组成, 纤维排列紧密并环绕围脑室层, 联络围脑室层和中央细胞层; 围脑室层较薄, 分为围脑室神经细胞(Periventricular neurocyte)及视盖深层纤维(Nerve fiber of deep tectum opticum)两部分, 围脑室神经细胞, 又称结层(Ganglian layer), 其中有神经幼芽细胞(Nerve budlet cell)密布,胞质极少, 胞核呈圆形, 细胞直径(2.296±0.570) μm,视盖深层纤维, 又称室管膜与视盖深层纤维层, 其中室管膜细胞(Ependymal cell)联结成束, 纤维束粗大可见(图版 Ⅲ-10)。
视盖背面与小脑瓣相接处左右各有一长梭形的突起, 称为纵枕(图版 Ⅰ-5、图版 Ⅱ-6), 其内细胞多且密集, 细胞直径(1.686±0.393) μm, 染色深(图版 Ⅲ-11)。背面的纤维束清晰可见, 将两视盖联系起来(图版 Ⅰ-7、8)。
中脑腹部为被盖, 被盖向视叶腔方向有三个明显突起, 分别为半圆枕(Torus semicircularis)、中隆起(Eminentia medialis)及峡核(Isthmi nucleus, 图版Ⅰ-7、图版 Ⅱ-8)。半圆枕是被盖与视盖在中脑前部相接处形成的半圆状突起, 包于小脑瓣腹外侧(图版 Ⅰ-7、图版 Ⅱ-8)。中脑后部半圆枕不显, 融入延脑(图版 Ⅰ-9、图版 Ⅱ-10)。半圆枕中有瓣侧核(Lateralis valrulae nucleus), 可见圆形细胞沿半圆枕呈一明显弧形(图 Ⅰ-7、图版 Ⅱ-7)。在中脑腹部两被盖中线双侧, 有一突起称中隆起(图版 Ⅰ-7、8), 向后延伸突起渐渐消失。中隆起中扁豆核(Lentiformis nucleus)在前部细胞小且密集, 后部逐渐消失(图版 Ⅰ-7)。中隆起和半圆枕中间有一明显突起为峡核, 其内细胞小且密(图版 Ⅰ-7、图版 Ⅱ-8)。中脑峡核靠近中隆起处可见动眼神经核(Oculomotor nerve nucleus)(图版 Ⅱ-8), 动眼神经核向后为滑车神经核(Trochlear nucleus, 图版 Ⅱ-9), 下方为滑车神经(Trochlear nerve, 图版 Ⅰ-8)。此外, 在中脑后部两下叶背外侧有三叉神经(Trigeminal nerve), 其向后延展至视叶消失(图版 Ⅰ-9、图版Ⅱ-9、10)。
视盖与被盖之间的空腔为视叶腔, 又称中脑腔,联通三、四脑室。两视叶间有中纵沟(Mesosulcus,图版 II-8)。
2.4 小脑
宽体沙鳅小脑发达, 位于中脑后背方, 由小脑瓣、小脑体(Corpus cerebellum)、小脑侧叶(Lateral lobe of cerebellum)和第三脑室组成(图1)。前方小脑瓣进入中脑, 将视盖分为两部分(图版 Ⅰ-7、图版 Ⅱ-7)。
小脑体极发达, 其横切显示细胞分层明显, 由外向内可分为分子层(Molecular layer)、蒲氏细胞层(Purkinje cell layer)和颗粒层(Granular layer)三层,各层均有血管分布(图版 Ⅲ-12)。分子层细胞多呈圆形, 少且分布稀疏, 直径(3.144±0.636) μm, 染色稍浅, 纤维较多, 呈束状伸入颗粒层; 蒲氏细胞层最薄, 围绕颗粒层, 细胞多呈椭圆形或不规则形, 数量较少, 胞核清楚可见, 细胞长径(10.516±2.050) μm,短径(7.700±1.319) μm; 颗粒层圆形细胞染色深且密集分布, 直径(1.966±0.507) μm (图版 Ⅲ-12)。颗粒层向后延伸进入小脑侧叶, 形成颗粒隆起(Eminentia granularis, 图版 Ⅱ-11)。
小脑瓣被视盖包围, 形态学上不可见, 同样分为分子层、蒲氏细胞层及颗粒层。其中, 颗粒层向两侧扩展。小脑前部分子层在腹部向下形成一对突起称为小脑腹脊(Crista cerebelli), 覆盖在延脑上方, 二者之间形成一个空腔为第三脑室(图版 Ⅰ-10)。在小脑前方小脑侧叶始发处下方, 延脑两侧有面神经(Facial nerve)穿过(图版 Ⅰ-10)。
2.5 延脑
延脑是位于脑的最后, 后部通出枕骨大孔即为脊髓(Spinal cord), 两者无明显的分界(图1)。延脑主要由面叶(Lobus facialis)、迷叶(Lobus vagus)、延脑本体和第四脑室(Fourth ventricle)组成, 其前部被小脑体略为遮盖(图1d)。
面叶、迷叶均位于延脑背面(图版 Ⅰ-11、图版 Ⅱ-12)。前方为面叶, 其纤维与细胞交错, 边缘细胞多且密, 中间细胞交织在神经纤维中, 细胞多呈圆形, 直径(2.697±0.846) μm (图版 Ⅰ-11、图版Ⅱ-12)。迷叶紧跟面叶后方, 较面叶发达, 通过前端斜切入面叶, 细胞直径(2.566±0.838) μm, 其形态、大小、分布与面叶相似(图版 Ⅰ-11、图版 Ⅱ-12)。面叶和迷叶交界处形成菱形窝(Rhomboid fossa)。第四脑室将面叶、迷叶和延脑本体分隔, 其后与脊髓的中心管相连(图版 Ⅰ-11、12)。在延脑本体中线两侧靠近第四脑室有迷走神经核(Vagal motor nerve nucleus, 图版 Ⅱ-12)。
3 讨论
3.1 宽体沙鳅脑组织结构特征
神经系统是生物体中最重要的机能调节系统,也是动物进化的重心。作为脊椎动物中种类最多的类群, 不同鱼类脑的外部形态及内部结构因生活方式不同略有差异[1]。
宽体沙鳅脑包括端脑、间脑、中脑、小脑和延脑, 外部形态符合真骨鱼类脑的结构组成[10—34],内部结构表现出一定差异性, 具体表现为: ①嗅束细且短, 属于典型的“鲤型”嗅叶[13,15]。嗅叶分层与贝氏高原鳅(Triplophysa bleekeri)[10]一致, 但僧帽细胞分布与之不同。②宽体沙鳅大脑连前核分群与贝氏高原鳅[10]相同, 但连前核细胞分布及形态在大脑前后部表现出一定的差异性。大脑具明显的脚内核。③宽体沙鳅视前核结构未见贝氏高原鳅[10]中分区明显的视前核大、小细胞。④宽体沙鳅间脑脑上腺不发达, 符合楼允东[3]对硬骨鱼类脑上腺的描述。背侧丘脑与下丘脑分界不明显, 松果缰、缰连和、血管囊清晰可见。⑤宽体沙鳅间脑中下叶发达, 一直延伸至中脑消失, 推测可能与间脑的综合交换作用相关[22]。⑥宽体沙鳅中脑较贝氏高原鳅[10]不发达, 但视盖组成结构与之相似。视盖边缘层与视神经层之间区分明显, 二者之间有一层排列稀疏的神经纤维层, 这一结构未在其他鱼类中发现[10—34]。⑦宽体沙鳅小脑体极发达, 蒲氏细胞层有许多不规则形细胞, 与贝氏高原鳅[10]不同。⑧延脑分化出发达的面叶和迷叶, 与贝氏高原鳅[10]结构类似。面叶位于延脑背侧前方, 表面有浅沟, 后方为迷叶, 与Evans[26]观点相符。

图版 Ⅰ 宽体沙鳅脑组织学图谱(HE染色)Plate Ⅰ The atlas of the brain of Sinibotia reevesae (HE-stained)

图版 Ⅱ 宽体沙鳅脑组织学图谱(Nissl染色)Plate Ⅱ The atlas of the brain of Sinibotia reevesae (Nissl-stained)

图版 Ⅲ 宽体沙鳅脑特殊结构观察Plate Ⅲ Histology of Sinibotia reevesae brain
3.2 宽体沙鳅脑结构与生活习性的相关性
为适应不同的生态习性, 鱼脑的不同部位发展出现一定的分歧。因此, 依据脑各结构的形态及发达程度可对各种鱼的生态习性进行初步推断。相关研究表明, 营底层生活的鱼类多数视盖退化, 面叶和迷叶发达, 表现为视觉退化, 触觉与味觉灵敏[10,13,14,22,32]; 营上层生活的鱼类视盖发达, 视觉接受区广, 延脑不分化[19,25,32]。且在生活习性相同的鱼中, 脑的形态结构越相似, 亲缘关系越近[15]。
宽体沙鳅脑僧帽细胞多且分布广, 可能与嗅觉发达有关[14]; 血管囊清晰可见, 推测其在垂直方向上的运动能力较强且能接受较大水压[22]; 视前核不分区且中脑不发达, 可能与宽体沙鳅眼小, 视觉略有退化有关; 小脑体发达, 表明具有较强的保持鱼体平衡及整合运动的能力[35]; 延脑分化出发达的面叶和迷叶与底栖生活有关[32]。综合脑各部分结构特点, 表明宽体沙鳅营底层生活, 主要以触觉、嗅觉和味觉来感知食物, 同时具备高的运动能力。由于目前对真骨鱼类脑各结构细胞的大小、数量、类型等的研究甚少, 鱼类脑组织结构是否与其生活习性表现出种属的特殊性还有待开展深入研究。
猜你喜欢
中国医药导报(2022年28期)2022-11-25
西北植物学报(2022年4期)2022-05-24
科学大众·教师版(2022年6期)2022-05-23
北京航空航天大学学报(2021年9期)2021-11-02
创新作文(小学版)(2019年4期)2019-07-24
哲思2.0(2017年12期)2017-03-13
西南军医(2016年2期)2016-01-23
中国医疗器械杂志(2015年5期)2015-12-31
医学研究杂志(2015年6期)2015-07-01
西南军医(2015年5期)2015-01-23
