
克氏原螯虾两种养殖模式的食物网结构及其食性比较
2020-03-04 03:16米武娟许元钊宋庆洋毕永红
水生生物学报 2020年1期
周 正 米武娟 许元钊 宋庆洋 毕永红
(1. 中国科学院水生生物研究所淡水生态与生物技术国家重点实验室, 武汉 430072; 2. 中国科学院大学, 北京 100049)
克氏原螯虾(Procambarus clarkii)原产于北美洲, 于20世纪30年代传入中国, 现已成为我国淡水甲壳类养殖第一品种; 2017年全国总产值约2685亿元, 比2016年增长83.15%, 目前的养殖模式主要有稻虾综合种养、池塘主养、河蟹池混养、莲藕田套养等[1]。稻虾综合种养为各地的主要养殖模式,可细分为稻虾共作、稻虾互作+共作等模式。稻虾共作模式(以下简称共作模式)是一种新兴的农业生产模式, 将种植业和养殖业有机结合起来, 每年8—9月中稻收割前投放亲虾, 翌年4—5月份收获成虾, 6月插秧, 亲虾与水稻一起生长, 水稻收割后亲虾在水稻田进行原位繁育, 如此循环轮替。稻田中天然饵料极为丰富, 还能提供虾的生长繁殖场所[2,3];而克氏原螯虾为杂食性动物, 高等水生植物、丝状藻类、底栖动物及腐殖质等均是其食物[4,5], 共作模式使稻田资源可得到充分利用。除了共作模式,池塘单养克氏原螯虾(以下简称精养模式)也是一种重要的养殖模式, 精养模式是指将池塘改造后进行克氏原螯虾养殖, 这种模式中物种单一、生态系统结构简单, 以大量饵料投喂获得克氏原螯虾的高产。精养模式一般亩产量为150—200 kg, 而共作模式相对较少, 为100—150 kg[1]。
尽管目前对克氏原螯虾养殖方面的研究很多,但关于其食性的专门研究却报道不多, 有文章通过胃含物分析对克氏原螯虾的食性进行了研究[4,5,7],也有通过稳定同位素方法对克氏原螯虾在稻田系统中的自然资源及投喂饵料利用情况进行了分析[5],暂无对两种养殖模式下克氏原螯虾食源情况的比较研究。本文通过稳定同位素技术, 比较共作模式和精养模式两种养殖模式下食物网结构, 并对两种模式虾克氏原螯虾对主要食源的利用情况进行计算, 确认了两种模式下食物网结构、物质传递损耗情况以及克氏原螯虾的食性差异, 为稻田物质能量的高效利用以及探索绿色健康的克氏原螯虾养殖模式提供理论依据。
1 材料与方法
1.1 地点与时间
所有样品于2018年3月、4月和5月在江汉平原的养殖水体采集, 该区域属于亚热带季风性湿润气候。精养模式样品采集地S1在湖北省洪湖市赤林口村(29°57′1.22″N, 113°34′34″E), 共作模式样品采集位S2在湖北省潜江市瞄新村(30°15′10.40″N,112°46′44″E)。两地相距不足100 km, 日照、温度、土壤、水环境等环境条件一致; 当地的农业种植水平和工艺不存在区域差异。两种养殖模式的单池养殖水体面积均为0.13 hm2, 在2月25日—3月1日之间投放虾苗, 精养模式下克氏原螯虾的放养密度(0.9—1.2)×105尾/hm2, 共作模式为5×104尾/hm2左右。每种模式选择两个养殖水体, 每个水体设置三处采样位点, 采集水体的POM、寡毛类、浮游动物等样品。其中精养模式养殖水体深约1.5 m, 投喂人工颗粒饲料为主, 日投饵量约为虾现存总重量的1%—4%, 投喂时全池均匀泼洒; 共作模式养殖池的中间是稻田部分, 水深约0.5 m, 四面环沟是克氏原螯虾的主要养殖水域, 环沟水深约1.5 m, 水稻的种植密度为1.8×105株/hm2, 除了稻田中的自然资源外, 辅助投喂人工颗粒饲料, 日投饵量为存虾总重量的1%—2%。
1.2 样品采集与处理
采集到的样品有: 克氏原螯虾、浮游动物、寡毛类底栖动物(以下简称寡毛类)、悬浮有机颗粒物(POM)、沉积颗粒有机物(SOM)、水生植物(金鱼藻Ceratophyllum demersum、伊乐藻Elodeanuttallii等)、湿生植物(水竹叶Murdannia triquetra)等。克氏原螯虾、POM、浮游动物、寡毛类样品均是在5月份采集, 但因为当温度升高后不一定能够采集到水绵、水生植物等样品, 所以部分样品在3—4月采集。
具体采样方法如下: 克氏原螯虾通过设置地笼捕获, 每种模式采集20只, 记录体长、体重, 体长是指眼柄基部至尾节末端的直线长度, 体重是指用滤纸吸去体表水分之后的湿重, 记录后取腹部肌肉,烘干待测; 浮游动物样品采用浮游生物网进行拖拽, 所得浮游生物用150 μm筛网过滤, 再用蒸馏水清养3h后, 过滤至450℃预烧2h的GF/C滤膜上; 寡毛类采用60目的抄网筛底泥后转移至60目的筛网, 用清水冲洗并挑选寡毛类, 放入清水中清养3h后, 过滤至预烧的GF/C滤膜上; POM是取500 mL水样, 过滤至预烧的GF/C滤膜上, 经过1 mol/L盐酸酸化处理, 再用蒸馏水冲洗去除盐酸; SOM是通过采集底泥后, 缓慢放入蒸馏水中, 待其沉入底部后, 轻轻搅动上层水, 这时会有部分颗粒物悬浮, 收集上层水,过滤至预烧的GF/C滤膜, 经1 mol/L盐酸酸化处理,再用蒸馏水冲洗去除盐酸; 水生植物、水绵、湿生植物等样品, 经过徒手采集, 用蒸馏水冲去表面杂物; 福寿螺、中华圆田螺等样品采集后, 清养24h,去壳取其肌肉组织。以上所有样品在采集、初步处理后通过车载冰箱(-20℃)运回实验室进行处理,之后60℃恒温烘干至恒重(24—36h), 放入干燥器保存等待稳定同位素分析。
1.3 稳定同位素分析
采用稳定同位素质谱仪(Isotope Ratio Mass Spectrometer, Delta V Advantage)进行样品的稳定性同位素δ13C和δ15N测定。样品烘干碾磨至粉末状后,采用锡杯(Tin capsules for solid, TLS24005300)包被样品进行上样, 其中动物样品每份称取300—500 μg,POM、DOM样品每份称取1000 μg左右, 植物样品称取700 μg左右。C、N稳定同位素的参考标准分别为: 美国拟箭石(PDB)、空气中的氮气(N2)。因在自然条件中, 稳定同位素13C和15N含量很低, 所以其测定结果以δ13C和δ15N形式表示, 计算公式如下:

其中X为13C或15N,Rsample为样本的同位素比值,Rstandard为参考标准样品的同位素比值。每测28个样品之后再测两个已知稳定同位素值的标样, 通过测量的标样的值来测试仪器的精准性, 两个标样型号分别为USGS40和USGS41。
1.4 数据处理与统计分析
通过Excel对原始数据进行处理, 计算平均值、标准差等; 使用OriginPro2017、R3.5.1进行作图。
食物链长度计算方法如下: 食物链长度=(最高营养级的δ15N-基准生物的δ15N)/3.4‰+a, 此处的3.4‰即为富集系数, a是在计算该生态系统消费者营养级时设的基线生物的营养级。为了避免误差出现, 此处的营养级计算不采用POM作为基线计算, 而采用食性更为稳定、国内外更为普遍应用的螺类作为基线, 因为福寿螺为初级消费者, 将其营养级定为2[6,8]。
通过基于R语言的稳定同位素混合模型(SIAR)对同位素数据进行食源分析[9], 分馏系数采用文献中甲壳类的分馏系数, C13为1.3‰, N15为3.9‰[10]。将δ13C、δ15N及其营养富集因子数据输入R语言的SIAR程序包, 拟合得出各食源对克氏原螯虾的贡献率。通过基于R程序的贝叶斯算法(SIBER)对同位素数据进行计算, 并将这些结果转换成一些指标, 使其能够反映生物生态位情况和群落特征[11,12]。选择两种模式中均存在的五种消费者(克氏原螯虾、浮游动物、寡毛类、福寿螺Pomacea canaliculata、中华圆田螺Cipangopaludina cahayensis)进行生物群落特征参数计算, 群落特征共6种指标, 分别为: 氮值范围NR(δ15N range)、碳值范围CR (δ13C range)、总面积TA (Total area)、平均离心距离CD(Mean distance to centroid)、平均最邻近距离NND(Mean nearest neighbor distance)、最邻近距离标准差SDNND(Standard deviation of nearest neighbor distance)。所有计算基于δ13C-δ15N双坐标图, 其中CR和NR通过该水域中13C、15N富集程度最高的消费者的δ13C值和δ15N值减去富集程度最低的消费者的δ13C值和δ15N值得到, 可分别反映碳源范围和营养长度; TA是δ13C-δ15N值双坐标图中各水域消费者围成凸多边形面积, 可反应生物对生态空间的利用情况; CD是同一水域中各消费者距其中心点(即该处水域消费者的δ13C、δ15N平均值)距离的平均值; MNND通过计算每种消费者和距离其最近的消费者距离的平均值得到, 可反应物种聚集密度情况; SDNND通过计算每种消费者和距离其最近的消费者距离的标准差得到, 可反应物种聚集均匀度情况[12]。在计算得到的六个生物群落指标中, 前四个指标(NR、CR、TA、CD)反映物种空间范围, 代表群落营养结构多样性, 后两个指标(NND、SDNND)反映物种相对位置, 代表营养冗余[11]。除了群落指标之外, 另外对这五种消费者的生态空间利用情况分别进行计算, 考虑到TA可能因个别数据产生较大偏差, 使用贝叶斯标准椭圆面积(SEA)可以减少样本误差, 对标准椭圆校正后的面积为SEAC, 三者均可反应物种的生态空间利用情况[13]。
通过SPSS软件进行克氏原螯虾体长、体重与δ13C、δ15N的相关性分析, 如果存在显著相关性, 则进一步对两者进行线性回归分析。
2 结果
2.1 碳、氮稳定同位素值与营养级
由表1可知, 精养模式中水生植物、湿生植物、POM等基础食源的δ13C值介于-29.42‰—-13.01‰,δ15N值介于-6.31‰—2.72‰, 消费者的δ13C值介于-31.76‰— -26.38‰,δ15N值介于2.33‰—8.05‰; 轮作模式中基础食源的δ13C值介于-32.97‰— -12.5‰,δ15N值介于-5.16‰—0.88‰,消费者的δ13C值介于-34.22‰— -25.34‰,δ15N值介于2.72‰—5.97‰。
计算各消费者营养级的富集系数采用在淡水生态系统中普遍应用的数值3.4‰, 采用福寿螺作为评价其他消费者营养级的基准线。经计算得到在精养模式的生态系统中各消费者的营养级分别为:克氏原螯虾2.34、浮游动物1.87、泽蛙3.46、蝌蚪2.90、麦穗鱼2.65、寡毛类1.77、中华圆田螺1.87、福寿螺2.00。在精养模式系统食物网中,蛙、蝌蚪处于较高营养级, 克氏原螯虾、麦穗鱼处于中间营养级, 寡毛类和浮游动物处于较低营养级;在共作模式的生态系统中各消费者的营养级分别为: 克氏原螯虾1.79、浮游动物1.44、寡毛类1.49、沼虾1.45、泥鳅2.39、中华圆田螺1.47、福寿螺2.00。在共作模式系统食物网中, 泥鳅处于较高营养级, 克氏原螯虾、福寿螺处于中间营养级,寡毛类、沼虾、浮游动物等处于较低营养级。
生态系统的食物链长度取决于该系统中营养级最高的消费者。经计算得到精养模式生态系统的食物链长度为3.46, 共作模式生态系统的食物链长度为2.39。
2.2 食物网结构
选择两种模式中共同存在的五种类群(虾、浮游动物、寡毛类、福寿螺、中华圆田螺)进行生物群落指标计算(图1)。在计算得到的六个生物群落指标中(表2), 两种模式的NR差别不大, 说明两生态系统营养级长度相似; 共作模式的CR大于精养模式, 说明其生态位多样化程度更高; 共作模式的CD大于精养模式, 说明其平均营养级多样性较低;共作模式的MNND和SDNND均大于精养模式, 说明其营养冗余程度更低。
由表3和图2可知, 克氏原螯虾的TA、SEA、SEAC接近, 说明两种模式下克氏原螯虾的生态空间利用率差别不大; 寡毛类、中华圆田螺在精养模式中的TA、SEA、SEAC大于共作模式, 说明寡毛类、中华圆田螺在精养模式中的生态空间利用率大于共作模式; 浮游动物和福寿螺在精养模式中的TA、SEA、SEAC小于共作模式, 说明浮游动物和福寿螺在精养模式中的生态空间利用率小于共作模式。将几种生物的生态空间利用率进行对比, 虾和寡毛类对生态空间利用率较大, 其他生物的生态空间利用率较小。另外由图3可以看出5种消费者的生态空间重叠不明显。
2.3 克氏原螯虾食性分析
由表4可知, 精养模式和共作模式中,δ13C值和体长体重均无相关性(P>0.05), 但δ15N值和体长体重均有相关性(P<0.05), 克氏原螯虾的体长、体重均和其δ15N存在线性关系, 体内的15N富集程度随体长、体重增加而逐渐增大(图4)。
在两种养殖模式下克氏原螯虾各饵料贡献率是通过SIAR(Stable isotope analysis in R)软件包在R软件上计算得出。从SIAR计算结果可知, 各种饵料在精养系统中的平均贡献率为: 水绵为3.94%、寡毛类为14.03%、浮游动物为30.49%、饲料为16.76%、POM为13.67%、湿生植物为2.03%、SOM为12.08%、水生植物为7.00%; 在共作模式中为: 水绵为11.17%、寡毛类为12.07%、浮游动物为12.16%、稻秆为10.75%、POM为7.61%、湿生植物为10.86%、SOM为11.32%、水生植物为12.31%、饲料为11.72%(图5)。
3 讨论
3.1 碳、氮稳定同位素值及营养级
在两种模式生态系统中, POM、SOM、水绵等基础食源的δ15N值均较小, 其中两系统POM、水绵的δ15N出现了负值, 可能是试验田或附近施用化学肥料导致, 化肥中的δ15N值一般较低, 会影响这些基础食源的δ15N值[14]。在精养模式中, 两种水生植物(金鱼藻和伊乐藻)的δ13C和δ15N值相差不大, 在共作模式中也类似, 光合作用方式和所处环境的类似性决定了其δ13C和δ15N接近[15,16], 另外两者在生理生态上都不存在太大差异, 且对于克氏原螯虾的适口性也相似, 所以我们在之后的食源分析中, 将同一生态系统中的不同水生植物合并为一个食源计算。两系统的水竹叶虽然所处环境不同, 但δ13C接近, 这是因为植物的稳定同位素值虽然受到环境影响, 但更大程度上是由其光合作用方式决定的, 两系统的水竹叶均为C4植物, 故其δ13C接近, 同理, 两系统的水生植物δ13C也较为接近[15,16]。δ13C反应消费者的食源[17], 在精养模式生态系统中,消费者的δ13C值范围为-31.74‰(浮游动物)—-25.68‰(泽蛙), 相差6.06‰; 共作模式生态系统中,消费者的δ13C值范围为-34.22‰(浮游动物)—-25.34‰(克氏原螯虾), 相差8.88‰。共作模式消费者的δ13C范围大于精养模式的, 说明共作模式生态系统的碳源范围更广, 这符合共作模式生态系统较为复杂, 具有更丰富生物多样性的特征[18—22]。
计算消费者营养级时所采用的富集系数一般为3.4‰[7], 国内外常将螺类作为基线进行营养级计算[6], 本研究的两系统中福寿螺较为常见, 生物量也远大于中华圆田螺, 所以将福寿螺作为初级消费者,并将其营养级定为2[8]。经计算得到, 在精养模式生态系统中各消费者的相对营养位置关系为: 泽蛙占据最高营养级, 克氏原螯虾、麦穗鱼处于中间营养级, 寡毛类和浮游动物处于较低营养级。泽蛙占据最高的营养级, 与其以动物性食物为主有关[23]; 克氏原螯虾和麦穗鱼都为杂食性消费者[7,24], 两者营养级处于中间; 而寡毛类和浮游动物的食性分别为沉积食性、滤食性[18,19], 两者处于营养级底层。在共作模式的生态系统中各消费者的相对营养位置关系为: 泥鳅占据最高营养级, 克氏原螯虾、福寿螺因其杂食性处于中间营养级, 寡毛类、沼虾、浮游动物则处于较低营养级。
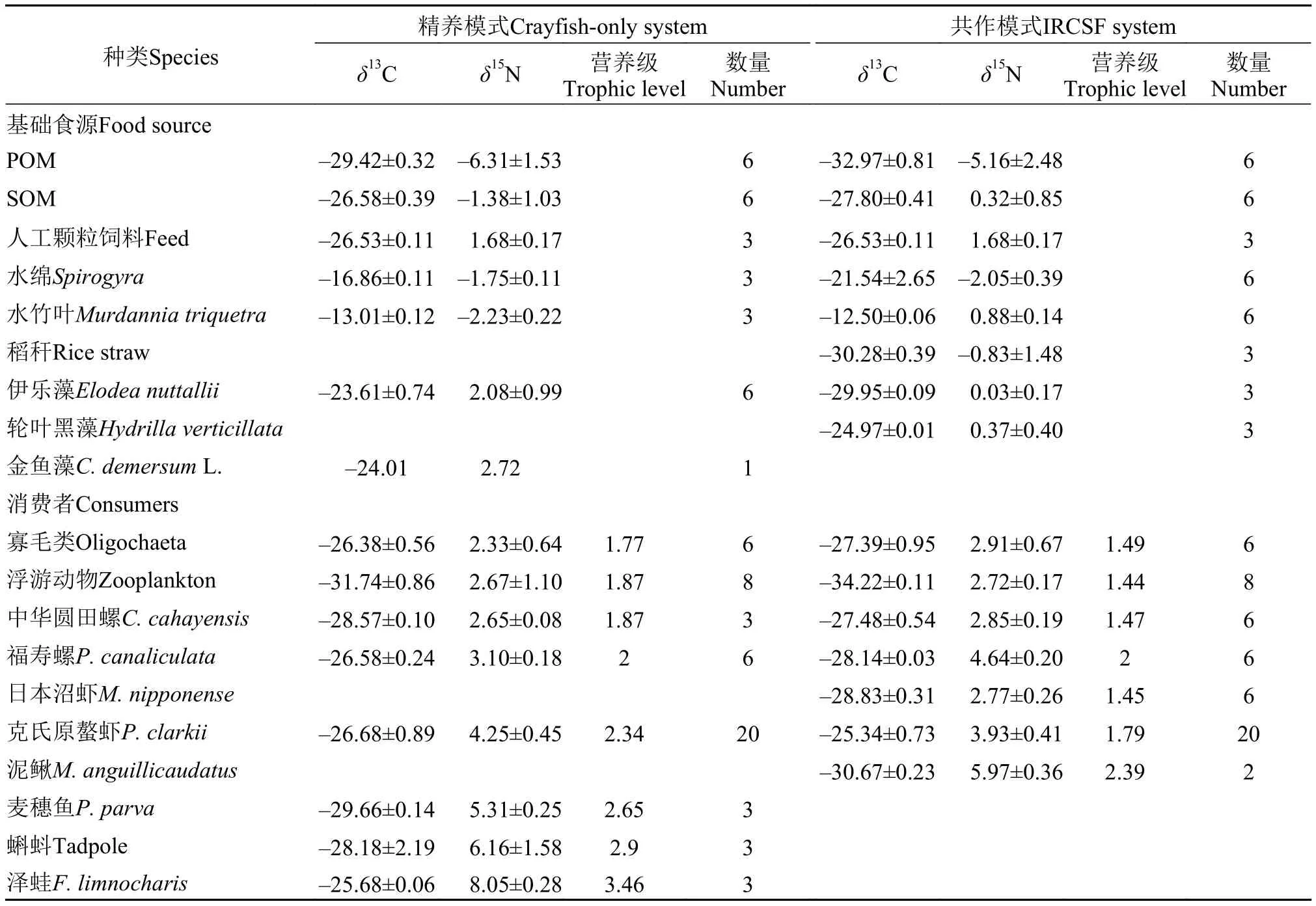
表1 两种养殖模式不同物种的碳氮稳定同位素值及营养级Tab. 1 Value of δ13C and δ15N for different species in two aquaculture systems
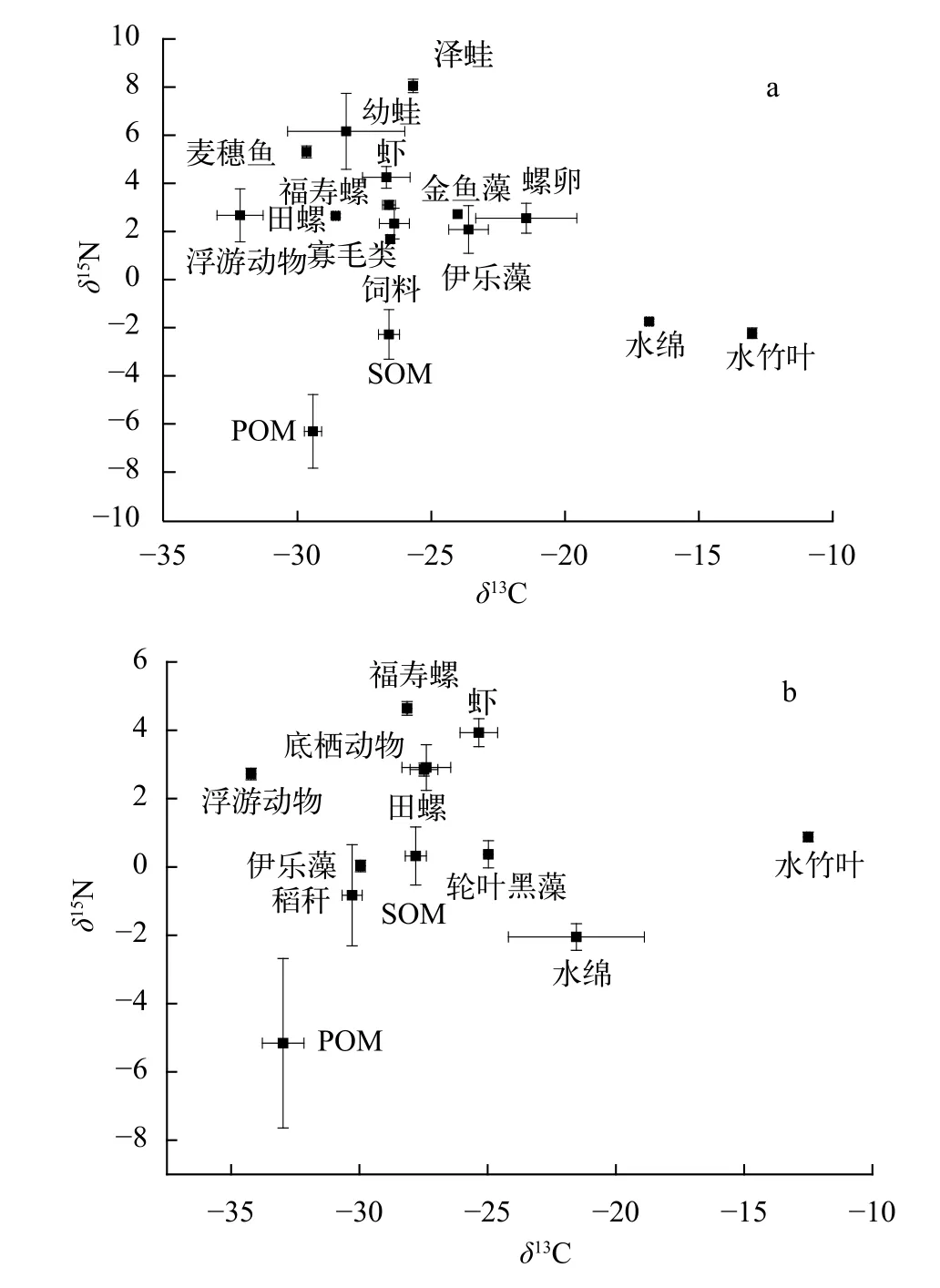
图1 两种养殖模式的食物网结构(a. 精养模式; b. 共作模式)Fig. 1 The food web of two systems (a. crayfish-only system;b. IRCSF system)

表2 两种模式的稳定同位素生物群落指标Tab. 2 The Layman’s community-wide metrics in two systems
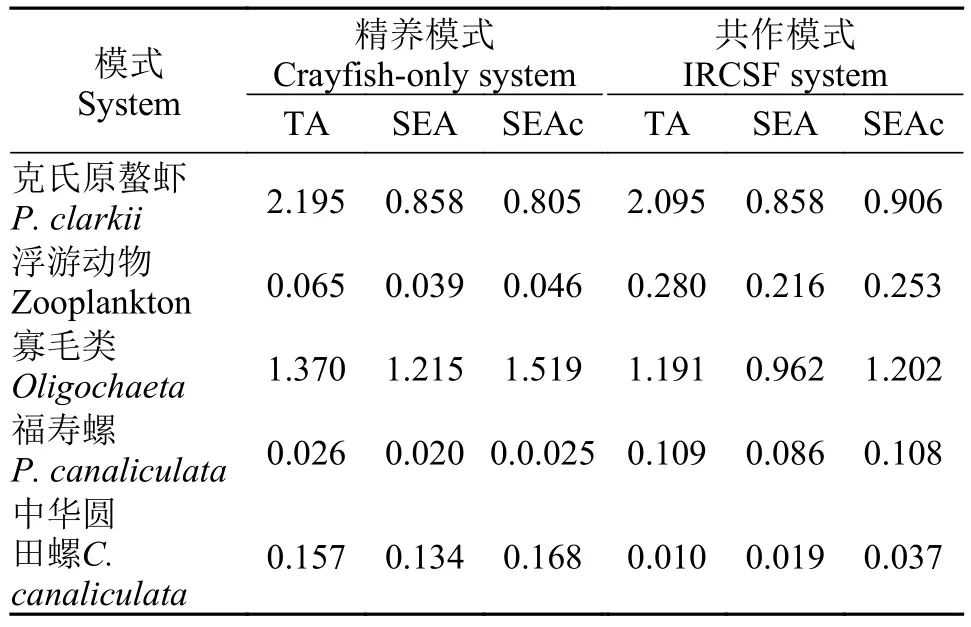
表3 两种模式下各生物的生态空间利用情况Tab. 3 Isotope niches/resource use areas of consumers in two systems
两种模式相对比, 在精养模式下克氏原螯虾的营养级(2.34)高于共作模式(1.79), 可能与精养模式下克氏原螯虾对于动物性饵料摄取更多有关(图5)。根据食物链长度的能量假说(Energetic hypothesis),食物链越长, 能量在食物链的传递过程中损耗就越多, 也就是说对于主要目的是产出克氏原螯虾成虾的养殖系统来说, 精养模式中能量从基础食源传递至克氏原螯虾的食物链传递损耗更高。但是根据统计, 精养模式一般亩产克氏原螯虾在150—200 kg,而共作模式为100—150 kg[1], 造成这种结果的原因可能是精养模式中饲料的添加导致其生态系统中的基础食源对于克氏原螯虾足够充足, 虽然在能量传递过程中的损耗大于共作模式, 但是亩产量高于共作模式。
3.2 食物网结构和生态空间利用情况

图2 两种养殖模式中虾类的贝叶斯标准椭圆Fig. 2 Standard ellipse area (SEA) of crayfishes in two different systems

图3 两种养殖模式中消费者的贝叶斯标准椭圆Fig. 3 Standard ellipse area of consumers in two systems

表4 克氏原螯虾稳定同位素值和体长体重的Pearson相关性系数Tab. 4 Pearson’s correlation coefficient between stable isotope and body length/weight of P. clarkia
两种模式的稳定同位素生物群落指标中(表2),NR值可以反映食物网营养结构多样性的垂直结构,两种模式的NR差别不大, 说明两生态系统营养级长度相似; CR值反映了食物网中初始食源的多样性特征, 共作模式的CR大于精养模式, 说明共作模式中各消费者的初始食源范围更广[20]; TA和CD分别反映了食物网营养结构多样性的总程度和平均程度, 共作模式的TA和CD均大于精养模式, 表明其食物网营养结构的多样性总程度和均匀程度均大于精养模式; MNND和SDNND反映了生态系统的营养冗余程度, 这两个值越小, 生态系统的营养冗余程度越高, 共作模式的MNND和SDNND均大于精养模式, 说明与精养模式相比, 共作模式生物群落中不同物种的营养生态位宽幅较大, 营养结构多样性较高, 群落的营养生态位范围较宽, 生态系统的营养冗余程度较低[12,21]。这可能是因为共作模式种植了水稻, 导致其物种的周围生态环境相对于精养模式更为复杂, 增加了消费者的食物来源途径,从而拥有更高的CR值, 另外基于不同物种间的相互作用, 具有更丰富的营养结构多样性[20,22], 因此其反应营养结构情况的TA和CD值也较高; 精养模式的营养冗余程度较高, 可能是因为其生态系统中含有较多的人工饲料, 导致不同消费者的基础食物来源较为集中, 人工饲料通过不同的途径进入不同的消费者中, 从而导致不同消费者的营养生态位重叠明显, 其生态系统的营养冗余程度也较高, 具体反映在较高的MNND和SDNND值上[12]。就食物网的营养结构而言, 共作模式具有较广基础食源(CR),更为丰富的营养结构多样性(TA、CD), 较低的营养冗余(NND、SDNND), 其消费者具有更多的基础食源和生态空间可以选择, 所以不同物种间的竞争程度相对于精养模式来说更小。

图4 精养模式(三角形)、共作模式(圆形)克氏原螯虾δ15N值和体长(a)/体重(b)关系Fig. 4 Relationships of δ15N values to body length (a)/weight (b)of P. clarkia in crayfish-only systems (triangle) and IRCSF system(circle)
在反应生态空间利用率的指标中(表3), 两种模式下克氏原螯虾的TA、SEA、SEAC差别都很小, 说明两种模式下其生态空间利用率十分接近。虽然共作模式中食物网的食源多样性高于精养模式, 但是仅克氏原螯虾来说, 其在两种不同模式中对于生态空间的利用率差别很小。造成这种现象的原因可能是克氏原螯虾属杂食性动物, 两种养殖模式提供的基础食源都足够充足, 生长环境也相对适宜。在这种情况下, 克氏原螯虾会优先选择适合自己的饵料和生存空间, 所以造成了虽然共作模式中食物网的食源多样性、食物网营养结构多样性都高于精养模式, 但两者克氏原螯虾的生态空间利用率相差不大的现象。
将同种模式下不同生物的生态空间利用率进行对比, 可以发现克氏原螯虾的生态空间利用率相对较大, 可能是因为克氏原螯虾的杂食性较为明显,其能够利用的食源相比其他消费者更为广泛; 浮游动物、福寿螺等的生态空间利用率相对较小, 这和它们食性较为单一的特性相符合。
3.3 克氏原螯虾食性分析
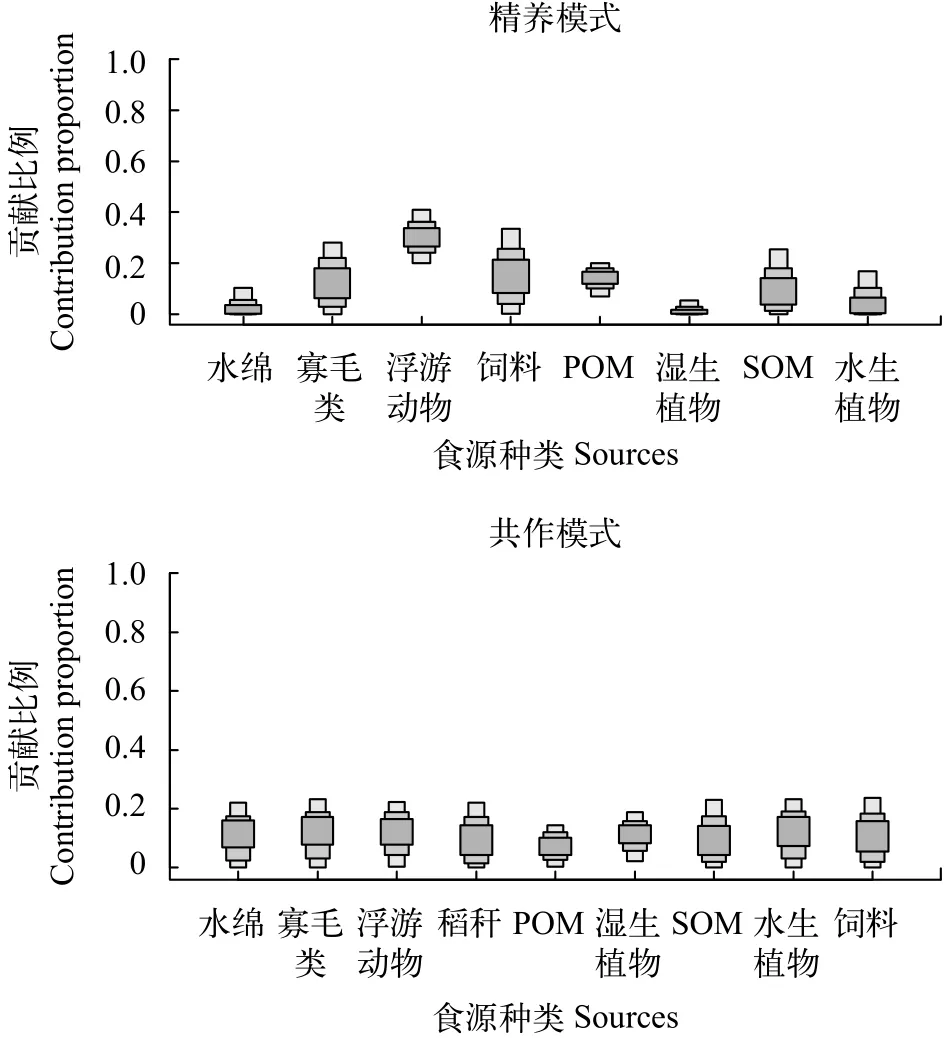
图5 利用SIAR分析的两种养殖模式下克氏原螯虾食物贡献率Fig. 5 Food contribution to P. clarkia analyzed by the SIAR mixing system
对两种养殖模式中克氏原螯虾的体长、体重与稳定同位素值进行相关性分析结果显示: 在两种养殖模式中, 克氏原螯虾的体长、体重均和其δ15N值存在线性关系, 其体内的15N富集程度随体长、体重增加而逐渐增大, 而δ13C值和体长、体重均无显著相关性(表4)。δ13C值和体长、体重并无显著相关性说明克氏原螯虾在生长的过程中食源范围并没有发生太大的改变, 而δ15N随着体长体重增大而增大说明随着克氏原螯虾的成长, 其在生态系统中的相对营养位置提高[24]。类似的现象在其他的甲壳类水生动物中也较为常见, 如日本沼虾(Macrobrachium nipponense), 幼虾主要摄食轮虫和原生动物, 而成虾偏向水生昆虫和寡毛类等大型动物; 口虾蛄(Oratosquilla oratoria)也出现了δ15N与其个体体重存在极显著的正相关, 个体大小对口虾蛄δ15N有影响的现象[25]。
根据其他人对于克氏原螯虾的食性研究[5,7,26],结合两种养殖模式生态系统中食源的易获取程度,我们挑选了克氏原螯虾的主要饵料(图5), 克氏原螯虾的δ13C值在这几种饵料的之间, 而且克氏原螯虾的δ15N均高于它们, 因此可以判断克氏原螯虾的食物组成主要来源于这几种饵料。通过SIAR对两种养殖模式生态系统中克氏原螯虾的食性进行计算, 结果显示: 在精养模式中, 克氏原螯虾偏向于摄食浮游动物, 其次是寡毛类和颗粒性饵料(SOM、POM), 对植物性饵料(水绵、湿生植物、水生植物)的摄食较少; 在共作模式中, 各食源对克氏原螯虾的贡献率较为接近, 克氏原螯虾也存在对动物性饵料(寡毛类、浮游动物)的偏好, 但是其对植物性饵料(水绵、湿生植物、水生植物)的摄食高于共作模式, 另外对颗粒性饵料(POM)摄食较少(图5)。
两种模式中的克氏原螯虾都偏向于摄食动物性饵料, 这和Correia等[26]和Oliveira等[27]的研究结果相似, 在本研究中, 两种模式下克氏原螯虾对水生植物类的摄食都较少, 这与李浪平等[4]的研究结果不同, 可能是因为李浪平等[4]采取的是胃含物分析法, 胃含物分析只能反映生物近期的摄食情况,且会高估不易消化的食物的贡献率[6], 而水生植物相对于其他饵料更不易消化, 因此导致其研究结果中水生植物的食源贡献率比本研究更高。虽然两种模式中克氏原螯虾都偏向于摄食动物性饵料, 但是精养模式中浮游动物在克氏原螯虾的食源贡献率具有绝对优势(30.4%), 而共作模式中并不明显,各食源的贡献率较为平均, 且对饲料的利用率较精养模式更低。
两种养殖模式相对比, 在共作模式中克氏原螯虾, 对湿生植物[28]、水绵这种植物性饵料的利用率明显高于精养模式, Alcorlo等[29]研究表明克氏原螯虾的食性情况和食物易得性有很大关系, 水稻种植提供了更为多样的天然食物, 增加了系统的复杂程度[7,28], 克氏原螯虾更容易获取不同类型的饵料食物, 故共作模式中克氏原螯虾的食性较精养模式更为均匀。除了食性较为均匀, 共作模式中克氏原螯虾对于绿色植物的利用率高于精养模式, Mcclain等[30]研究表明绿色植物可以作为克氏原螯虾的类胡萝卜素和植物甾醇的来源, 而且虾类可以将获得的类胡萝卜素转化为虾青素[31,32], 虾青素对虾类的健康养殖有益, 所以共作模式中克氏原螯虾的食性情况可能更利于其健康生长、繁殖。
4 结论
在两种模式下克氏原螯虾的生态空间利用率接近, 但是共作模式中生物群落的营养结构多样性较高, 群落的营养生态位范围较宽, 食物网的营养冗余程度较低, 生态系统稳定性较高; 克氏原螯虾的δ15N值随着体长、体重增加而增加, 其营养级随着体型的增大而增高; 两种模式相比, 共作模式中克氏原螯虾的各食源贡献率较精养模式更为均匀,植物性饵料贡献率较精养模式更高。
致谢:
感谢中国科学院水生生物研究所分析测试中心的左艳霞老师在仪器使用方面提供的便利!
猜你喜欢
实用手外科杂志(2022年2期)2022-08-31
当代水产(2022年6期)2022-06-29
文萃报·周五版(2022年15期)2022-04-21
读者(2022年8期)2022-04-07
今日农业(2021年11期)2021-08-13
中国生殖健康(2019年8期)2019-01-07
幸福(2018年33期)2018-12-05
现代园艺(2017年19期)2018-01-19
扬子江诗刊(2016年2期)2016-11-25
海峡姐妹(2016年5期)2016-02-27
