
E.coli K-12琥珀酸脱氢酶sdhC基因纳米锥的设计及自组装
2020-04-11 06:30周末刘彦君佟欣瑞董延甲吴欣瑜王迎香应明
化工进展 2020年2期
周末,刘彦君,佟欣瑞,董延甲,吴欣瑜,王迎香,应明
(1天津理工大学化学化工学院,天津300384;2天津理工大学计算机科学与工程学院,天津300384)
除小部分病毒外,所有生命的遗传物质都是双链DNA。DNA 分子不仅具有特异性识别能力,而且遵守严格的自组装机制,因此天然的DNA 分子是研制靶向药物载体的优选材料。近几年悄然发展起来的DNA折纸术(DNA origami),就是一种采用计算机精准设计进行核酸纳米材料自组装的新方法[1-2]。2008 年丹麦Aarhus 大学的Andersen[3]开发了一款UNIX 系统适用的DNA 折纸软件——SARSE,利用该软件首先进行了DNA 二维结构设计,成功得到了约50nm×150nm 尾巴酷似在摆动的海豚图案;不久后该团队又设计出一个带有可“开-关”盖子的DNA 盒子,将该软件用于了DNA 三维折叠设计[4]。DNA 折纸用到最多的开源软件是Douglas设计的caDNAno,研究者可利用其中的蜂巢模型或方格模型进行脚手架链和订书钉链的设计,在二维界面构建堆叠DNA 图形[5]。麻省理工学院(MIT)Bathe 团队研发的CanDo 和DAEDALUS,是目前进行DNA 三维立体设计功能最完善的软件,CanDo是一款在线分析软件,能依据caDNAno 的设计图进行DNA立体结构预测[6];DAEDALUS 是由Matlab编写的涵盖多种立体构象的源代码,Bathe 团队依靠这种软件,采用M13mp18 单链环形DNA 构建出了立方体、正八面体、正二十面体等纳米结构DNA[7],最近该团队还报道了使用caDNAno 构建了结构复杂的荷花型和贝壳形DNA纳米材料[8]。2006年上海交通大学Bio-X 中心,同样采用M13mp18单链核酸折叠出一幅中国地图[9]。从前期研究可以看出,人们已经能够将一个单链核酸折叠成各种尽可能复杂的图形,M13mp18 单链环形DNA 是进行这种单链折叠的主要材料。但如何将DNA 折纸术用于普通基因组核酸材料,如何将这项技术应用到实际的研究领域中,都是该项技术是否能得到广泛应用的关键。
本文作者项目组即以此为目的,采用生物实验室最常用的模式菌株E.coliK-12 MG1655,作为琥珀酸脱氢酶sdhC 基因的供体菌。sdhC 是中心代谢的一个基因,在生命体中普遍存在,其研究结果具有一定的代表性。首先利用改进后的DAEDALUS软件,设计出与sdhC 基因序列互补的16 条订书钉链[10]。这16 条订书钉链与PCR 扩增得到的sdhC 双链DNA 进行链内置换,最终组装出了与预设模型基本一致的正四面锥。在实验中发现,sdhC 基因的核酸链在自组装前后,DNA 凝胶电泳条带的位置发生了明显变化。采用扫描电子显微镜(SEM)和透射电子显微镜(TEM)进行观察,发现样品为均匀分散的锥形颗粒,尺度约为15.02~23.97nm。采用原子力显微镜进行该纳米锥液下的三维立体结构成像,可观测到一个完整清晰的正四面锥体,说明大肠杆菌的普通基因序列,被成功组装成了一个对称的纳米锥结构。这种核酸纳米锥具有很强的生物相容性,可以帮助尺度适宜的药物进入目标细胞,提高药物精准释放和靶向作用的能力。
1 实验材料和方法
1.1 sdhC基因纳米锥设计
首 先 改 进 DAEDALUS 软 件 中 的DX_cage_design模块,使该软件能读取sdhC基因的核酸序列。按照Waston-Crick 的B 型DNA 双螺旋模型,每螺旋为10.5 个碱基对时,DNA 折叠的应力最小,所以待设计的正四面锥的每条棱应有n10.5bp(n为正整数)个碱基,且由两条订书钉单链和两条脚手架链组成。在DAEDALUS源代码中,加入一个求余函数rem(edge_length,21),检测每条棱的碱基数是否为21bp 的整数倍,如果不是,程序则会自动在棱上增加5、10 或11 个碱基。同理,DAEDALUS 输出订书钉链也会尽量接近n21bp,以保证核酸自组装时不产生过度弯曲[11]。理论上DAEDALUS 允许输入脚手架链的碱基个数没有限制,但是源代码中的参数均依照M13mp18单链DNA(7249bp)设定。为重新设定计算参数,对sdhC 基因序列进行了上下游延伸处理,即在sdhC基因序列上下游各补充117bp,其基因图谱如图1 所示,总长为624bp,其中118~507bp 为sdhC基因。将该624bp 的碱基序列作为脚手架链输入DAEDALUS软件,脚手架链序列如表1所示。经过计算正四面锥各棱含有两组碱基数为52bp 的DNA双链,图1中的箭头表示折叠后每条棱对应的碱基位置,AD 棱的碱基序列包括(1,33)、(606,624)、(138,189);CD棱包括(34,86)、(503,553);BD 棱包括(87,137)、(242,294);AB 棱包括(190,241)、(348,399);BC 棱包括(295,347)、(452,502);AC棱包括(400,451)、(554,605)。

表1 脚手架链碱基序列
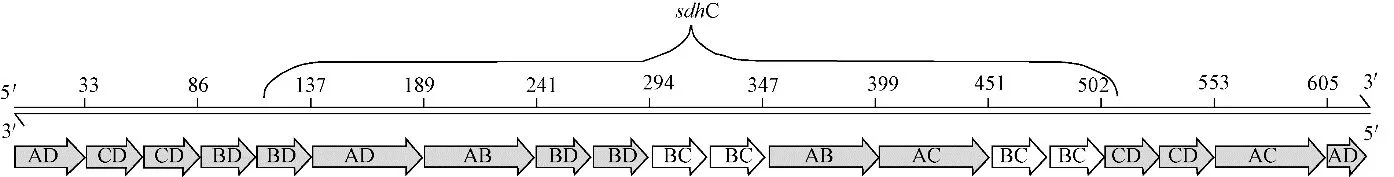
图1 sdhC基因纳米锥脚手架链基因图谱

图2 sdhC基因纳米锥设计图(1Å=0.1nm)
图2为sdhC基因纳米锥的设计过程,深色粗实线代表脚手架链,浅色粗实线代表订书钉链。624bp 的脚手架链在16 条互补的订书钉链作用下,不中断地来回折叠,每条棱都是由两条脚手架单链和两条互补的订书钉单链相互缠绕形成[12]。如图2(c)所示,正四面锥6条棱的设计略有不同,根据脚手架链的形态可分为两组:第一组AB、AC、AD为无隔断棱;第二组BC、BD、CD 为有隔断棱,脚手架链的3’端和5’端均在AD棱上。订书钉链共有4 种类型,分别为回型(Clip)、钩型(Hook)、夹型(Pin)以及凸型(Convex)。图2(a)给出了与顶点B相连的3条棱的结构,每条棱两端皆是部分凸型链,BC 与BD 棱中部由两个31bp 的钩型钉书钉链形成,AB棱中部由一条长42bp回型订书钉链和一条20bp 夹型订书钉链形成。棱结构AD、AC 与AB 相同,BD、CD 与BC 相同。顶点B由78bp凸型订书钉链形成,如图2(b)所示,且各顶角的订书钉链形状、长短皆相同。其中每个构成顶角的凸型订书钉链中加入了一段长5bp 的poly(T)结构,与脚手架链不匹配,形成一个单链的折角。表2为在DAEDALUS输出的16条订书钉链的序列,带下划线的碱基表示poly(T)结构。图2(c)为DAEDALUS 设计出的最终纳米锥的立体结构,最终形成每条棱包含52bp 的正四面体,长度为17.68nm,按照B 型DNA 双螺旋模型计算,每两个碱基之间距离为0.34nm,所以每条棱长是52×0.34nm=17.68nm,每条棱的两条脚手架链长度相同,均为52bp,且上下各错开一个碱基,以形成拐点,如图2(a)所示。
1.2 实验材料
大肠杆菌(E.coliK-12 MG1655)甘油菌为本实验室保藏[13];引物、订书钉链、Taq DNA 聚合酶由TaKaRa 公司合成;DNA MarkerII 购自北京鼎国生物公司。实验所用仪器包括:凝胶电泳仪(DYCP-31CN 型,北京六一生物科技有限公司);场发射扫描电镜(MERLIN Compact 型,德国ZEISS 公司);原子力显微镜(ICON 型,德国Bruker公司);透射电子显微镜(JEM-2100型,日本JEOL 电子株式会社);PCR 仪(T-Gradient Thermoblock型,德国Biometra公司)
1.3 PCR法制备脚手架链
在Genebank中查询的E.coliK-12 MG1655sdhC基因序列(Gene ID:945316,390bp),并在上下游各补充117bp,得到624bp 的碱基序列。依据该序 列 设 计 上 下 引 物 : Primer-1 (5′-TGTCTGACCCGCAAGC-3′) ; Primer-2 (5′-CGCCACTGGTAGCGA-3′)。 提 取E. coliK-12 MG1655染色体,作为扩增脚手架链的模板,进行PCR 反 应[3]。PCR 反 应 体 系 为:10×PCR Buffer(Mg2+) 5µL; 4×dNTP Mixture 4µL; Primer-1(27.6µmol/L)1µL;Primer-2(28.2µmol/L)1µL;E.coliK-12 MG1655 染色体(150nmol/L)2µL;Ex Taq 酶0.25µL 及无菌水36.75µL 混匀,构成50µL PCR 反应体系。PCR 程序为:94℃预变性4min,94℃变性1min,60℃退火30s,72℃延伸1.5min,从第二步循环30轮后,72℃保温10min。实验结束后进行琼脂糖凝胶电泳检测[14]。

表2 订书钉链碱基序列
1.4 自组装sdhC基因纳米锥
在200µL薄壁管中,将脚手架链和订书钉链按照1∶10加入到自组装缓冲溶液TAE/Mg2+中(2mol/L Tris,0.1mol/L CH3COOH,0.05mol/L 乙二胺四乙酸,12.5mmol/L Mg(CH3COO)2),混合成50µL 自组装体系,放入可梯度控温的PCR 仪中,组装程序为:94℃变性4min,以0.6℃/s 的降温速率降温至20℃[15],4℃保存10min。实验结束后取出样品进行琼脂糖凝胶电泳(AGE)、扫描电子显微镜(SEM)、透射电子显微镜(TEM)和原子力显微镜(AFM)进行检测。
2 结果与讨论
2.1 sdhC基因纳米锥自组装策略
实验中采用双链DNA 作为DNA 折纸的脚手架链,双链DNA 是包括sdhC 基因在内的624bp 双链PCR 产物。因此与以往使用单链DNA 作为脚手架链的自组装策略不同,本研究采用的是一种链置换组装策略,如图3(b)所示。当加入的订书钉链浓度远远超过双链脚手架链时,在DNA 变性和复性过程中,订书钉链取代了双链DNA 中的一条链,发生链内置换,将另一条链折叠为预设的核酸模型。实验的每一步均进行了琼脂糖凝胶电泳验证,图3(a)为PCR 得到的双链脚手架链电泳图,大小为624bp,图3(c)为组装前后的电泳对比图,泳道1为16 条订书钉链,均为21~78nt 的短链核苷酸,泳道2为双链脚手架链,泳道3为自组装产物,很明显组装前后的电泳条带存在一定差距,而且没有检测到订书钉链,说明订书钉链与脚手架链结合后,脚手架链发生了折叠,形成了与组装前不同的空间结构[16]。
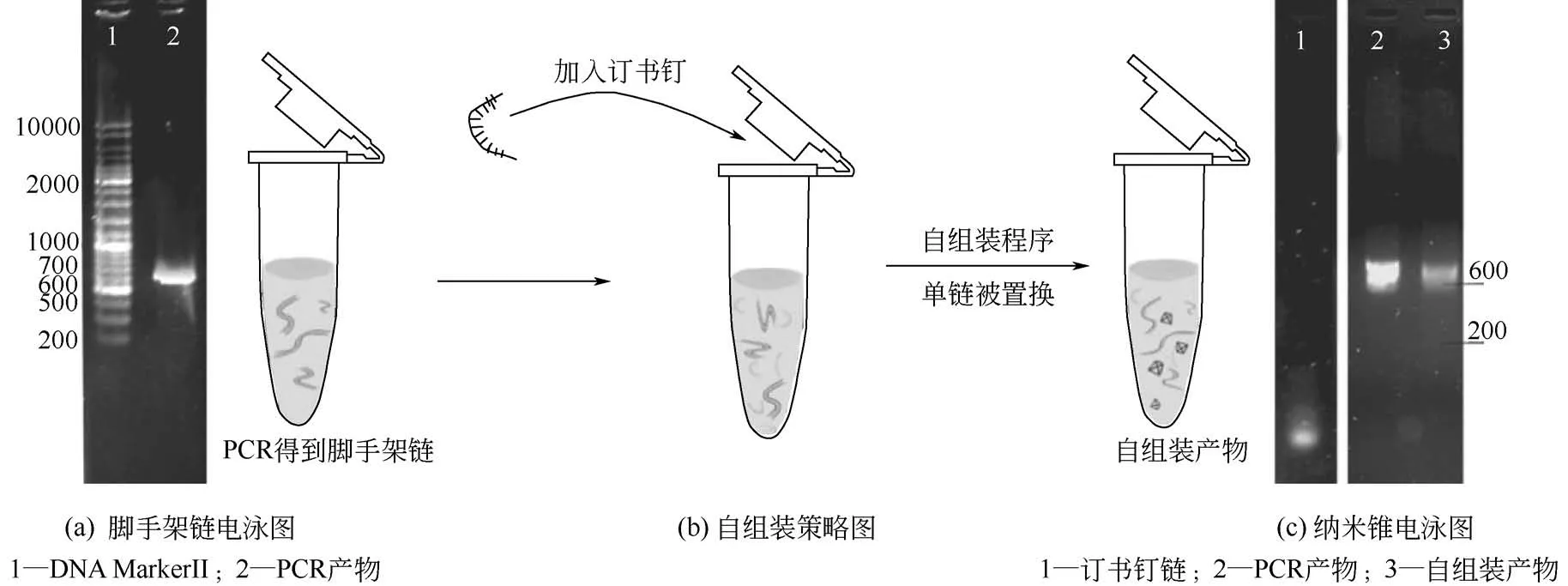
图3 sdhC基因纳米锥组装策略及凝胶电泳检测
2.2 sdhC基因纳米锥形貌分析
为了进一步分析折叠后的DNA 空间构象,将得到的自组装产物超声震荡15~20min,取1µL 样品涂布在云母片中央,沉积过夜,镀金后使用SEM 进行扫描检测。图4 是在工作电压10kV,工作距离13.3mm,放大倍数10万倍条件下的sdhC基因纳米锥全貌图。从图中可以看出,自组装产物为均匀分散、边缘清晰且结构完整的锥形纳米颗粒,而在涂布了TAE/Mg2+的空白云母片上,未观察到任何纳米颗粒;左上角的插图2 为一个锥体的放大图,与插图1中预设模型基本一致。通过对多次实验样品的不同视野中几百个颗粒的测定,得到的纳米锥平均边长为19.05nm,与最初设计仅相差1.37nm,这有可能是仪器检测和手动测量产生的误差。图5为SEM的能谱报告,滴加了样品的云母片C、O的含量分别升高了9.3%、29.58%,样品中还检测出含0.19%的N 和1.64%的P,说明样品为生物大分子。为了观测自组装产物在溶液中的形貌,本研究组采用TEM,取2µL 样品进行了检测,结果如图6 所示,得到的三角锥边长在(17.68±2)nm范围内,与SEM测定的尺度基本相符。

图4 SEM扫描图像

图5 能谱分析
2.3 AFM分析

图6 TEM扫描图像
为了能更清晰地分析样品在液体中的立体构型,本研究使用AFM 进行3D 分析。将新解离的1.3cm×1.8cm 云母片固定在2cm×2.5cm 的干净玻璃片上,用透明胶将云母片表层清洁干净,滴加3µL样品于云母片中央,沉积3min。将BRUKER SNL-10 镀金扫描探针,固定在液体探针夹上,并滴加30µL TAE/Mg2+缓冲溶液,进行液下检测。AFM 检测结果如图7所示,样品表面受探针压力影响产生了一定的形变,但仍能观察到清晰的锥体结构,图7(a)是其中一个纳米锥的3D 图像,其最高峰约为2.2nm,推测应该是纳米锥预设模型中的B 顶点,与B 顶点相连的3 条棱分别是CB、AB 和DB。图7(b)为将探针与样品间作用力转化为样品高度的曲线图,即记录了探针扫描过样品每个点高度的数据,可以看出探针的走向是从一个低点升至一个最高点,再回到一个低点,3个方向趋势相同,扫描过的距离即为3条棱CB、AB、DB的长度,分别为23.838nm、 15.245nm、 18.654nm, 平 均 为19.246nm;与预设模型的边长相差1.566nm,基本符合预期设计。产生误差的原因,有可能是原子力显微镜的探针对样品产生的压力,导致样品形变,使测定边长与实际边长产生偏差。
3 结论

图7 sdhC基因纳米锥原子力显微镜3D成像图
本研究采用E.coliK-12 MG1655琥珀酸脱氢酶C 亚基基因sdhC 核酸序列构造出一种核酸纳米锥体,是首次使用普通基因的双链DNA 进行DNA 折纸术设计。经过琼脂糖凝胶电泳(AGE)、扫描电子显微镜(SEM)、透射电子显微镜(TEM)和原子力显微镜(AFM)多种检测手段验证,sdhC 基因的双链DNA 确实被组装成了与DAEDALUS 软件预设模型基本一致的正四面锥,但尺度上存在微小差距,且不同表征方式检测出的差距很接近。除了考虑是仪器检测产生的测量误差外,还有可能是组装体系中的离子强度影响了B型DNA的螺旋方式,导致DNA 复性时与标准Watson-Crick 模型产生了偏差。所以在今后的研究中,还需要进一步探索组装体系和双链DNA 长度对自组装效率的影响,以及如何解决置换出的单链对自组装过程干扰的问题。总之,本研究证实了双链DNA 可以作为DNA折纸术的核酸材料,甚至可以按照药物分子的需求“定制”相应的DNA药物载体。例如,sdhC基因纳米锥可以载入边长17nm 以内的药物分子,帮助药物在细胞内定点释放出,尤其是在恶性肿瘤细胞的精准治疗中发挥一定的作用。
猜你喜欢
力学学报(2021年2期)2021-03-10
海外星云 (2021年21期)2021-01-19
供应链管理(2020年10期)2020-12-23
教学考试(高考生物)(2020年6期)2020-11-23
食品与生物技术学报(2020年8期)2020-01-06
中国海洋大学学报(自然科学版)(2019年2期)2019-12-07
学苑创造·B版(2019年5期)2019-06-14
科学24小时(2019年5期)2019-06-11
智富时代(2018年9期)2018-10-19
智富时代(2018年9期)2018-10-19
