
骨髓间充质干细胞外泌体改善缺氧诱导内皮细胞损伤与STAT3的相关性
2020-03-18 09:03孙石峰李烈友冯志海陈良龙
福建医科大学学报 2020年6期
范 林, 孙石峰, 江 琼, 李烈友, 冯志海, 陈 煌, 陈良龙
血管内皮损伤是众多缺血性疾病的共同病理基础,有效阻止和减轻内皮损伤对延缓缺血性疾病的发展进程具有重要意义。既往研究显示,干细胞外泌体能够减轻各种因素刺激下的内皮细胞损伤,但多聚焦于传递治疗性微小RNA[1-3]。对于是否能够通过传递蛋白发挥保护性作用,目前尚不明确。
信号转导和转录激活因子3(signal transducer and activator of transcription, STAT3)是STAT家族成员之一,作为重要的转录因子在细胞内起着信号传递作用,当受到细胞外信号刺激,活化的JAK,ERK1/2和Akt1能够与STAT3蛋白的相应位点(如Y705,S727)结合,磷酸化修饰STAT3并使之活化,活化的STAT3以二聚体形式进入细胞核,诱导靶基因表达,进而调控细胞功能。研究表明,STAT3介导了干细胞的心血管保护效应,人子宫内膜干细胞移植能够增强血管内皮细胞增殖及内皮管形成能力[4]。近来,国外学者通过蛋白质组学分析发现,骨髓间充质干细胞(mesenchymal stem cells,MSCs)来源的外泌体中含有大量蛋白激酶和转录因子等蛋白分子,包括STAT3[5]。但干细胞外泌体能否通过传递STAT3蛋白发挥内皮细胞的保护作用尚不明确。本研究拟采用携带绿色荧光蛋白(green fluorescence protein, GFP)的慢病毒转染标记大鼠骨髓MSCs,采用缺氧无血清培养构建内皮细胞损伤模型,探讨干细胞外泌体减轻内皮细胞损伤的作用及可能机制。
1 材料与方法
1.1主要材料 Sprague Dawley(SD)大鼠骨髓MSCs(BNCC100382)和大鼠主动脉内皮细胞(rat aorta endothelial cell, RAEC)(BNCC341217)(北京北纳创联生物技术研究院);携带GFP的慢病毒原液(2×108转导单位/mL)(上海吉凯基因化学技术有限公司);低糖DMEM培养基及胎牛血清(美国Gibco公司);外泌体提取试剂盒(美国Invitrgon公司);重组抗CD9抗体(ab92726)、重组抗CD81抗体(ab109201)、重组抗STAT3抗体(ab68153)、重组抗STAT3 (phospho Y705)抗体(ab76315)、重组抗血管内皮细胞生长因子(vascular endothelial growth factor, VEGF)抗体(ab32152)、重组抗血管生成素2(angiopoietin 2/Ang2)抗体(ab155106)、抗缺氧诱导因子1(hypoxia-inducible factor-1, HIF-1)抗体(ab216842)(均为英国ABCAM公司产品);重组抗CD63抗体(OM245785)(美国OmnimAbs公司);BCA法蛋白浓度测定试剂盒(上海碧云天生物科技有限公司);cDNA合成试剂盒(美国Promega公司);实时荧光定量聚合酶链反应(qRT-PCR)试剂盒(美国Bimake公司)。
1.2方法
1.2.1构建稳定表达GFP的大鼠骨髓MSCs 以每孔2.5×105个细胞的密度接种大鼠骨髓MSCs于6孔板,在37 ℃、体积分数为0.05的CO2培养箱中,以含10%胎牛血清的低糖DMEM培养基培养,待细胞达30%~40%融合度时,以感染复数(multiplicity of infection, MOI)为15的重组慢病毒进行感染,48 h后通过荧光显微镜观察细胞状态,再以5 μg/mL嘌呤霉素筛选感染48 h后的细胞,每2天换液1次,以未做病毒感染处理的细胞为对照;筛选后的细胞在培养箱中继续培养48 h(培养条件同上),荧光显微镜下观察GFP的表达情况,并于细胞达80%~90%融合度时传代冻存。
1.2.2外泌体提取与扫描电镜鉴定 将稳定表达GFP的大鼠骨髓MSCs接种于10 cm培养皿中,待细胞增殖至80%融合度时,更换无血清DMEM培养基,放入厌氧培养盒中缺氧培养48 h,收集细胞上清,以外泌体分离提取专用试剂盒提取外泌体,具体按试剂盒说明书操作。提取的外泌体以锇酸溶液4 ℃固定,乙醇梯度脱水后丙酮包埋,将包埋板置于65 ℃下聚合48 h,然后以醋酸双氧铀和醋酸铅各染色10 min,清洗后上机检测。
1.2.3大鼠RAEC缺氧模型的建立与分组 取长势良好的RAEC 细胞,胰酶消化,接种量为24 h后达70%的融合度为宜。细胞分为3组:对照组、缺氧组和外泌体+缺氧组。细胞置于厌氧培养盒中以无血清培养基培养构建细胞缺氧模型,具体培养时间根据不同实验而定,以正常条件培养的细胞为对照。外泌体工作终浓度为50 μg/mL。
1.2.4CCK-8检测RAEC细胞的增殖活性 取对数期生长的RAEC细胞,按每孔3×103个接种,每孔100 μL完全培养基,每组设6个复孔,置于37 ℃、体积分数为0.05的CO2培养箱中培养16 h,吸弃原培养基,根据分组要求,加入相应培养液进行干预,分别于24,48及72 h后取出,细胞每孔加入10 μL CCK-8溶液,37 ℃培养箱中孵育4 h后取出,酶标仪450 nm波长下检测OD值。
1.2.5黏附实验 用10 μg/mL的纤连蛋白预铺96孔板,每孔100 μL,4 ℃过夜,用1% BSA于37 ℃封闭1 h。取分组干预48 h后的RAEC细胞,按每孔5×103个接种,每孔100 μL完全培养基,每组设3个复孔,37 ℃孵育1 h,PBS洗去未黏附的细胞;4%多聚甲醛室温固定30 min,PBS洗涤 3遍;加入100 μL的姬姆萨染料染色15 min,PBS洗涤后,在光镜200倍视野下随机选取6个不同视野进行计数。
1.2.6Transwell迁移实验 细胞接种于T25培养瓶,按分组要求加入相应培养液干预48 h后,收集细胞并调整密度为每室5×104个接种于小室中,加入无血清培养基200 μL,下室加入500 μL完全培养基(含10% FBS),将带有小室的24孔板置于细胞培养箱(37 ℃、体积分数为0.05的CO2)中培养,18 h后取出,PBS洗涤,每孔加入预冷的500 μL固定液固定20 min,以结晶紫500 μL染色30 min,PBS洗涤3次;在倒置显微镜400倍视野下随机选取6个不同视野进行计数,取平均数,实验重复3次后进行统计分析。
1.2.7内皮管形成实验 取对数期生长的细胞接种于T25培养瓶,加入相应培养液干预48 h,以每孔3×104个细胞加入铺有Matrigel的96孔板中,每孔100 μL完全培养基,每组3个复孔,37 ℃培养12 h(每隔一段时间观察),选取最佳成管时间;在倒置显微镜100倍视野下观察内皮管形成效果,并随机选取视野拍照计算成管个数。

1.2.9qRT-PCR法检测VEGF,Ang-2和HIF-1基因mRNA表达 细胞干预同Western-blot实验,提取总RNA,以Promega反转录试剂盒合成cDNA第1链,以β-actin为内参照,使用qRT-PCR试剂盒及特异性引物进行扩增,采用2-ΔΔCt法计算基因相对表达量,所有特异性引物序列见表1。

表1 实时定量聚合酶链反应引物序列
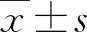
2 结 果
2.1大鼠骨髓MSCs荧光表达及其外泌体形态和蛋白表达 GFP稳定表达的大鼠骨髓MSCs培养3 d后,细胞融合度达90%以上,呈平行旋涡状排列(图1A),荧光显微镜下见细胞均匀散发绿色荧光(图1B);扫描电镜下见外泌体直径30~100 nm,呈杯状双层膜结构(图1C);Western-blot检测证实外泌体标志蛋白CD9,CD81和CD63表达,且内含有丰富的STAT3蛋白(图1D)。

A:光镜下大鼠骨髓间充质干细胞(MSCs)呈旋涡状平行排列(×100);B:荧光显微镜下MSCs均匀散发绿色荧光(×100);C:扫描电镜下外泌体形态呈杯状双层膜结构;D:Western-blot检测外泌体蛋白.图1 大鼠骨髓间充质干细胞绿色荧光表达及其外泌体形态和蛋白表达Fig.1 Green fluorescence expression of rat bone marrow mesenchymal stem cells (MSCs) and MSCs-derived exosomes' morphology and protein expression
2.2大鼠骨髓MSCs外泌体对缺氧损伤RAEC增殖的影响 CCK-8法检测RAEC增殖结果显示(图2),24 h后,缺氧组的OD值(0.83±0.09)明显低于对照组(1.20±0.24)(P<0.05),与外泌体+缺氧组(0.94±0.10)比较,差别无统计学意义(P=0.448);48 h后,缺氧组的OD值进一步下降,与对照组(1.82±0.21)比较,差别有统计学意义,而外泌体+缺氧组(1.32±0.14)明显高于缺氧组(0.99±0.09)(P=0.001);72 h后,对照组的OD值为(2.27±0.12),明显高于缺氧组和外泌体+缺氧组(P<0.001),但外泌体+缺氧组(1.24±0.13)明显高于缺氧组(0.82±0.11)(P<0.001)。

n=6. 与缺氧组和缺氧+外泌体组比较,△:P<0.001;与缺氧组比较,☆:P=0.001,#:P<0.001.图2 3组间细胞增殖比较Fig.2 Comparison of cell proliferation among three groups
2.3大鼠骨髓MSCs外泌体对缺氧损伤RAEC黏附、迁移、内皮管形成的影响 缺氧组姬姆萨染色黏附细胞数及结晶紫染色的迁移细胞数均明显少于对照组,外泌体干预后两种细胞数均明显多于缺氧组(图3A,3B)。同样,外泌体+缺氧组内皮管数量少于对照组,但明显多于缺氧组(图3C)。进一步定量统计分析发现,对照组黏附、迁移细胞数和内皮管数分别为(553.0±28.1),(740.0±17.4)和(13.2±1.5)个;缺氧无血清培养后,与对照组比较,细胞黏附、迁移和内皮管形成能力明显受损,黏附、迁移细胞数和内皮管数分别为(114.0±15.6),(202.7±19.8)和(2.5±0.6)个(均P<0.05);而外泌体干预后,RAEC细胞功能明显改善,黏附、迁移细胞数和内皮管数分别为(224.7±21.5),(387.7±22.1)和(9.3±0.6)个,与缺氧组比较,差别有统计学意义(均P<0.05,图4)。

A:姬姆萨染色检测细胞黏附(×200),外泌体+缺氧组(A3)阳性细胞明显多于缺氧组(A2);B:结晶紫染色检测细胞迁移(×400),外泌体+缺氧组(B3)阳性细胞明显多于缺氧组(B2);C:内皮管形成实验(×100),外泌体+缺氧组(C3)内皮管明显多于缺氧组(C2).图3 3组细胞黏附、迁移和内皮管形成Fig.3 Representive images of cell adhesion, migration and tube formation in three groups

n=3. A:200倍视野下各组黏附细胞比较;B:400倍视野下各组迁移细胞比较; C:100倍视野下各组内皮管数量比较. 与对照组比较,☆:P<0.001;与缺氧组比较,△:P<0.001.图4 光镜下3组间细胞黏附、迁移和内皮管形成比较Fig.4 Comparison of cell adhesion, migration and tube formation among three groups under light microscope
2.4大鼠骨髓MSCs外泌体对缺氧损伤RAEC的STAT3,VEGF,Ang-2和HIF-1蛋白表达 与对照组比较,缺氧诱导RAEC细胞VEGF,Ang-2和HIF-1蛋白表达上调(均P<0.05);予外泌体干预后,VEGF,Ang-2和HIF-1蛋白表达进一步增高;尽管缺氧组与对照组比较,STAT3蛋白表达无明显差别,但缺氧刺激后p-STAT3表达明显增强,而外泌体+缺氧组STAT3和p-STAT3的表达均明显高于缺氧组和对照组(P<0.01,图5)。
2.5大鼠骨髓MSCs外泌体对缺氧损伤RAEC的VEGF,Ang-2和HIF-1基因的mRNA表达 与对照组比较,缺氧组VEGF,Ang-2和HIF-1基因的mRNA表达均明显升高(P<0.05),且于外泌体干预后进一步上调(P<0.05,图6)。
2.6外泌体与STAT3蛋白共表达 对照组和缺氧组未见GFP表达,STAT3表达呈弱阳性,外泌体+缺氧组RAEC可见GFP表达,STAT3表达(红色荧光)明显强于对照组和缺氧组,并且GFP定位与STAT3蛋白一致(图7)。

n=3. VEGF:血管内皮细胞生长因子;Ang-2:血管生成素2;HIF-1:缺氧诱导因子1;STAT3:信号转导与转录激活因子3;p-STAT3:磷酸化STAT3蛋白. A~E分别展示了VEGF,Ang-2,HIF-1,STAT3和p-STAT3蛋白表达水平;F:各个基因代表性蛋白电泳图. 与对照组比较,☆:P<0.05,☆☆:P<0.05;与缺氧组比较,#:P<0.05,##:P<0.01.图5 3组间VEGF,Ang-2,HIF-1,STAT3和p-STAT3蛋白表达比较Fig.5 Comparison of VEGF, Ang-2, HIF-1, STAT3 and phosphorylated STAT3 protein expression among three groups

n=3. VEGF:血管内皮细胞生长因子;Ang-2:血管生成素2;HIF-1:缺氧诱导因子1. 与对照组比较,☆:P<0.05;与缺氧组比较,#:P<0.05.图6 3组间VEGF,Ang-2和HIF-1基因的mRNA表达比较Fig.6 Comparison of VEGF, Ang-2 and HIF-1 mRNA expression among three groups
3 讨 论
血管内皮损伤是众多缺血性疾病发生发展的始动及核心环节,如何有效减轻血管内皮损伤已成为此类疾病防治的研究热点之一。本研究利用缺氧无血清诱导RAEC损伤模型,发现骨髓MSCs来源的外泌体能够改善缺氧状态下内皮细胞功能并促进成血管活性,可能与传递STAT3蛋白进而上调VEGF,Ang-2和HIF-1基因的mRNA和蛋白表达有关。
众所周知,血管内皮细胞是位于血管表面的单层扁平细胞,调节血液和组织间物质代谢,起着维持血管稳态的作用。缺血缺氧作为血管内皮损伤的主要因素之一,可通过损伤血管内皮结构、破坏血液-组织屏障,进而导致缺血性疾病的发生发展。与众多研究结果一致[6-8],本研究发现,缺氧无血清培养能够有效诱导血管内皮细胞损伤,表现为血管内皮细胞增殖、黏附、迁移及成血管活性受损,这为后续干预研究提供了有力保障。

图7 激光共聚焦扫描显微镜下绿色荧光蛋白和STAT3蛋白共表达Fig.7 Co-expression of GFP and STAT3 under laser confocal scanning microscope
虽然研究已证实骨髓MSCs可直接通过内皮细胞定向分化和(或)旁分泌机制减轻血管内皮损伤[9-10],但缺血组织中移植细胞的低存活率及成瘤风险限制了其临床应用。近年来研究发现,外泌体是MSCs等多种干细胞移植治疗发挥生物学作用的关键效应介质,不仅体积足够小、易于通过组织屏障,而且无干细胞移植治疗的潜在成瘤性风险[11-12]。目前,已有研究发现,干细胞外泌体不仅可改善缺血心脏的心功能,还能够诱导缺血组织血管新生[13-14],有望成为干细胞治疗的有效替代工具。本研究通过电镜和Western-blot技术证实成功获取MSCs来源外泌体,并发现将MSCs外泌体添加至缺氧无血清培养的RAEC,能够明显改善细胞增殖、黏附、迁移能力和成血管活性。
尽管如此,但MSCs外泌体保护损伤内皮细胞的机制至今仍未完全清楚。理论上,外泌体可通过传递蛋白、microRNA及DNA等介导细胞间信号交互,进而发挥生物学效应。既往蛋白组学研究显示,骨髓MSCs外泌体中含有STAT3蛋白,但其功能角色尚无定论[5]。本研究通过Western-blot印迹法证实,骨髓MSCs外泌体富含STAT3蛋白,且于外泌体干预后表达明显增加,而且以p-STAT3为主。共聚焦扫描显微镜也发现,外泌体与STAT3共定位于缺氧损伤的血管内皮细胞内,提示MSCs外泌体可传递STAT3至缺氧无血清培养的血管内皮细胞并发生磷酸化修饰。结合功能学研究结果,进一步证实MSCs外泌体对缺氧无血清损伤内皮细胞发挥保护作用可能与传递STAT3蛋白有关。
此外,本研究还探讨了STAT3蛋白对下游保护性基因的调控作用。既往研究已明确,血管内皮细胞功能稳态涉及众多细胞因子或基因[15]。体外研究发现,添加外源性VEGF可促进内皮细胞存活,改善内皮细胞增殖、黏附、细胞迁移,促进血管新生[16]。而HIF-1可促进VEGF和Ang-2等基因的转录激活,在缺氧条件下,HIF-1过表达能够明显增加血管内皮细胞Ang-2和VEGF表达,促进血管生成[17]。虽然当内皮细胞受到缺血缺氧损伤后,也通过上调VEGF,Ang-2和HIF-1发挥内源性保护作用,但低水平上调并不足以对抗缺血缺氧损伤。本研究结果显示,缺氧组VEGF,Ang-2和HIF-1的蛋白和mRNA表达水平均明显高于对照组,但其细胞功能(如增殖、黏附、迁移和成血管活性)仍比对照组差。既往血管生成和神经再生等领域的研究发现,VEGF,Ang-2和HIF-1作为STAT3的下游靶基因,STAT3蛋白经磷酸化修饰后可上调VEGF,Ang-2和HIF-1[4,18-21]。本研究发现,MSCs外泌体干预后的RAEC,p-STAT3表达明显增高,同时伴随VEGF,Ang-2和HIF-1表达上调,提示MSCs外泌体的内皮细胞保护作用机制可能与传递STAT3至内皮细胞并发生磷酸化后激活VEGF,Ang-2和HIF-1表达有关。
本研究存在一定局限性:如未进行外泌体量效研究,且研究中未使用STAT3抑制剂或RNA干扰进行反向验证。但研究结果表明,骨髓MSCs外泌体可以减轻缺氧无血清诱导的内皮细胞损伤,并探讨了STAT3蛋白在外泌体介导的内皮保护作用中的功能角色,这不仅为将来缺血性疾病的防治提供了新的靶点,也有助于拓展临床医师对外泌体细胞保护作用机制的理解。
猜你喜欢
基础医学与临床(2022年1期)2022-11-25
现代泌尿外科杂志(2022年3期)2022-11-23
中国生物制品学杂志(2022年3期)2022-11-15
中南药学(2022年2期)2022-11-14
体育科技文献通报(2022年4期)2022-10-21
空军军医大学学报(2022年5期)2022-10-12
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中国眼耳鼻喉科杂志(2022年3期)2022-05-29
昆明医科大学学报(2022年2期)2022-03-29
中国循证心血管医学杂志(2022年1期)2022-03-15
