
基于无人机的昆明海口林场宽地坝林区植物群落建群种格局研究
2020-06-08 11:00李富利彭晓昶王崇云彭明春谢映旭左艳洁
生态科学 2020年3期
李富利, 彭晓昶, 王崇云, 彭明春, 谢映旭, 左艳洁
基于无人机的昆明海口林场宽地坝林区植物群落建群种格局研究
李富利, 彭晓昶, 王崇云*, 彭明春, 谢映旭, 左艳洁
云南大学生态学与地植物学研究所, 昆明 650500
建群种是植物群落优势层中的主要物种, 对群落环境有显著控制作用。空间格局是植物种群的基本特征之一, 探讨建群种的空间格局是理解生物与生境之间关系和认识群落生态过程的有效途径。空间格局特征包括种群空间关联性及其空间分布型。利用无人机技术, 获取昆明海口林场半湿润常绿阔叶林和落叶栎类林的多光谱遥感影像, 采用ArcGIS Pro软件进行遥感影像分类并提取滇青冈()、光叶石栎()、滇石栎()、栓皮栎()、槲栎()植株树冠中心点空间坐标。在Programita Febrero 2014软件中进点格局分析。结果表明: 各物种10m以下呈均匀分布或随机, 随尺度增加, 渐渐表现为聚集分布。光叶石栎种群和滇石栎种群空间关联性表现为负相关。
半湿润常绿阔叶林; 多光谱影像; 种群格局; 点格局
0 前言
种群是理解植物群落结构与动态变化机制的关键[1], 其空间属性表现为种群个体在水平空间的配置状况或分布状况, 即种群格局(Population pattern)[1-9]。种群个体在水平空间上彼此间的相互关系是种内竞争、种间竞争及自然环境等各种因素共同作用下形成的结果[1, 4-5, 13]。种群空间分布格局反映该种群与生境的关系及其在群落中的地位和作用, 同时与尺度关系密切[2, 6, 7]。种群空间分布格局特征包括空间关联性、空间分布型和格局纹理, 种群空间关联有正关联、负关联、无关联三种方式, 种群格局通常分为随机分布、均匀分布、聚集分布三种类型[5]。
种群空间分布格局及其关联性对种群的生长、发育、资源利用等具有显著的影响[3-5, 16], 研究植物种群格局一方面有助于理解它们的生态过程, 另一方面可以揭示生物与生境的相互关系[1-9, 16]。建群种空间格局动态与尺度密切相关, 传统的样方取样和分析方法, 无论样方大小是多么的合理 , 都不能全面反映一个种在空间的分布特点和种间的相互关系[6]。点格局分析方法可显示任意尺度下的建群种格局变化, 该方法由Ripley 在1977 年提出, 它克服了传统方法只能分析单一尺度空间分布格局的缺点, 目前已被广泛应用于种群布格局的分析中并成为建群种空间格局研究的主要方法之一[5, 6-9]。点格局分析以植物个体的空间坐标为基础数据, 将每个个体视为二维空间的一个点, 所有个体组成空间分布的点图, 以点图为基础进行格局分析, 可探讨各种尺度下的种群格局特征和彼此间的关系[1, 4, 5, 20]。点格局分析法不仅能够提供全面的空间格局信息, 还可以最大限度地利用坐标图的信息, 具有较强的检验能力[5]。有人称点格局分析才是真正意义上的空间格局分析。
卫星遥感是景观和空间生态学研究的重要数据源, 但是早期的卫星遥感影像受分辨率和重访周期的限制(例如MODIS数据空间分辨率250 m; Landsat数据空间分辨30 m, 访问周期18 d), 难以满足研究对不同尺度的需求[12-13]。随着无人机技术的飞速发展, 无人机(Unmanned Aerial Vehicle, UAV)低空摄影测量和遥感(Photogrammetry and Remote Sensing, PaRS)技术由于具有拍摄影像分辨率高、重叠率大、姿态角大、相幅小、数量多等特点, 因此在许多科学应用方面具有巨大的发展潜力[12], 例如近地面无人机生态调查技术的应用, 可以获取种群个体的空间分布信息。随生态保护工作的推进, 经营性林区逐渐转变为生态公益林性质的林区, 昆明市海口林场宽地坝林区就是一个典型的例子。本文拟探讨①自然因素与人为因素影响下森林群落建群种的种群格局特征; ②利用近地面无人机技术获取植被林冠影像图, 进行建群种种群点格局分析, 与传统的研究方法所得结果进行比较, 探讨群落建群种空间格局和种间关联性特征, 空间格局的尺度效应, 为生态公益林生物多样性保护提供科学建议。
1 研究内容与方法
1.1 研究区概况
昆明海口林场成立于1956年, 现属昆明市林业局。昆明海口林场海拔1800 m—2400 m, 相对高差600 m[15]。海口林场地处昆明市西山区海口镇, 距离昆明市中心40 km, 地理位置为102°28′—102°38′ E、24°43′—24°56′ N之间。昆明海口林场气候温和, 冬无严寒, 夏无酷暑。林区总面积7563 hm2, 林业用地面积6683.8 hm2, 森林覆盖率达80.54%。降水分布不均, 年平均气温15.4 ℃, 年均日照时间2200 h, 属亚热带半湿润气候类型[14, 15, 17]。半湿润常绿林主要分布在海拔1700 m—2500 m, 是云南中部的基本植被类型之一。
海口林场植被类型非常丰富, 由半湿润常绿阔叶林的天然林和自然更新的次生林相间混杂。原生性植被为滇中地带性植被—半湿润常绿阔叶林, 在林区以天然林和次生林的形式存在, 主要树种包括滇青冈()、光叶石栎()、滇石栎()、高山栲()、锥连栎()等。人工植被包括华山松()林、干香柏()林、云南松()林、杉木()林等。
1.2 研究方法
1.2.1 野外调查
野外群落调查采用样方法进行, 设置28个20 m×20 m的样方, 在样方四个角设置5 m×5 m大小灌木样方及1 m×1 m大小草本样方。野外调查时记录所有样方的GPS、经度和纬度, 研究区域位置见图1, 样方信息见表1。根据野外记录的样方GPS以及海口林场Google earth可见光影像确定无人机飞行区域, 运用无人机(GV1300型, 数字绿土公司, 中国北京)遥感低空飞行, 飞行航高200m, 航线间距50m, 挂载Micasense RedEdge多光谱传感器(Micasense 美国)拍摄获取高光谱遥感影像, 并对获取的影像信息叠加并处理后进行正射矫正。影像空间分辨率0.1 m× 0.1 m, 坐标系选择WGS_1984_UTM_Zone_48N。
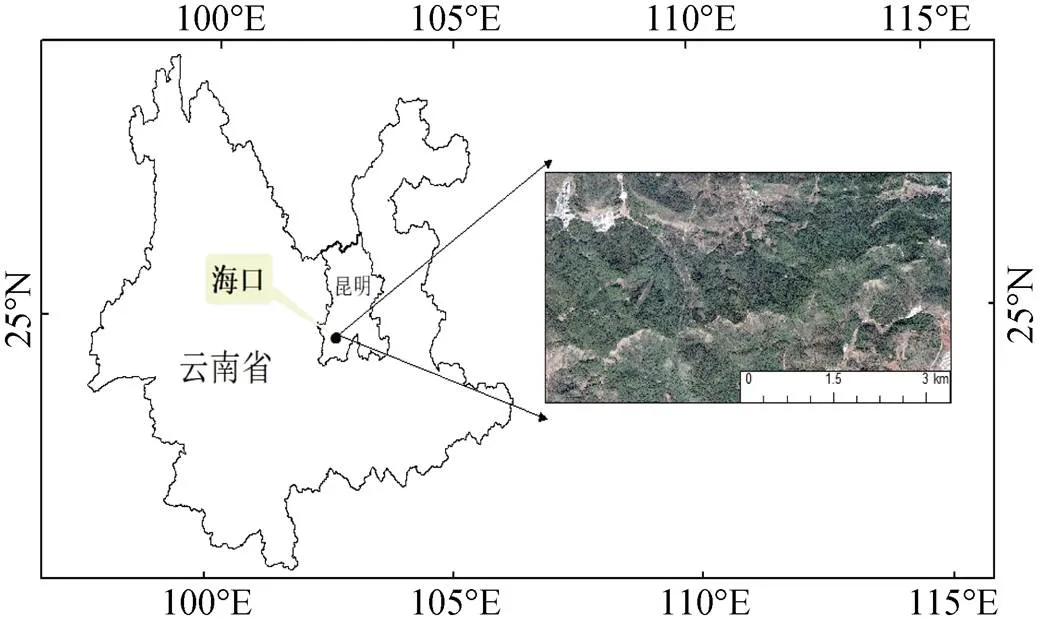
图1 研究区位置图
Figure 1 Location of the study area
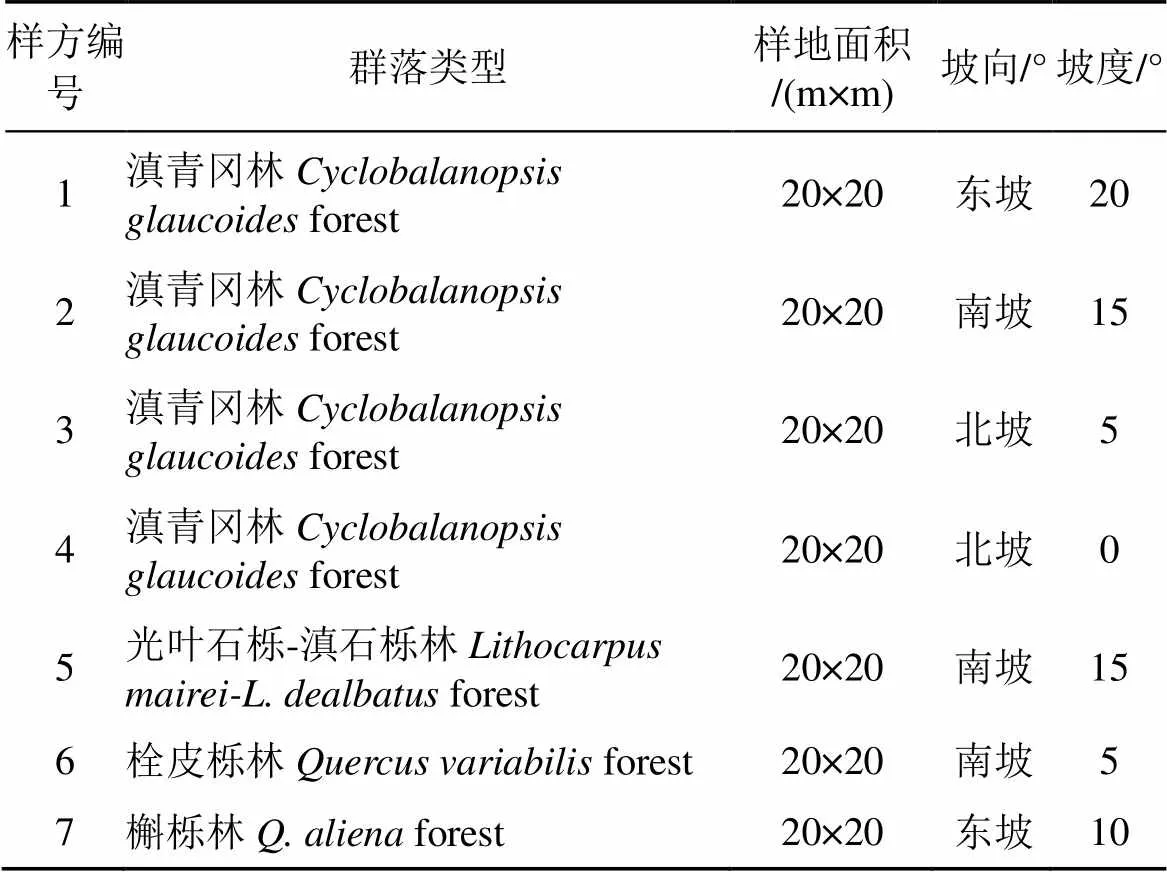
表1 群落样地调查信息表
1.2.2 遥感影像分类方法
利用Photoscan软件对无人机多光谱图像进行重叠合成。以野外调查样方GPS点为中心做100米的方形缓冲区, 提取缓冲区大小的无人机影像。根据野外调查数据建立群落建群种的训练区, 利用ArcGIS pro软件的支持向量机(Support vector machine, SVM)方法进行监督分类, 该方法具有较高的计算速度和较快的收敛速度[30], 并对分类图进行精度检验, 各物种分类结果总体精度为0.95。利用分类图提取建群种树冠中心点空间坐标进行格局分析。
1.2.3 格局分析方法
利用野外调查数据计算小尺度上样方内建群种聚集度大小指数(ICS):
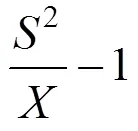
该指数也称之为聚集指数, 当ICS<0时为均匀分布; ICS=0时为随机分布; ICS>0时为聚集分布[3, 20]。
用分类图提取群落建群种植株树冠中心点空间坐标, 在Programita Febrero 2014软件中分析群落建群种空间格局和群落共建种之间的相关性。分析结果图的源代码导入RSstudio中查看, 并把数据导入Origin中作图, 以提高输出结果图片的分辨率。运用点格局分析法中的Ripley K函数, 对滇青冈、光叶石栎、滇石栎、栓皮栎和槲栎的空间分布格局进行分析。单变量K(t)函数公式如下:

式中:为测定面积;为测定区内植物个体总数;d为与之间的间距;表示空间尺度, 当d≤时,(d)=1, 当d>时,(d)=0;W是圆周长在面积中的比例。
为了使结果更可靠, 方差更稳定, 通常引入/的平方根对()函数进行了修正, 得出()函数, 公式如下:
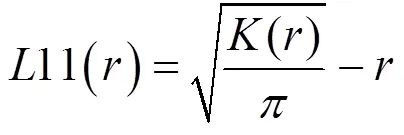
若11()=0, 则种群为随机分布; 若11()>0, 则种群为聚集分布; 若11()<0时, 则种群为均匀分布。采用Monte-Carlo模拟99%置信区间, 进行结果偏离随机状态的显著性检验。()值位于置信区间之上, 种群呈聚集分布;()值位于置信区间之下, 种群呈均匀分布;()值位于置信区间之内, 种群呈随机分布[1, 4-6, 8, 9, 11, 16]。
1.2.4 空间关联分析方法
空间关联分析可采用 Ripley K函数转换的双变量函数进行分析, 但为了结果更可靠, 方差更稳定, 修正后的函数为:

当12()>0时, 表示不同龄级个体在范围正关联; 当12()=0时, 不同龄级个体在范围没有显著相关性; 当12()<0时, 不同龄级个体在范围负关联[26, 31]。采用Monte-Carlo方法来模拟99%的置信区间, 得到上下两条包迹线。计算空间分布的上下包迹线, 从而形成()以及12()的置信区间[4, 5, 8, 9, 11, 16]。若实测12()值大于置信区间上限为显著正相关, 在置信区间内为不相关, 小于置信区间下限为显著负相关[1, 6]。
2 结果与分析
2.1 聚集指数
海口林场植物群落的优势种有滇青冈、光叶石栎、滇石栎、栓皮栎和槲栎。种群的空间分布格局计算结果如表2所示, 根据ICS值显示, 各优势种群在空间上呈现聚集分布, 但聚集程度不同。
2.2 点格局分析
群落优势种群的个体分布点图如图2所示, 种群个体分布点图差异较大, 种群分布格局与尺度密切相关。各群落优势种群的空间分布格局如图3所示。由图3可知: 滇青冈种群在研究尺度0—5 m范围内总体表现为均匀分布, 在尺度5—10 m表现为随机分布, 当>10 m时表现为聚集分布,=40 m时聚集程度达到最大,>40 m滇青冈聚集程度逐渐下降如图3(样地1—4)。光叶石栎种群和滇石栎种群在研究尺度0—5 m内表现为均匀分布, 光叶石栎在5 m以后表现为随机分布, 滇石栎在研究尺度5—8 m内表现为随机分布, 当>8时, 表现为聚集分布,=30时聚集程度最大, 随后聚集程度逐渐下降如图3(样地 5(a、b))。光叶石栎种群和滇石栎种群空间关联性分析12()<0, 两种群个体分布格局关联性为负关联如图3(样方编号5(c))。栓皮栎种群和槲栎在0—7 m内表现为均匀分布, 在研究尺度7—15 m内表现为随机分布,>15 m时表现为聚集分布,=45 m聚集程度最大, 随后聚集程度均有所下降(样地6, 7)。
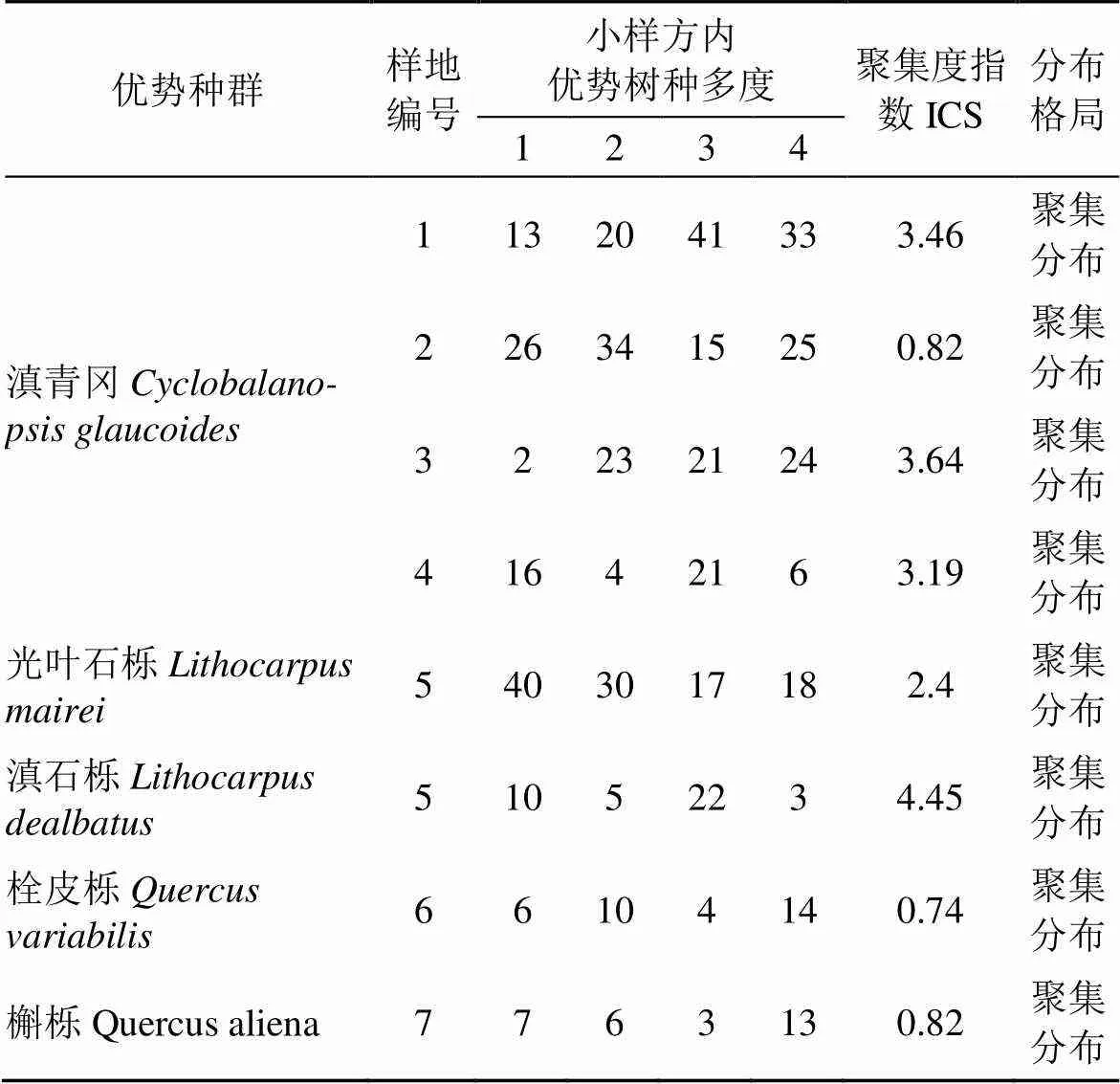
表2 优势种群空间分布格局聚集度指数测定结果
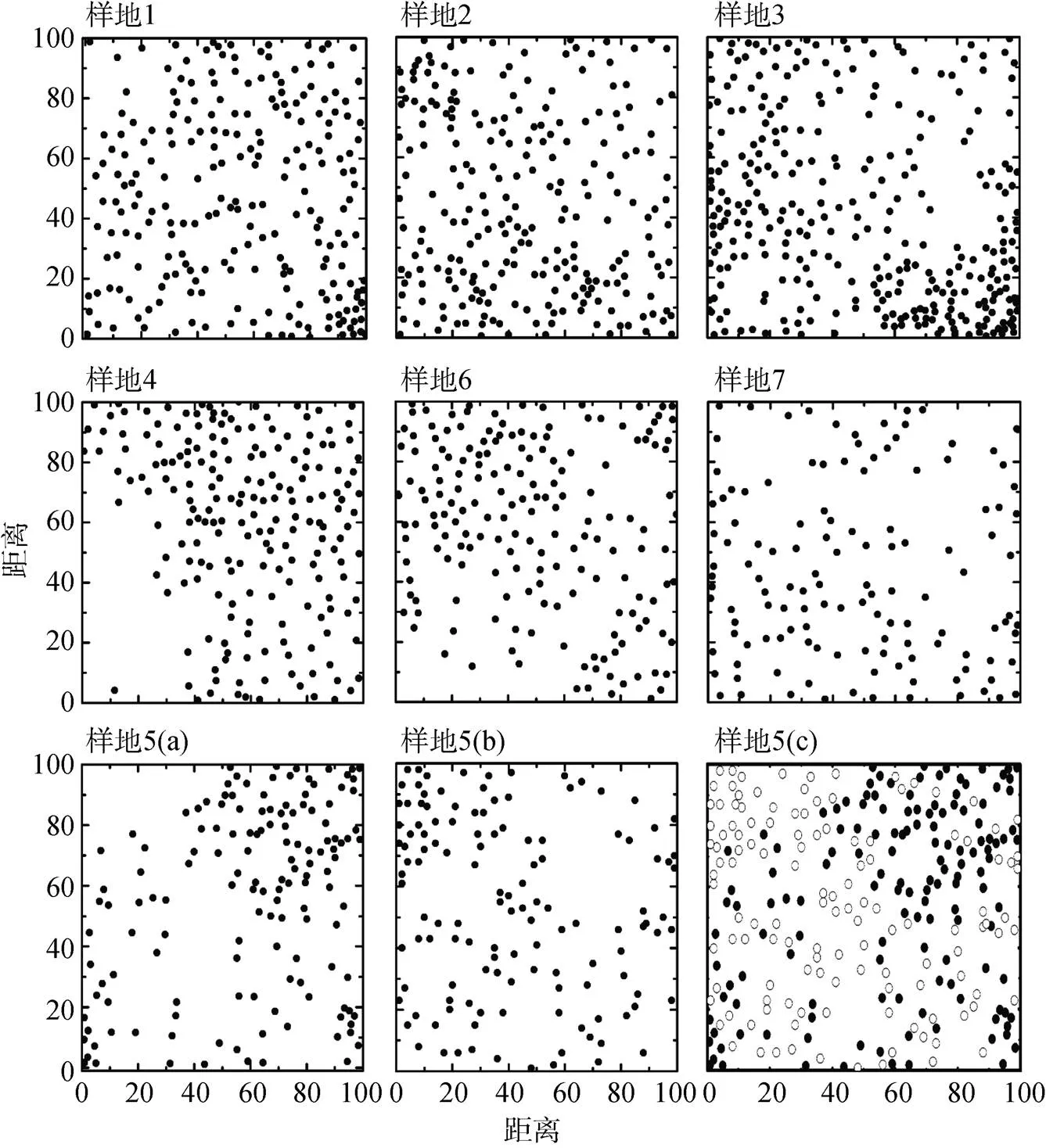
注:样地5(c): ○滇石栎; ●光叶石栎。
Figure 2 The point diagrams of individual distribution of the tree populations
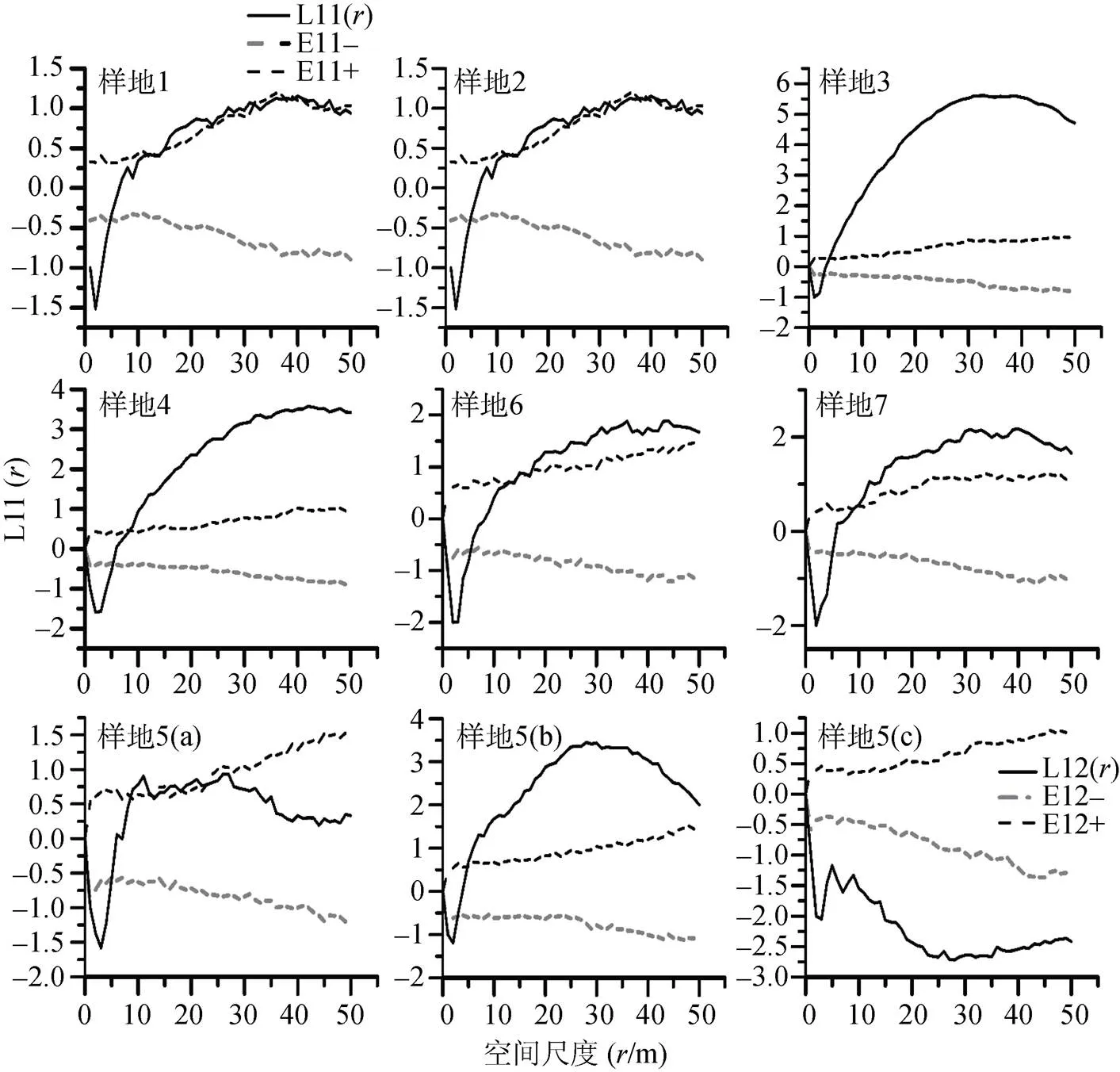
图3 各树种种群的点格局
Figure 3 The point pattern of the populations
3 讨论与结论
种群的空间分布格局与空间尺度密切相关, 空间分布格局随着尺度的改变而变化, 种群空间分布格局的这种尺度变异性受多因素制约, 如种内或种间竞争、生境的异质性和斑块性等[5]。野外20 m×20 m样方调查数据表明, 5种建群种的种群分布格局为聚集分布; 根据无人机获取的遥感影像图, 对群落建群种空间分布格局分析表明海口林区植物群落中建群种在10 m以下分布均匀, 群落建群种在细微尺度上呈均匀分布, 当空间尺度开始增大时, 种群空间分布格局呈现随机分布, 之后逐渐过渡为聚集分布。聚集指数可提供格局类型的信息, 点格局能进一步反映出格局的尺度效应信息。一般而言, 在粗尺度上物种往往呈聚集分布。对于大多数物种的分布格局来说, 寻找单一的限制因素和简单的解释可能是没有成效的。一个物种不仅可能受到其范围内不同因素的限制, 而且即使在一个局部地区, 几个因素也可能以复杂的方式相互作用。种群的空间分布格局通常是植物自身的生物学特性、生境异质性以及种内种间竞争等生物因素与非生物因素综合作用的结果[4, 5, 20, 24, 25]。植物繁殖特性、种子散布方式、竞争强度以及微地形、光照、土壤养分、水分等生境差异, 都可在不同尺度对某一物种的空间分布格局起作用[24, 25, 31]。关于群落中建群种格局形成的原因, 不同的群落有不同的形成原因。局域土壤和地形等立地条件差异, 影响不同树种聚集的程度。同时自然界中普遍存在干扰。无论是细尺度干扰还是粗尺度干扰, 对森林群落的自然更新都起着重要作用。昆明海口林场林区经多年当地森林管理局保护和管理, 以滇青冈、光叶石栎、滇石栎、栓皮栎、槲栎为乔木层优势种的群落组成和结构相对单一, 小尺度上树种呈均匀分布, 随着距离增大, 各种群由随机分布逐渐过渡为聚集分布, 符合天然林自然分布特征。
各建群种的种群格局特征表现不一。滇青冈种群在尺度10 m以下表现为均匀或随机分布, 10 m以上呈聚集分布, 40 m后聚集程度逐渐下降。光叶石栎种群和滇石栎种群在8 m以下表现为均匀或随机分布, 8 m以上表现为聚集分布, 30 m后聚集程度逐渐下降。栓皮栎和槲栎在尺度15 m以下表现为均匀或随机分布, 15 m后表现为聚集分布, 45 m后聚集程度逐渐下降。落叶树种的格局纹理规模更高。吴初平[32]在研究以青冈为建群种的常绿阔叶林时发现, 生境异质性在群落格局形成中起到重要作用。他还认为青冈栎的种子粒大、质量较重, 野外调查发现海口林场林区内分布有坡度较大的青冈生境, 青冈栎种子在掉落过程中可能会滚落至山脚或平缓处, 从而形成强烈的聚集分布[32]。
没有一种生物是独立, 以至于它的分布不受其他物种的影响。刘振国[33]等研究表明, 空间正关联和负关联主要发生在细尺度下, 空间关联性随尺度的增大而减弱, 并趋于无关联。种间关联性认为, 一个物种对另一个物种存在依赖或有共同的环境需求, 则它们呈正相互关系; 负相互关系是由于两物种在对资源竞争过程中相互排斥, 或者不相似的环境需求[2,4]。细尺度上光叶石栎和滇石栎种群空间关联性表现为负相关。主要是各树种细尺度上存在竞争, 为竞争环境资源而产生负相关性, 而随着尺度的增大, 大径阶的个体数量越来越少, 个体之间距离也越来越远, 因此, 大部分树种在大尺度上表现为不相关。在这一研究结果体现出了种群空间格局的尺度依赖性, 说明同一个群落内种群内或不同个体之间的相互作用具有显著的尺度效应, 当大于某一个尺度时, 个体间的相互作用将明显减弱。
昆明海口林场林区内以常绿阔叶林为植被主体, 华山松和云南松等针叶林镶嵌其中。研究的5个群落的建群种空间格局虽然聚集尺度存在一定的差异, 但总体上均呈现细尺度均匀或随机分布, 随着尺度的增大, 各建群种趋向于从均匀或随机分布转变为聚集分布, 之后聚集程度逐渐下降。光叶石栎种群和滇石栎种群之间的空间格局动态呈现负相关性, 负相关性表明两种树种在空间上相互排斥。因此, 植被恢复中应以植物群落优势种间的空间关联性为依据进行合理配置, 减少种间竞争, 增加生物多样性, 从而更好地发挥森林的生态作用。
多尺度下空间格局分析是点格局分析方法的优势, 它不仅能够直观地表现出空间格局随尺度变化的分布情况, 还可对野外受尺度限制的调查进行补充, 而无人机技术提供了这种多尺度格局分析的一种新的平台。
[1] 杨乃坤, 邹天才, 刘海燕, 等. 贵州特有植物长柱红山茶种群年龄结构及空间分布格局研究[J]. 热带亚热带植物学报, 2015, 23(2): 205–210.
[2] HUI Gangying, LI Li, ZHAO Zhonghua, et al. The comparison of methods in analysis of the tree spatial distribution pattern[J]. Acta Ecologica Sinica, 2007, 27(11): 4717–4728.
[3] 钱军, 邱明红, 杜尚嘉, 等. 五指山市金钟藤种群分布格局分析[J]. 生态科学, 2016, 35(4): 97–101.
[4] 刘旻霞, 连依明, 李文. 微地形对优势种群点格局和关联性的影响[J]. 应用生态学报, 2018, 29(5): 1569–1575.
[5] 樊登星, 余新晓. 北京山区栓皮栎林优势种群点格局分析[J]. 生态学报, 2016, 36(2): 318–325.
[6] 张金屯. 植物种群空间分布的点格局分析[J]. 植物生态学报, 1998, 22(4): 344–349.
[7] 马志波, 肖文发, 黄清麟, 等. 生态学中的点格局研究概况及其在国内的应用[J]. 生态学报, 2017, 37(19): 6624– 6632.
[8] HAASE P. Spatial pattern analysis in ecology based on Ripley’s K-function - introduction and methods of edge correction[J]. Journal of Vegetation Science, 1995, 6(4): 575– 582.
[9] 刘旻霞, 夏素娟, 南笑宁, 等. 基于Ripley’s K函数的荒漠锦鸡儿种群分布格局[J/OL]. 干旱区研究, 2019, 36(3): 606–613.
[10] DIGGLE P J. Statistical analysis of spatial point patterns[M]. 2. NY, US: Hodder Education Publishers, 1983: 52–60.
[11] 王鑫厅, 侯亚丽, 梁存柱, 等. 基于不同零模型的点格局分析[J]. 生物多样性, 2012, 20(02): 151–158.
[12] 张志明, 徐倩, 王彬, 等. 无人机遥感技术在景观生态学中的应用. 生态学报, 2017, 37(12): 4029–4036.
[13] 郭程轩, 徐颂军, 袁中友. 3S 与模型方法在湿地景观格局研究中的应用述评[J]. 生态科学, 2007, 26(3): 250–255.
[14] 李志敏, 孙航. 滇中半湿润常绿阔叶林主要优势及常见种叶形态结构特征[J]. 云南植物研究, 1998, 20(1): 47– 48.
[15] 李小双, 宋亮, 陈军文, 等. 昆明西山半湿润常绿阔叶林及其次生演替群落的更新特征[J]. 广西植物, 2012, 32(4): 475–482.
[16] 张建亮, 李先琨, 韩文衡, 等. 广西木论自然保护区掌叶木种群结构与分布格局[J]. 生态科学, 2009, 28(6): 495– 502.
[17] 张如松, 吴宁, 安科, 等. 滇中半湿润常绿阔叶林群落类型演替的植物多样性调查与保护[J]. 林业调查规划, 2018, 43(1): 34–38.
[18] DRUCKENBROD D L, SHUGART H H, DAVIES I. Spatial pattern and process in forest stands within the Virginia piedmont[J]. Journal of Vegetation Science, 2005, 16(1): 37–48.
[19] Leps Jan, KINDLMANN P. Models of the development of spatial pattern of an even-aged plant population over time[J]. Ecological Modelling, 1987, 39(1/2): 45–57.
[20] 邓东周, 贺丽, 鄢武先, 等. 川西北高寒区不同沙化类型草地优势种群空间格局分析[J]. 草地学报, 2017, 25(3): 492–497.
[21] 董灵波, 刘兆刚, 张博, 等. 基于Ripley L和O-ring函数的森林景观空间分布格局及其关联性[J]. 应用生态学报, 2014, 25(12): 3429–3436.
[22] DAVIS J H, HOWE R W, DAVIS G J. A multi-scale spatial analysis method for point data[J]. Landscape Ecology. 2000, 15(99): 99–114.
[23] Tomáš M, SAMUEL S, Mari M, et al. Monte Carlo testing in spatial statistics, with applications to spatial residuals[J]. Spatial Statistics, 2016, 18(A): 40–53.
[24] 王道亮, 李卫忠, 曹铸, 等. 黄龙山天然次生林辽东栎种群空间格局[J]. 生态学报, 2016, 36(09): 2677–2685.
[25] 梅军林, 庄枫红, 马姜明, 等. 桂林喀斯特地区克隆生长红背山麻杆种群的点格局分析[J]. 生态学报, 2017, 37(9): 3164–3171.
[26] LAN Guoyu, GETZIN S, WIEGAND T, et al. Spatial distribution and interspecific associations of tree species in a tropical seasonal rain forest of China[J]. Plos One, 2012, 7(9): e46074.
[27] HOU J H, MI X C, LIU C R, et al. Spatial patterns and associations in a-forest in northern China[J]. Journal of Vegetation Science, 2004, 15(3): 407–414.
[28] FRANK M S, OLIVER B, SUZANNE J M, et al. Spatial pattern formation in semi-arid shrubland: a priori predicted versus observed pattern characteristics[J]. Plant Ecology, 2004, 173(2): 271–282.
[29] ATKINSON P M, FOODY G M, GETHING P W, et al. Investigating spatial structure in specific tree species in ancient semi-natural woodland using remote sensing and marked point pattern analysis[J]. Ecography, 2007, 30(1): 88–104.
[30] 孟梦, 田海峰, 邬明权, 等. 基于Google Earth Engine平台的湿地景观空间格局演变分析: 以白洋淀为例[J]. 云南大学学报(自然科学版), 2019, 41(2): 416–424.
[31] LAN Guoyu, HU Yuehua, GAO Min, et al. Topography related spatial distribution of dominant tree species in a tropical seasonal rain forest in China[J]. Forest Ecology and Management, 2011, 262(8): 1507–1513.
[32] 吴初平, 袁位高, 盛卫星, 等. 浙江省典型天然次生林主要树种空间分布格局及其关联性[J]. 生态学报, 2018, 38(2): 537–549.
[33] 刘振国, 李镇清. 植物群落中物种小尺度空间结构研究. 植物生态学报, 2005, 29(6): 1020–1028.
Research on population pattern of dominate tree species in plant communities based on UAV at the Kuandiba section of Haikou Forest Farm in Kunming
Li Fuli, Peng Xiaochang, Wang Chongyun*, Peng Mingchun, Xie Yingxu, Zuo Yanjie
Institute of Ecology and Geobotany, Yunnan University, Kunming 650091, China
Constructive species are the dominant species of plant communities and have a significant influence on the community environment. Spatial pattern is one of the basic features of plant populations. Exploring the spatial pattern of constructive species is an effective way to understand the relationship between organisms and habitats and the ecological processes of communities. The characteristics of spatial pattern include spatial correlation and spatial distribution of populations. In this study, Unmanned Aerial Vehicle (UAV) technology was used to obtain the multi-spectral remote sensing images of the semi-humid evergreen broad-leaved forest and deciduous oak forest in Kunming Haikou forest farm. ArcGIS Pro software was used to interpret the remote sensing images and to derive the center coordinates of forest trees’ crown of,,,andspecies. Spatial pattern analysis was conducted by Programita Febrero 2014 software. The results showed that the tree species were uniformly distributed at small scales (<10m), and gradually showed aggregation distribution with the increasing of spatial scales. Among tree species,population andpopulation had a negative spatial correlation.
semi-humid evergreen broad-leaved forest; multi-spectral remote sensing images; population pattern; pointpattern
10.14108/j.cnki.1008-8873.2020.03.009
Q948
A
1008-8873(2020)03-057-07
2019-05-05;
2019-08-14
云南大学一流大学建设项目“高原山地-城市-湖泊复合生态系统研究站”; 云南省科技厅-云南大学“双一流”重大建设项目“《云南植被志》研编”。
李富利(1994—), 女, 硕士, 主要从事景观植被及遥感和地理信息系统应用研究, E-mail: 450075805@qq.com
王崇云(1971—), 副教授, E-mail: cywang@ynu.edu.cn
李富利, 彭晓昶, 王崇云, 等. 基于无人机的昆明海口林场宽地坝林区植物群落建群种格局研究[J]. 生态科学, 2020, 39(3): 57–63.
Li Fuli, Peng Xiaochang, Wang Chongyun, et al. Research on population pattern of dominate tree species in plant communities based on UAV at the Kuandiba section of Haikou Forest Farm in Kunming[J]. Ecological Science, 2020, 39(3): 57–63.
猜你喜欢
好孩子画报(2020年11期)2020-12-21
今日农业(2020年15期)2020-12-15
绿色中国(2019年17期)2019-11-26
今日农业(2019年13期)2019-08-12
人民调解(2019年5期)2019-03-17
现代装饰(2017年12期)2017-05-22
中成药(2017年3期)2017-05-17
中国环境监察(2016年12期)2016-10-24
中国卫生标准管理(2015年6期)2016-01-14
中国铸造装备与技术(2015年5期)2015-12-10
