
不同光照强度下白及光合生理特性的研究
2020-05-22 02:56崔波周一冉王喜蒙袁秀云蒋素华许申平
河南农业大学学报 2020年2期
崔波,周一冉,王喜蒙,袁秀云,蒋素华,许申平
(1.郑州大学生命科学学院,河南 郑州 450001; 2.郑州师范学院生物工程研究中心,河南 郑州 450044)
白及(Bletillastriata)是兰科白及属多年生草本植物,其块茎广泛用于治疗皮肤皲裂、疮疡肿毒、外伤出血、烧伤烫伤、咳血吐血、胃溃疡等症,很早就被收录在中国药典中[1]。除药用价值外,白及还具有较高的观赏价值和经济价值[2-4]。长期以来,白及药材主要依赖野生采挖,野生资源被过度开采。2017年白及已被《濒危野生动植物种国际贸易公约》附录Ⅱ收录,而目前人工栽培技术体系尚不成熟,市场供需矛盾日渐紧张[5-6]。目前,有关白及的研究多集中在药理作用、遗传多样性、组培苗再生、化学成分与地理分布等方面[7-10]。随着对白及的研究逐渐深入,白及种苗繁育技术也日渐成熟[11],而白及人工栽培条件下的光合生理生化研究还比较欠缺。光照是植物生长发育的物质和能量来源,不同植物具有特有的光照适应性[12-15]。植物为适应不同光照环境会产生一系列光化学变化,如对车前轻度遮光可提高其光化学效率和电子传递效率,降低热耗散,而全光照和重度遮光会降低其光合能力[16];长春花在弱光下叶绿素含量上升,但重度弱光胁迫会使其光合速率、ΦPSⅡ、qP和qN下降,并导致光合机构发育不良[17];紫背天葵在轻度遮光条件下叶片具有更高的最大光合速率、光补偿点、光饱和点、瞬时水分利用效率及光能利用率[18]。吴明开等[19]通过对白及光合与蒸腾生理生态及抗旱特性研究认为,白及为阴生植物,栽培时应注意适度遮光,适宜白及生长的具体遮光度目前并不清楚。本研究在人工栽培条件下设置4种光照强度,研究白及在不同光照强度下的光合、荧光及生理指标,分析白及对不同光强的生理响应和动态变化,进一步探讨白及对光照的适应特性,为白及大规模栽培及管理提供理论依据。
1 材料与方法
以2 a生大田栽培白及为材料,于2018年5—9月在郑州师范学院兰花工程技术研究中心实践基地进行试验。采用遮光率为50%、70%、90%的黑色遮光网分别对栽培白及进行遮光处理,全光照为对照(0,CK),每个处理进行3个生物学重复。
1.1 光合参数的测定
5—9月,每月选择3个连续无云的晴天,9:00—11:00用Li-6800便捷式光合系统测定仪(LI-COR,USA)测量叶片净光合速率(Pn,μmol·m-2·s-1)、气孔导度(Gs,mol·m-2·s-1)、蒸腾速率(E,mol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1),并测定光合有效辐射(PAR,μmol·m-2·s-1)日变化,每个处理测量4株,每株测1个叶片,每个叶片重复3次,选取空间取向基本一致的叶片进行测定。
1.2 光响应曲线的测定
使用Li-6800便捷式光合系统测定仪测定光响应曲线,设定光照强度梯度为:1 500、1 200、1 000、800、500、300、100、50、20、0 μmol·m-2·s-1,测定时每一光照强度下停留120~200 s,测量前将待测叶片在1 000 μmol·m-2·s-1光照强度下诱导30 min活化光合系统。CO2浓度通过外接CO2小钢瓶控制,浓度设为接近大气CO2浓度的400 μmol·mol-1,根据非直角双曲线模型[20]回归法求得光补偿点(LCP,μmol·m-2·s-1)、光饱和点(LSP,μmol·m-2·s-1)、暗呼吸速率(Rd,μmol·m-2·s-1)及最大净光合速率(Pnmax,μmol·m-2·s-1)。
1.3 叶绿素荧光参数的测定
使用PAM-2500便捷式叶绿素荧光分析仪(Walz,Germany)测定白及叶片叶绿素荧光诱导动力学参数。参照许申平等[21]的方法测定光化学猝灭系数qP、PSⅡ最大光化学效率Fv/Fm、非光化学猝灭系数qN、表观电子传递速率ETR等叶绿素荧光参数,每个处理测4株并取平均值。测定暗适应下荧光参数时,叶片提前暗适应30 min。
1.4 光合色素含量的测定
叶绿素含量的测定采用80%丙酮浸提法,采用Lichtenthaler的公式[22]计算叶绿素含量。
1.5 抗氧化相关酶活性的测定
过氧化氢酶(CAT)活性的测定采用紫外吸收法[23],过氧化物酶(POD)活性的测定参考CHEN等[24]的方法,超氧化物歧化酶(SOD)活性测定采用WST法[25]。
1.6 数据分析
采用SPSS 20.0进行数据差异性分析。
2 结果与分析
2.1 不同光照强度对白及叶片光合参数的影响
图1显示,5—9月各处理白及叶片光合有效辐射(PAR)的日变化呈明显的单峰曲线,并在11:00—13:00之间达到最高峰,同时,PAR在不同光照条件下的大小依次为CK>50%>70%>90%。
不同光照条件下白及叶片净光合速率Pn测定结果表明(图2-A),从5月到9月,白及叶片Pn呈现先升高后降低的趋势;不同光照条件对Pn的影响有较大差异, 5月70%遮光条件下Pn显著高于其他处理,6—9月50%遮光条件下Pn较高,分别比CK高68.27%、102.74%、42.16%、53.49%,并在7月达到最大值7.66 μmol·m-2·s-1;90%遮光条件Pn一直处于较低状态,9月其Pn只有1.35 μmol·m-2·s-1;CK条件下Pn始终低于50%和70%遮光处理。结果表明,全光照和90%遮光条件均不利于白及叶片光合作用的进行,50%和70%的遮光条件有利于提高白及叶片的净光合速率。
胞间CO2浓度Ci体现大气的输入和细胞光呼吸、光合利用CO2的瞬间动态平衡浓度。在白及旺盛生长的5—9月,90%遮光条件下白及叶片Ci始终较高,而其Pn相对较低;在5月,70%和90%遮光条件下Ci显著高于其他2个处理(图2-B);在6—9月,遮光条件下白及叶片的Ci均高于CK条件,50%和70%遮光条件下Ci没有显著差异。



图1 5—9月不同遮光条件下白及叶片PAR的日变化Fig.1 Daily variation of PAR in leaves of Bletilla striata under different shading treatments from May to September
气孔导度Gs的大小直接影响植物的光合作用和蒸腾作用[25]。3个遮光条件下白及叶片Gs呈先升高后降低的趋势,CK条件下Gs始终偏低(图2-C);在5月,70%遮光条件下Gs显著高于其他处理,比CK高138.16%;在6—7月,50%、70%和90%遮光条件下Gs有明显提高,50%遮光条件下Gs显著高于其他处理,Gs从高到低依次为50%>70%>90%>CK;6—9月,尽管CK条件下白及叶片Pn显著高于90%遮光条件,但其Gs却显著低于90%遮光条件。
由图2-D可知,蒸腾速率E的变化与Gs变化相似,呈先升高后降低的趋势,6—9月50%遮光条件下白及叶片E显著高于其他处理,并在7月达到最大值,此时比CK高225.61%;在6—7月50%遮光条件E显著高于其他处理,而在6—9月CK条件下Gs、E较其他处理偏低;在8—9月,4个处理Pn、Gs、E都有明显下降,表明白及在6—7月生长最为旺盛;遮光有利于提高叶片的Ci、Gs和E。
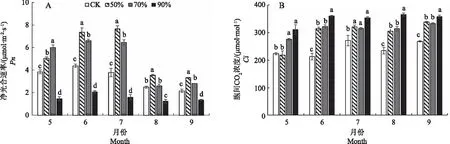
注:不同小写字母表示同一月份不同处理具有显著性差异(P<0.05)。下同。
2.2 不同光照强度对白及叶片光响应参数的影响
表观量子效率AQY代表了植株对光能的利用效率。自然环境下,植株的表观量子效率一般为0.03~0.05 mol·mol-1[29]。本研究结果表明,5月50%和70%遮光条件下白及叶片具有较高水平的AQY、最大净光合速率Pnmax、光饱和点LSP和较低的光补偿点LCP;6月70%遮光条件下AQY、Pnmax、LSP显著高于其他处理,同时LCP较低,表明此时70%遮光条件下白及利用弱光的能力较强;在7—8月,50%遮光条件下白及叶片AQY明显高于其他处理,分别比CK提高了34.48%、23.60%;9月CK条件下Pnmax、AQY和LSP高于3个遮光条件,同时LCP较低;在5—7月,暗呼吸速率Rd随着光照强度的降低而呈逐渐下降趋势。从白及叶片对不同光强的响应来看,4个处理LCP基本维持在10~40 μmol·m-2·s-1之间,LSP均低于1 000 μmol·m-2·s-1(表1),光补偿点和光饱和点均较低,表明白及更适宜于生长在有一定遮光条件的环境。
2.3 不同光照强度对白及叶片叶绿素荧光参数的影响
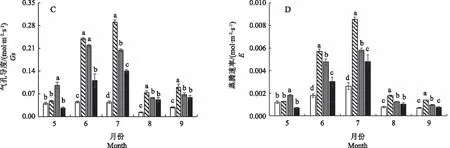
由图3-A可知,在6—8月,3个遮光条件下白及植株Fv/Fm差异不显著,而CK条件下Fv/Fm明显低于遮光条件。光化学淬灭系数qP代表PSII天线色素吸收的光能用于光化学电子传递的比例,反映了光合效率和光能利用效率[27]。本研究中,4个处理条件下qP呈现先升高后降低的趋势,qP与光照强度呈正相关,随着光照强度的降低,qP呈逐渐降低趋势;6月CK处理qP最高,比50%、70%、90% 3个遮光条件分别提高了37.70%、64.50%、118.66%。
由图3-C可知,在5—9月,CK条件下白及叶片qN明显高于3个遮光条件;在5月,70%和90%遮光条件显著降低白及叶片qN,在6—9月,3个遮光条件下的qN差异不大;从5—9月,ETR呈现先升高再降低的趋势,而且ETR的大小高低与光照强度呈正相关(图3-D),90%遮光条件下白及叶片的ETR明显低于其他3个处理。
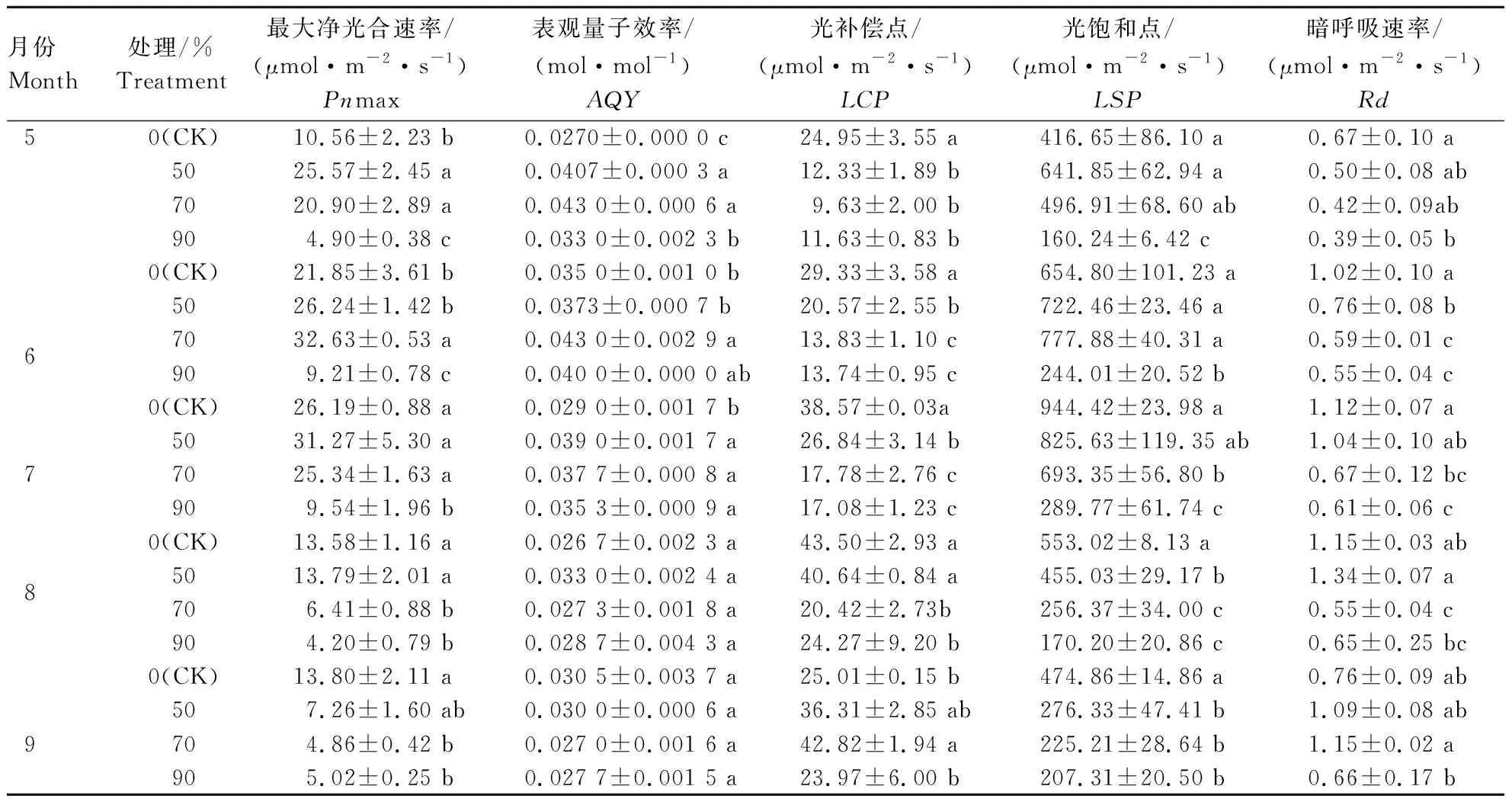
表1 不同光照强度对白及叶片气体交换参数的影响Table 1 Effects of different light intensities on gas exchange parameters in leaves of Bletilla striata
注:同列不同小写字母表示同一月份不同处理在P<0.05水平上差异显著。
Note: Different lower case letters in the same column indicate that different treatments in the same month have significant differences at the level ofP< 0.05.

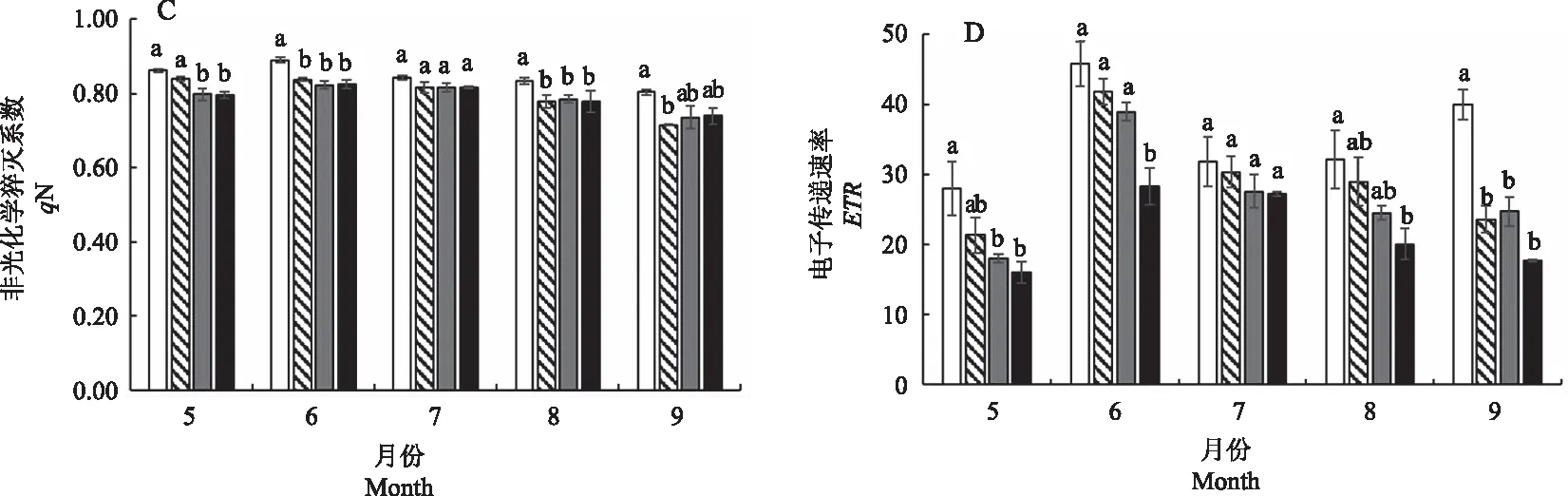
图3 不同光照强度对白及叶片荧光参数的影响Fig.3 Effects of different light intensities on fluorescence parameters of Bletilla striata leaves
2.4 不同光照强度对白及叶片叶绿素含量的影响
不同光照强度下白及叶片叶绿素含量有明显变化(图4-A)。5—8月,3个遮光条件下白及叶片叶绿素含量普遍高于CK处理,50%遮光条件下叶绿素含量在8月为1.73 mg·g-1;70%遮光条件下叶绿素含量在7月最高,此时比CK条件下高10.62%;90%遮光条件下叶绿素含量呈先增高后降低的趋势,并在7月为最大值;随着光照强度的降低,Chla/Chlb比值呈逐渐降低趋势,相对于其他3个处理,90%遮光条件下Chla/Chlb比值始终处于较低水平。

图4 不同光照强度对白及叶片叶绿素含量的影响Fig.4 Effects of different light intensities on photosynthetic pigment contents in Bletilla striata leaves
2.5 不同光照强度对白及叶片抗氧化酶活性的影响
如图5所示,从5月到9月,CK、50%和70%遮光条件下SOD酶活性逐渐升高(图5-A),90%遮光条件下白及叶片SOD酶活性先升高后降低;5—6月各处理SOD酶活性均处于较低水平,7月50%和70%遮光条件下SOD酶活性急剧升高,90%遮光条件下白及叶片SOD酶活性达到最大值,此时3个遮光条件下SOD酶活性分别比CK高712.17%、654.91%、242.45%;8月50%遮光条件下SOD酶活性明显高于其他处理,而90%遮光条件下SOD酶活性显著低于其他处理;9月50%遮光条件下酶活性达到最大值6 936.90 U·g-1,90%遮光条件下酶活性只有247.52 U·g-1。

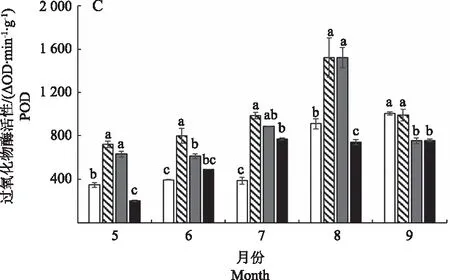
图5 不同光照强度对白及叶片抗氧化酶活性的影响Fig.5 Effects of different light intensities on activities of antioxidant anzymes in Bletilla striata leaves
在5—9月,50%、70%、90%遮光条件下CAT酶活性呈逐渐降低的趋势,CK条件下酶活性先升高后降低(图5-B)。5月,50%和70%遮光条件下CAT酶活性显著高于CK和90%遮光条件,50%遮光条件下白及叶片CAT酶活性比CK高116.68%,比90%遮光条件高121.74%,70%遮光条件下比CK、90%遮光条件高147.87%、153.66%;6月50%遮光条件下CAT酶活性显著高于其他3个处理,5—8月90%遮光条件下,CAT酶活性始终低于其他处理,9月各处理CAT酶活性达到最低值,且70%和90%遮光条件下的CAT酶活性显著低于CK和50%遮光条件。
不同遮光条件下白及叶片POD酶活性在5—9月之间的变化趋势不同(图5-C)。CK和90%遮光条件下POD呈现逐渐升高的趋势,而50%和70%遮光条件下呈现先升高后降低的趋势;5—8月,50%和70%遮光条件下POD酶活性较高,并在8月达到最大值;而9月CK和50%遮光条件POD酶活性显著高于70%和90%遮光条件,同时CK条件下POD酶活性达到最大值。
3 结论与讨论
白及植株在春季进入急速生长阶段,4月开花,5—9月为物质积累阶段,10月末地上部分逐渐枯萎凋谢。不同遮光条件下白及外观形态也有较大差异:CK条件下植株叶片颜色发黄,株型短小,叶脉粗壮突出,7月叶尖开始出现褐色坏死斑点;50%和70%遮光条件下植株生长旺盛,株型高大紧实,叶脉清晰明显;90%遮光条件下植株叶片颜色较深,叶型宽大,但长势萎靡,叶脉触感不明显。不同光照条件明显影响白及叶片的光合及生理代谢,并且在整个生长期内随着不同月份光合有效辐射的不同,光合速率、PSⅡ最大光化学效率、叶绿素含量和抗氧化相关酶活性均有明显差异。
植物在受到胁迫时,影响植物光合作用的因素主要有气孔因素与非气孔因素[28],当Pn的下降伴随着Ci和Gs的降低时,此时影响植物光合能力的原因主要是气孔因素,否则为非气孔因素。本研究中,CK条件下白及叶片Pn降低,Ci、Gs、E始终处于较低水平,表明过度光强对白及植株产生了光抑制,此时白及Pn降低的主要原因是气孔限制,强光下白及通过降低气孔开放程度来减少蒸腾作用,防止叶片过分失水;50%和70%遮光条件下白及叶片保持较高的Pn、Gs、E,说明50%~70%的遮光条件有利于提高白及叶片光合作用;90%遮光条件下,Pn显著降低,而Ci始终高于其他3个处理,此时影响白及光合作用的是非气孔因素,推测认为长期的弱光环境影响了白及光合相关酶的活性或使光合机构受损。
遮光初期(5—6月),50%和70%遮光条件下白及拥有更高水平的Pnmax;遮光后期(7—9月),CK和50%遮光条件的Pnmax较高,表明50%遮光下白及有更高的光合潜能,同时白及为了适应高光强环境,不断提高自身光合潜能;光抑制可导致AQY下降[29],70%遮光条件下AQY在5—6月较高,50%遮光条件下AQY在7—9月处于较高水平,表明50%~70%遮光条件下的白及光合效率更高,利用弱光的能力较强,有利于白及生长;光饱和点和光补偿点的高低体现了植物对光能的利用范围,5—6月50%、70%遮光条件下白及具有更高水平的LSP,而在7—9月CK、50%遮光条件LSP更高,表明50%、70%遮光条件下的白及对光环境的适应性更强,能适应的光强范围更广,同时全光照条件下白及通过不断调节自身光合机构以适应高光强环境,表现出一定的光合可塑性;此外可发现,白及LSP和LCP较低,具有充分利用低光强的光合特性,是一种典型的阴生植物。
叶绿素荧光动力学参数对于观测植物叶片光合作用过程中对光能的吸收、传递、耗散、分配等方面具有重要意义[30]。非胁迫条件下,Fv/Fm值一般为0.8~0.83,是检测植物是否受到胁迫的参数之一[31];光化学淬灭系数qP反映PSⅡ吸收的光能用于光化学电子传递的部分[14],qN反映了PSⅡ吸收的光能以热的形式耗散而非用于光合电子传递的部分[32]。郑淑霞等[33]通过对8种阔叶树种叶片叶绿素荧光特性的研究表明,qP与qN呈极显著正相关关系(P<0.01)。本研究中,3个遮光条件下的白及植株Fv/Fm均维持在较高水平,更接近于0.8,说明一定的遮光环境有助于提高白及光化学效率;而CK条件下较低的Fv/Fm值表明高光强对白及光合系统产生了损伤;但CK条件下较高水平的qP、ETR、qN表明,白及通过增加热耗散的比例,提高电子传递速率以适应高光强环境,避免植株受到永久性光破坏;研究表明,过低的光照环境也会对植物造成光抑制[34],90%遮光条件下,白及叶片qP、ETR低于其他3个处理,说明重度遮光会降低白及光化学效率和电子传递速率,这也可能是90%遮光条件下Fv/Fm较高而其净光合速率较低的原因。
叶绿素含量的高低和比值可以体现出植物利用光能的能力差异,遮光一般会导致透射光中红光比例降低,而蓝光比例升高[35],叶绿素a主要吸收红光部分,而叶绿素b主要吸收蓝紫光部分。本研究结果表明,5—8月3个遮光条件下白及叶片叶绿素含量较高,CK条件下叶绿素含量处于较低水平,前人研究结果显示,遮光下植物叶绿素含量增加,叶绿素a与叶绿素b比值降低,植物通过提高叶绿素b的含量来捕获更多的光能,弥补环境中光强的不足,这是植物对弱光环境的一种适应表现[35-37]。本试验研究结果与前人研究结果一致,表明遮阴条件下白及通过增加叶绿素含量来提高对光能的利用;9月90%遮光条件下白及叶绿素含量降低,推测长期的弱光环境影响了白及叶绿体的正常发育,不利于白及叶片光合作用的进行。
在逆境条件下,植物体内活性氧(ROS)和自由基含量升高[38],膜脂过氧化加剧,细胞正常功能受损。SOD、POD、CAT是植物中抗氧化酶保护系统,在抵抗外界胁迫、降低氧化物质积累、维持膜结构完整等方面起着重要作用[39]。本研究中,不同光照条件下,白及抗氧化系统呈现出不同的反应机制:SOD酶活性逐渐升高,同时50%遮光条件下白及SOD酶活性始终处于较高水平,表明50%遮光条件下白及通过保持较高的SOD酶活性来清除活性氧自由基,从而保护膜系统免受损害;CAT主要起清除线粒体电子传递、β-脂肪酸氧化以及光呼吸等过程中产生的过氧化氢的作用,防止活性氧和自由基对植株造成损害[40]。本研究中,CAT酶活性呈逐渐降低趋势,推测CAT在白及叶片抗氧化初期起主要作用,在抗氧化后期起辅助作用;各处理POD酶活性变化趋势不同,在5—8月50%和70%遮光条件下白及POD活性处于较高水平,表明50%~70%的遮光条件更有利于白及清除过氧化物,降低活性氧对植株的伤害;同时9月CK条件下白及POD酶活性达到最高水平,表明白及通过不断调节自身抗氧化系统机能来适应当前环境。
综上所述,白及更适合生长于有一定遮光的环境下,高光强不利于白及生长,而过度遮光会使白及光合机构受损,50%遮光条件提高了白及光合效率、叶绿素含量以及抗氧化相关系统;长期生活在高光环境中的白及通过不断调节自身光合水平和抗氧化能力来适应高光环境,避免长期高光胁迫对植株造成不可逆损伤;白及为阴生植物,栽培管理期间对其进行50%~70%遮光处理,可有效提高白及的光合效率,降低氧化胁迫对白及造成的损伤,有利于植株生长发育。
猜你喜欢
现代盐化工(2021年3期)2021-08-27
阅读(科学探秘)(2020年8期)2020-11-06
食品与生物技术学报(2020年4期)2020-01-06
绿色科技(2019年2期)2019-05-21
中国果业信息(2019年1期)2019-01-05
绿色科技(2017年17期)2017-09-30
卷宗(2017年15期)2017-07-19
中国中药杂志(2017年8期)2017-05-11
文史杂志(2016年6期)2016-12-03
新课程·下旬(2015年10期)2015-10-21
