
谷子NBS-LRR类基因家族全基因组鉴定及表达分析
2020-03-11 10:43李任建申哲源李旭凯韩渊怀张宝俊
河南农业科学 2020年2期
李任建,申哲源,李旭凯,韩渊怀,张宝俊
(1.山西农业大学 农学院,山西 太谷 030801; 2.山西农业大学 生命科学院,山西 太谷 030801)
谷子[Setariaitalica(L.) Beauv.]为禾本科狗尾草属二倍体C4植物,距今已有9 000多年的栽培历史。谷子具有耐干旱、耐贫瘠、适应性强等特点[1],在我国西北、华北、东北地区广泛种植,年种植面积21万~23万hm2。谷子脱壳后称小米,富含蛋白质、维生素B2、烟酸、钙、铁等营养物质,对糖尿病、皮肤病、心脑血管疾病等多种疾病有食疗作用[2],以小米为主的杂粮逐渐成为人们餐桌的必备食品。但谷子病虫害较多,特别是以谷子白发病、谷瘟病及谷子锈病等为主的谷子病害在种植区普遍发生,对特色杂粮的生产造成了严重威胁[3]。因此,寻求更为安全有效的防治措施、发掘抗病相关基因和关键调控因子以加快谷子抗病育种进程等具有重大意义。
NBS-LRR(Nucleotide binding site-leucine rich repeat)类基因家族是植物中所含抗病基因数目最多的基因家族[4]。目前,克隆到的抗病基因大多数是NBS-LRR类家族基因,具有NBS和富含亮氨酸的LRR结构,NBS结构域为该类家族最保守的区域[5],其中存在8个保守基序(Motif):P-loop、RNBS-A、Kinase 2、RNBS-B、RNBS-C、GLPLA、RNBS-D和MHDV[6]。根据NBS的N末端结构,将含有TIR(Toll interleukin-1 receptor)结构的一类蛋白质定义为TNL(TIR-NBS-LRR)类蛋白,将N末端有卷曲螺旋(Coil-coil,CC)结构的非TNL类蛋白定义为CNL(CC-NBS-LRR)类蛋白[7]。NBS-LRR类抗病基因介导的抗性主要发生在效应因子触发的免疫反应(Effector-triggered immunity,ETI)阶段,通过编码抗病蛋白识别病菌无毒基因编码的效应蛋白,从而激活植物体内相关的抗病信号途径,并在侵染部位产生局部的细胞和组织过敏性坏死(Hypersensitive response,HR),阻碍病菌的进一步扩展[8]。随着高通量测序的迅速发展,研究人员已经对水稻、乌拉尔小麦、玉米等禾本科类作物中的NBS-LRR类基因家族进行了全基因组鉴定和分类[9]。谷子全基因组测序在2012年完成[10],为在基因组水平上解析各类抗病基因特征提供了便利。目前,尚未见NBS-LRR类基因家族全基因组鉴定的研究。为此,利用生物信息学方法,基于HMMER和Blast同源比对相结合的方法对谷子NBS-LRR类家族基因进行筛选和鉴定,同时利用基因组间共线性分析和选择压力分析研究谷子NBS-LRR类家族基因的遗传进化特征,以期为NBS-LRR类家族基因在谷子抗病分子育种中的应用奠定基础。
1 材料和方法
1.1 全基因组序列获取
从Phytozome数据库(https://phytozome.jgi.doe.gov/pz/portal.html)中获取谷子(Setariaitalicav2.2)、狗尾草(Setariaviridisv1.1)、水稻(Oryza sativa v7_JGI)和拟南芥(ArabidopsisthalianaTAIR10)的基因组数据,并从NCBI数据库(https://www.ncbi.nlm.nih.gov/pubmed)下载完整注释为NBS-LRR的36条水稻蛋白质序列和12条拟南芥蛋白质序列。
1.2 谷子NBS-LRR类基因的鉴定及基因定位分析
利用Pfam数据库(http://pfam.xfam.org/)下载含有NBS-LRR类结构域的HMMER模型(PF00931)。通过HMMER软件中的hmmsearch命令对谷子的全基因组蛋白质数据进行搜索[11],筛选阈值E<0.01的基因为谷子NBS-LRR类候选基因。通过Blast软件对水稻和拟南芥蛋白质序列与谷子的蛋白质序列进行比对,筛选阈值E<1e-5的基因为谷子NBS-LRR类候选基因[12]。使用Pfam数据库中的Batch search(http://pfam.xfam.org/search#searchBatchBlock)对HMMER和同源比对得到的候选基因进行进一步验证。利用Paircoil2(http://cb.csail.mit.edu/cb/paircoil2/paircoil2.html)对候选基因进行卷曲螺旋结构域预测[13]。
利用谷子的基因组注释gff文件,对筛选到的NBS-LRR类家族基因进行位置信息的提取,利用TBtools[14]对基因定位的结果进行分析。
1.3 谷子NBS-LRR类基因的序列特征和系统进化分析
由于谷子中NBS-LRR类基因数目较多,故选取55条长度完整的谷子NBS-LRR类基因编码的氨基酸序列进行比对,利用Jalview对序列比对结果进行可视化。对所筛选的411个谷子NBS-LRR类家族基因通过MEGA 7[15]构建邻近树(NJ),设置Bootstrap为500次重复。
1.4 谷子NBS-LRR类基因的结构和Motif分析
由于谷子中NBS-LRR类基因数目较多,故选取80条长度完整的NBS-LRR类基因编码的氨基酸序列,利用MEGA 7[15]分析进化关系。使用TBtools工具对筛选的80条NBS-LRR类基因序列进行基因结构分析;利用在线工具MEME(http://meme-suite.org/)[16]对所筛选的80条NBS-LRR类基因编码的氨基酸序列进行Motif预测,使用TBtools进行可视化。
1.5 谷子NBS-LRR类基因组间共线性及选择压力分析
采用MCScanX[17]软件分析谷子与拟南芥、水稻、狗尾草种间的共线性,使用TBtools对结果进行可视化。利用DnaSP 6[18]软件对存在的共线性基因进行选择压力分析。
1.6 谷子NBS-LRR类家族基因表达模式分析
谷子转录组数据来源于NCBI的SRA(Sequence Read Archive)数据库。利用FastQC[19-20]软件对原始测序数据进行质量评估,再利用Trimmomatic[21]软件去除接头,过滤低质量的reads,得到clean data。利用Hisat2[22]软件进行比对。从Phytozome数据库(https://phytozome.jgi.doe.gov/pz/portal.html#!info?alias=Org_Sitalica)下载谷子基因组数据,采用StringTie[23]进行表达分析,利用R包绘制表达热图。
2 结果与分析
2.1 谷子NBS-LRR类基因的鉴定及染色体定位
利用含有NBS-LRR类结构域的HMMER模型共鉴定到460个NBS-LRR类候选基因;利用Blast比对鉴定到387个NBS-LRR类候选基因;通过Pfam数据库分析,最终筛选到含有NBS结构的基因411个(表1)。根据结构域划分,只含有NBS结构的基因仅有Seita.7G163800,既含有NBS结构又含有CC结构的基因有376个,同时含有NBS结构、LRR结构和CC结构的基因有33个,谷子中无TNL结构基因,仅有1个TN结构基因为Seita.1G161200。通过基因定位发现,NBS-LRR类家族基因中,有158个基因定位于8号染色体,57个基因定位于2号染色体,仅有16个基因定位于1号染色体(图1)。

表1 谷子NBS-LRR类家族成员分类Tab.1 Membership classification of NBS-LRR family genes in Setaria italica
2.2 谷子NBS-LRR类基因的序列比对和进化分析
由图2可知,谷子NBS-LRR类基因中存在4段保守序列,Ⅰ类序列为--G--G-GKT-;Ⅱ类序列为--V-DD---;Ⅲ类序列为---TTR---;Ⅳ类序列为-G-PL--。在系统进化分析中发现,谷子中NBS-LRR类家族基因大致分为3组,大多数具有CC结构的基因被聚类到Ⅰ组,谷子中仅有的1个TIR结构基因Seita.1G161200聚类到Ⅲ组(图3)。
2.3 谷子NBS-LRR类基因的结构及Motif预测
选择谷子中长度完整的80个NBS-LRR类家族基因进行基因结构分析及Motif预测。结果表明,有8个基因无UTR区,其余均存在UTR区,有36个基因存在2段NBS区域(图4A—B)。对80条NBS-LRR类基因编码氨基酸序列进行Motif预测发现,其含有8个Motif,较保守的Motif为Motif1(P-loop)、Motif2(Kinase 2)、Motif3(GLPLA)和Motif5(Kinase 3)(图4C、 图5);大多数NBS-LRR类蛋白中的Motif以Motif5、Motif1、Motif3、Motif6、Motif4、Motif2、Motif8、Motif7的顺序排列,其中Motif3、Motif2、Motif7、Motif4会发生移位,Motif5在每个NBS-LRR类蛋白上较其他Motif保守,仅Seita.2G176400和Seita.3G369800不含有Motif5。

图1 谷子NBS-LRR类家族成员基因染色体定位Fig.1 Chromosome location of NBS-LRR family members in Setaria italica
2.4 谷子NBS-LRR类基因的共线性分析及选择压力分析
共线性分析结果(图6)表明,谷子与狗尾草有23个NBS-LRR类基因存在共线性,与拟南芥中仅有1个基因存在共线性,与水稻有21个基因存在共线性。其中,Seita.8G088500的水稻共线性基因LOC_Os11g11810为已鉴定的Pi5抗稻瘟病基因,Seita.8G088400的水稻共线性基因LOC_Os11g11790为已鉴定的Pia抗稻瘟病基因。利用DnaSP 6[18]软件对存在的共线性基因进行选择压力分析(表2)发现,谷子与水稻、拟南芥共线性基因的Ka/Ks值均小于1,表明谷子与水稻、拟南芥的共线性基因在进化中可能受到纯化选择,Seita.8G088500和Seita.8G088400可能在进化中未发生结构功能上的改变;在与狗尾草共线性基因的Ka/Ks分析中发现,Seita.6G014500、Seita.6G023500的Ka/Ks值大于1,表明这2个基因在进化过程中存在正选择。
2.5 谷子NBS-LRR类家族基因表达模式分析
对NBS-LRR类家族基因的表达情况进行分析(图7)发现,除13个基因(Seita.8G136700、Seita.3G400800、Seita.4G250500、Seita.4G244400、Seita.8G244000、Seita.4G244000、Seita.6G021300、Seita.8G124300、Seita.8G239400、Seita.6G014700、Seita.2G015000、Seita.8G166700、Seita.2G10300)在谷子各组织中均未检测到表达外,其余NBS-LRR类家族基因在谷子根、叶、幼芽、穗等组织中表达;表达模式可分为4类:Ⅰ类在叶中特异性表达,Ⅱ类在幼芽中特异性表达,Ⅲ类在穗中特异性表达,Ⅳ类在根中特异性表达,且在根和穗中高表达的基因数目较多,幼芽中最少,表明NBS-LRR类家族基因的表达可能存在组织特异性。

图2 谷子NBS结构序列特征分析Fig.2 Characteristic analysis of NBS structural sequence in Setaria italica
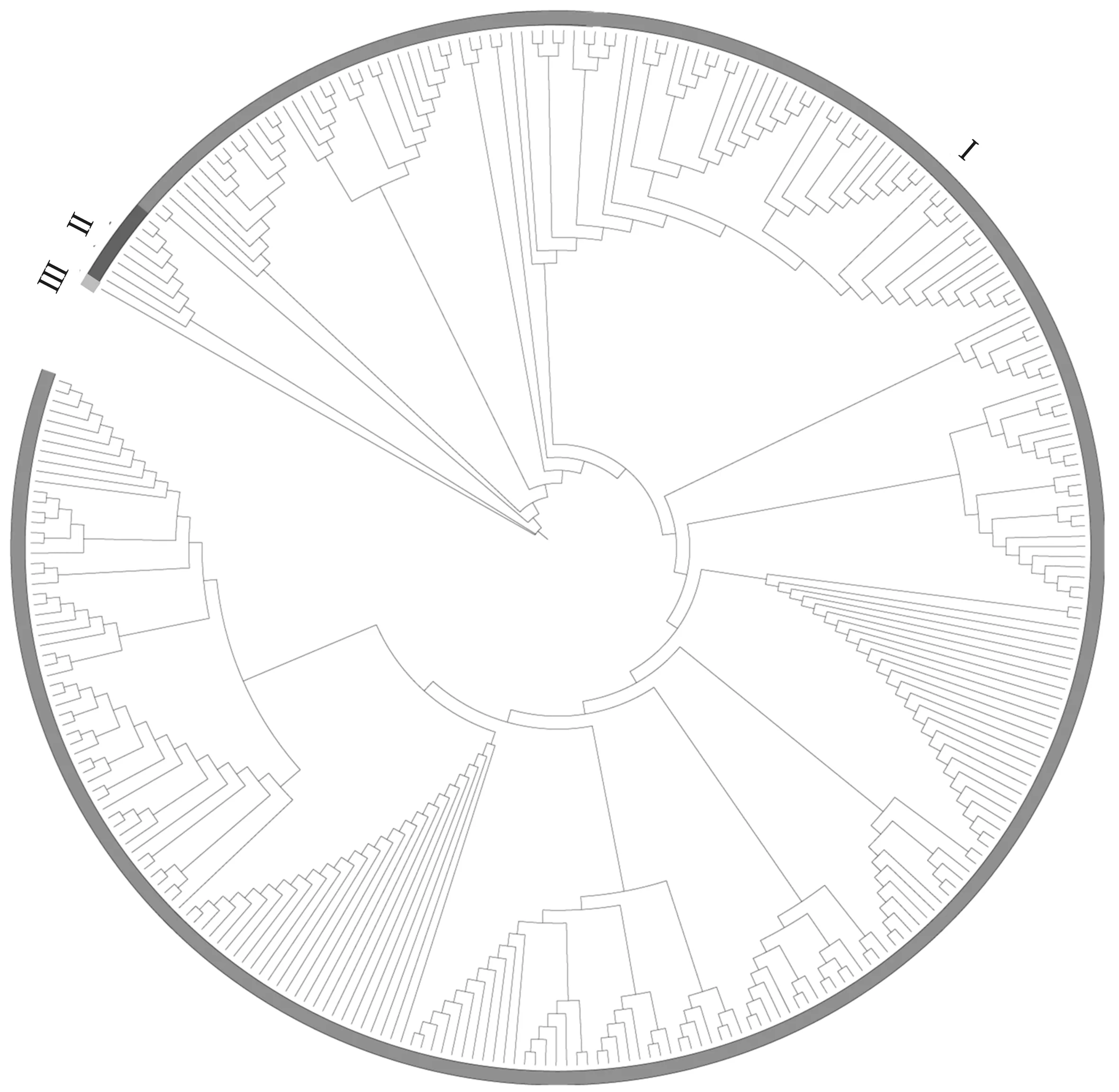
图3 谷子NBS-LRR类基因家族系统进化树 Fig.3 Phylogenetic tree of NBS-LRR family genes in Setaria italica

图4 谷子部分NBS-LRR类基因的进化 (A)、基因结构 (B)及Motif分析 (C)Fig.4 Analysis of evolution,gene structure and motif of NBS-LRR genes in Setaria italica

图5 谷子 NBS-LRR类家族基因Motif结构分析Fig.5 Analysis of motif structure of NBS-LRR family genes in Setaria italica

图6 谷子与狗尾草、水稻及拟南芥中NBS-LRR类基因的共线性分析Fig.6 Colinearity analysis of NBS-LRR genes between Setaria italica and Setaria viridis,Oryza sativa,Arabidopsis
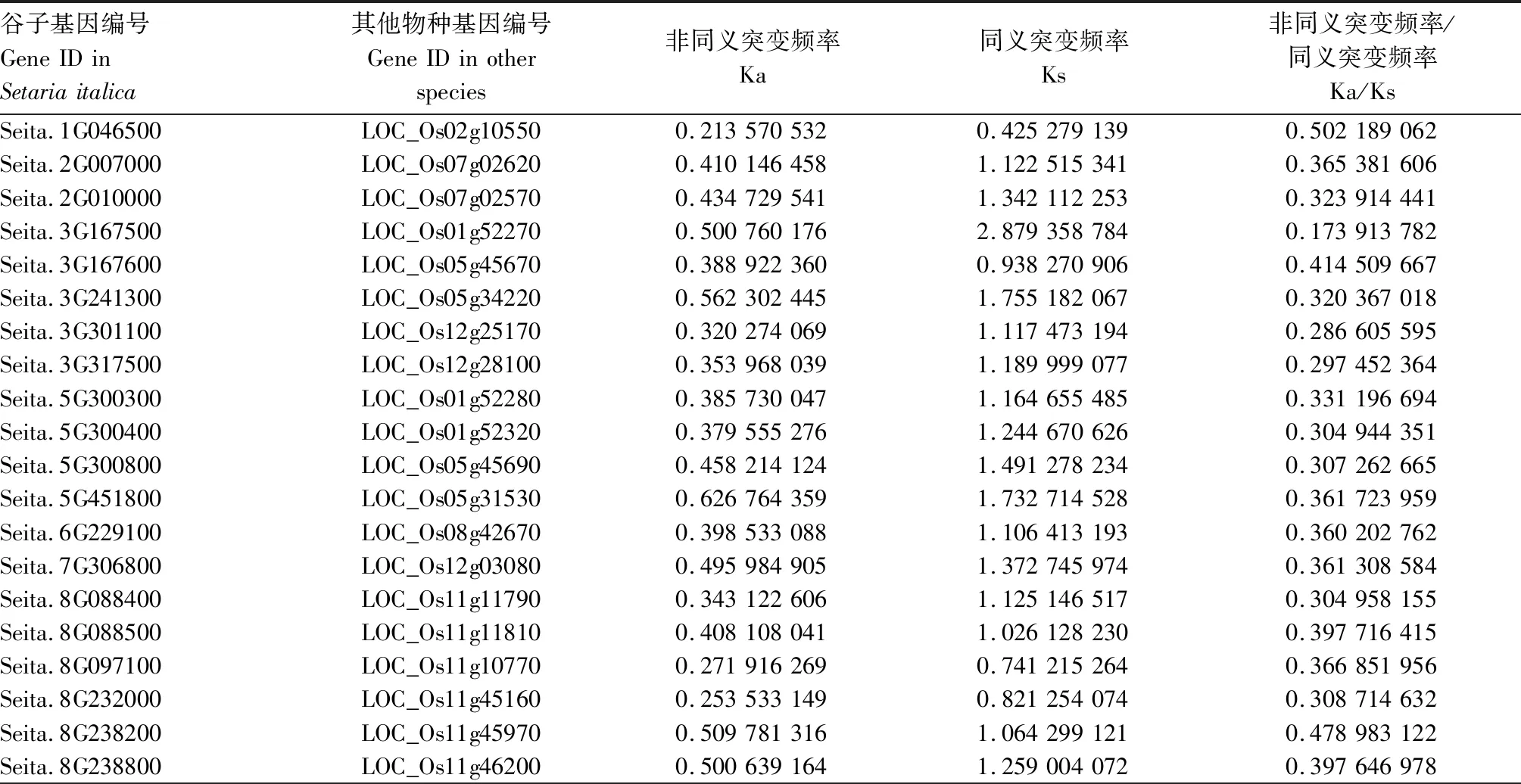
表2 谷子NBS-LRR类基因进化选择压力分析Tab.2 Analysis of evolutionary selection pressure of NBS-LRR genes in Setaria italica

续表2 谷子NBS-LRR类基因进化选择压力分析Tab.2(Continued) Analysis of evolutionary selection pressure of NBS-LRR genes in Setaria italica

图7 谷子不同组织中NBS-LRR类家族基因的表达谱Fig.7 Expression profile of NBS-LRR genes of Setaria italica in different tissues
3 结论与讨论
NBS-LRR类家族基因作为抗病基因中数目最多的家族之一,在植物响应生物胁迫中发挥重要作用。NBS-LRR类家族基因一般通过超表达启动受侵染部位的程序性死亡,阻止病原菌的扩展[24]。目前,多个NBS-LRR类抗病基因已被克隆,如拟南芥抗霜霉病基因RPs4[25]、水稻抗稻瘟病基因Pi-2[26]和水稻抗白叶枯基因XA21[27]等。本研究共鉴定出411个NBS-LRR类家族基因,其中定位到8号染色体上的基因最多。基因结构和Motif分析表明,多数基因具有相似的结构及组成,Motif的组成及排布也有一定的规律,部分Motif有位置和数量的变化,可能与谷子在病原菌等生物胁迫或非生物胁迫条件下产生的遗传变异相关,这与乌拉尔小麦[28]、油菜[29]的研究结果类似。在共线性分析中发现,谷子与狗尾草共线性基因数目最多,水稻次之,与拟南芥最少,一方面因为禾本科作物亲缘性更高,另一方面可能是在进化过程中发生了基因丢失或该家族基因进化速度较快,这种现象在禾本科植物中普遍存在[30]。在与狗尾草NBS-LRR类基因的选择压力分析中发现,仅有2个NBS-LRR类基因发生了纯化选择,说明狗尾草在驯化为谷子的过程中,可能受到了自然选择作用,并且多数基因进化速度较快。
NBS-LRR类家族基因的组织特异性表达在小麦[31]、水稻[32]中已有报道,但目前尚无该家族基因在谷子中的组织表达特异性研究,本研究对该家族基因在根、穗、幼芽及叶中的表达情况进行分析发现,除少数基因在不同组织中均有表达外,多数基因均在特定的组织中高表达,表明该家族基因在谷子中也可能存在组织特异性表达的特点,且在根部及穗部抗病过程中发挥重要作用。
致谢:感谢本科生申哲源参与部分试验数据的处理,感谢李旭凯老师和韩渊怀老师对本研究的方法和思路提供建议,感谢张宝俊老师指导数据处理、论文写作和修改。
猜你喜欢
今日农业(2022年4期)2022-06-01
广西植物(2021年1期)2021-03-24
园艺与种苗(2020年12期)2021-01-08
科学与财富(2020年33期)2020-03-10
发明与创新·大科技(2019年5期)2019-07-31
温州大学学报(自然科学版)(2019年2期)2019-06-04
现代商贸工业(2019年5期)2019-02-18
飞碟探索(2015年9期)2015-11-05
红领巾·探索(2015年9期)2015-09-10
恋爱婚姻家庭·养生版(2011年8期)2011-05-14
