
光照强度对8种红树植物幼苗生长和生物量分配的影响
2020-06-08 12:09谭淑娟李婷1余素睿蔡水花1叶万辉1沈浩1
生态科学 2020年3期
谭淑娟, 李婷1,2,, 余素睿, 蔡水花1,2, 叶万辉1, 沈浩1,*
光照强度对8种红树植物幼苗生长和生物量分配的影响
谭淑娟1,2, 李婷1,2,5, 余素睿4, 蔡水花1,2, 叶万辉1,3, 沈浩1,3,*
1. 中国科学院退化生态系统植被恢复与管理重点实验室/广东省应用植物学重点实验室, 中国科学院华南植物园, 广州 510650 2. 中国科学院大学, 北京 100049 3. 南方海洋科学与工程广东省实验室(广州), 广州 511458 4. 仲恺农业工程学院园艺园林学院, 广州 510225 5. 中国水产科学研究院南海水产研究所, 广州 510300
选取无瓣海桑(Buch-Ham.)、卤蕨(Linn.)、桐花树(Blanco.)、老鼠簕(Linn.)、木榄(Lam.)、秋茄(Druce.)、银叶树(Ait.)和黄槿(L.)为研究对象, 通过遮荫控制实验, 研究这8种红树植物幼苗在不同光照强度(自然光强的100%、45%、30%、10%)下的生长和生物量分配特征。结果表明: (1)遮荫对无瓣海桑幼苗相对生长速率、生物量积累、基径的增长和叶片数的增长均有显著的抑制作用; (2)桐花和老鼠簕在遮荫处理下生长状况较好, 遮荫对桐花生长的影响不显著, 对老鼠簕幼苗的生长则有一定的促进作用; (3)当光照强度降低时, 无瓣海桑、秋茄、老鼠簕、桐花树的幼苗都对地上部分投入了更多的生物量, 其中秋茄、老鼠簕和桐花树通过提高叶生物量来获取更大的光能捕获能力。研究结果表明, 从对光照的生长适应角度来看, 桐花、老鼠簕和卤蕨适宜种植在郁闭度较高的林下; 秋茄适宜在郁闭度较低的林下种植; 真红树无瓣海桑和木榄以及半红树黄槿和银叶树则不适合在林下种植。
红树植物; 光照强度; 相对生长速率; 生物量; 生物量分配
0 前言
红树林是指生长在热带、亚热带低能海岸潮间带上部, 受周期性潮水浸淹, 以红树植物为主体的常绿灌木或乔木组成的潮滩湿地木本生物群落[1]。红树林对热带、亚热带海湾生态系统的维持与发展起到关键性作用[2]。然而, 红树林目前面临着极大的威胁, 全球35%的红树林已经消失, 而我国红树林的总面积从20世纪50年代到21世纪初减少近60%[3]。随着人们对红树林生态价值认识的进一步加深, 全球开始开展红树林的生态恢复研究[4]。
目前, 红树植物生理生态的适应性研究主要集中在温度、盐度、潮汐等单个环境因子对红树植物生长影响方面[5-9], 而红树植物对光照响应的研究相对较少。植物对光的捕获和利用能力是其能否适应环境并在种间竞争中获胜的决定性因素[10]。特别在森林生态系统中, 光照被认为是最重要的单一限制性资源[11]。森林植被幼苗在郁闭林低光环境下会展现出种间差异[12-13]。植物体通常通过改变生物量分配和形态的变化, 即通常所说的形态可塑性来适应光照强度的改变。已有研究表明, 低光照导致红树林植物的光合速率下降、植物生长激素分泌增加, 从而致使植物个体的茎高增加、生物量下降、相对生长率降低[14-15]。同时, Smith和Lee指出白骨壤()在林外比在林内生长得更好[16]; 叶勇等研究表明生长在林外的秋茄幼苗比林内生长更好[17]; 廖宝文等发现无瓣海桑在光线较弱的林下层生长状况不佳[18]。以上红树植物幼苗的生长发育对光照水平的响应结果说明光照条件的减弱对天然红树林幼林的生长会有影响, 但是不同红树植物对光照强度的响应策略的差异有待进一步深入研究。
我国红树林中80%为退化的次生灌丛, 经由自然演替形成健康完善的红树林生态系统的难度极大[19]。近年来我国华南沿海大范围开展红树造林, 选用无瓣海桑等速生种进行红树林恢复。占总面积10.5%的无瓣海桑人工林的演替趋势很大程度上决定了我国红树林面积、林分多样性、生态效益和社会效益。这种高大、生长迅速的树种降低了林下环境的光照水平, 必然会对林下的本土红树树种的生长造成一定影响。同时, 在人为和自然因素的双重作用下, 我国的红树植物群落结构日趋简单, 由原先的混交林向树种单一的纯林退化, 导致红树林生态系统日益脆弱, 抵御外界干扰的能力降低。因此, 红树林群落结构亟待进一步改造和优化。针对以上问题, 本研究比较了8种红树植物的生长和生物量分配对不同光照条件的适应性差异, 旨在为人工红树林群落的优化配置和合理的林分改造提供理论与实践指导, 进一步促进人工红树林从植被恢复向功能恢复过渡。
1 材料与方法
1.1 试验区和种源区概况
遮荫试验在位于广东省广州市的中国科学院华南植物园大型实验生态学综合试验场进行。地理位置为23°10´42.79" N, 113°21´25.28"E, 海拔40 m。本地区属南亚热带海洋季风气候类型, 全年平均气温20—22 ℃, 平均相对湿度77%, 年降雨量为1982.7 mm。
购置实验材料的苗圃位于广东省珠海市的淇澳岛红树林自然保护区附近。其地理位置为22°23′40"—22°27′38" N, 113°36′40″—113°39′15″ E, 属南亚热带海洋性季风气候。区内年均温为22.2 ℃, 年降水量为1875.7 mm[20]。保护区受降雨、江河径流和潮汐的影响, 海水盐度在3.31‰—7.05‰之间变化, 海域潮汐属不正规半日潮。
1.2 实验材料
在珠海淇澳岛红树林自然保护区附近的苗圃内挑选苗龄1年、长势均一的8种红树植物的幼苗(见表1)。其中, 无瓣海桑为外来红树植物, 原产孟加拉国, 对我国沿海环境表现出很强的生态适应性, 在珠海淇澳岛已有成规模的造林。其他7种红树植物均为本土种, 包括5种真红树(秋茄、桐花、木榄、老鼠簕和卤蕨)和2种半红树植物(银叶树和黄槿)。8种红树植物幼苗的初始参数如表1所示。
1.3 实验设计
实验共设置4个光强梯度, 分别为相对自然光强的100%(T0对照)、45%(T1)、30% (T2)和10% (T3)[21]。实验通过不同透光度的黑色尼龙网获得不同光强(T1, T2, T3)。在遮荫实验初期, 选择全晴天, 用Li-250A照度计 (Li-cor, Inc, USA), 每天记录上午9:00、中午12:00和下午3:00各遮荫棚内和全光照处理下的光量子通量密度(Photosynthetic photon flux density), 连续测定3天, 4个处理的相对全日照光强实测值如表2所示。
将定植好的幼苗标号并分别置于4个处理区中, 进行遮荫处理1年。栽培基质使用珠海淇澳岛海泥, 其盐度、含水率, 全氮含量和有机碳含量分别为8.3‰、42.9%、1.61%和1.85%。人工海水以粗海盐和自来水调配而成, 含盐量模拟淇澳岛近岸海水平均盐度, 约为6‰。试验期间监测盐度, 适时补充人工海水。
1.4 生长测定
遮荫实验开始后, 每个月记录每个处理下的红树幼苗的茎高、基径、叶片数、分枝数、冠幅等植物生长指标。生物量和相对生长速率在培养结束时采用收获法测定。求出如下参数: 根生物量比(Root mass ratio, RMR, 根重/植株总重); 茎生物量比(Stem mass ratio, SMR , 茎重/植株总重); 叶生物量比(Leaf mass ratio, LMR , 叶重/植株总重); 地上部分与地下部分生物量比率(shoot/root dry weight ratio, S/R)=(茎生物量+叶生物量)/根生物量。相对生长速率(relative growth rate, RGR, g·g-1·d-1)

式中, 以遮荫后0 d(时间为0=0)红树幼苗的总生物量(W)作为初始数值;指收获的序次;W是t时期植株的总生物量。其中, 初始生物量(W)通过植物生长的对数模型, 既单株生物量()与基径()和茎高(H)的函数关系拟合估算。
lg= alg (D H)+b (1-2)
式中, a, b为拟合系数, 不同红树树种拟合系数和相关系数如表3所示。
1.5 数据分析
相关数据采用EXCEL 2013、SPSS 13.0软件进行统计分析, 当<0.05时表示有统计学意义。各处理间生长指标的差异使用单因素方差分析(One-way ANOVA)方法进行显著性检验, 当差异显著时以Tukeycomparisons法进行多重比较。
2 结果与分析
2.1 光强对8种红树幼苗成活率的影响
由表4可知, 各树种幼苗成活率均在60%以上, 其中无瓣海桑幼苗成活率最低(66%), 老鼠簕、桐花树和卤蕨幼苗成活率较高均在90%以上。在30%遮荫处理下, 7种本土红树植物幼苗的存活率都达到70%以上。无瓣海桑和木榄幼苗成活率均随相对光照强度的下降而降低, 在10%光照处理下, 除无瓣海桑和木榄幼苗成活率低于50%, 而其他红树植物幼苗存活率仍都在70%以上。

表1 8种红树植物及幼苗生长参数初始值(平均值±标准误, n=5)
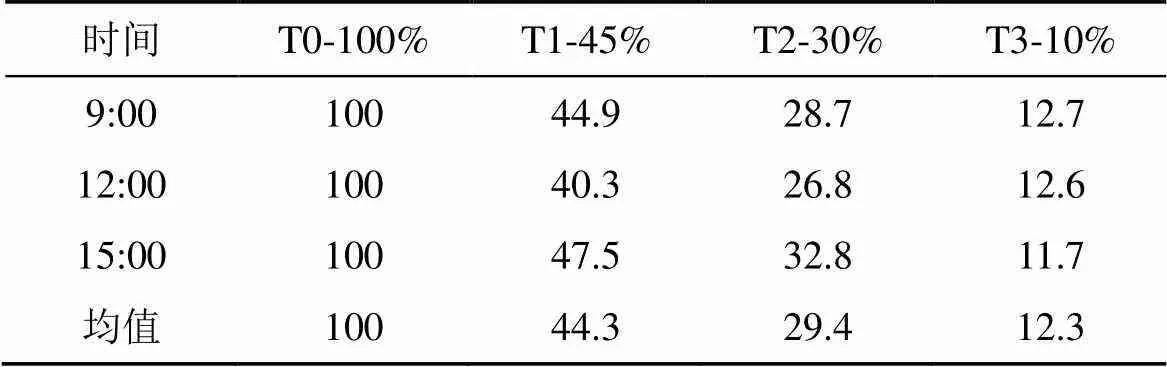
表2 各遮荫处理相对光强实测值

表3 8种红树幼苗初始生物量拟合曲线
注: 物种缩写见表1。
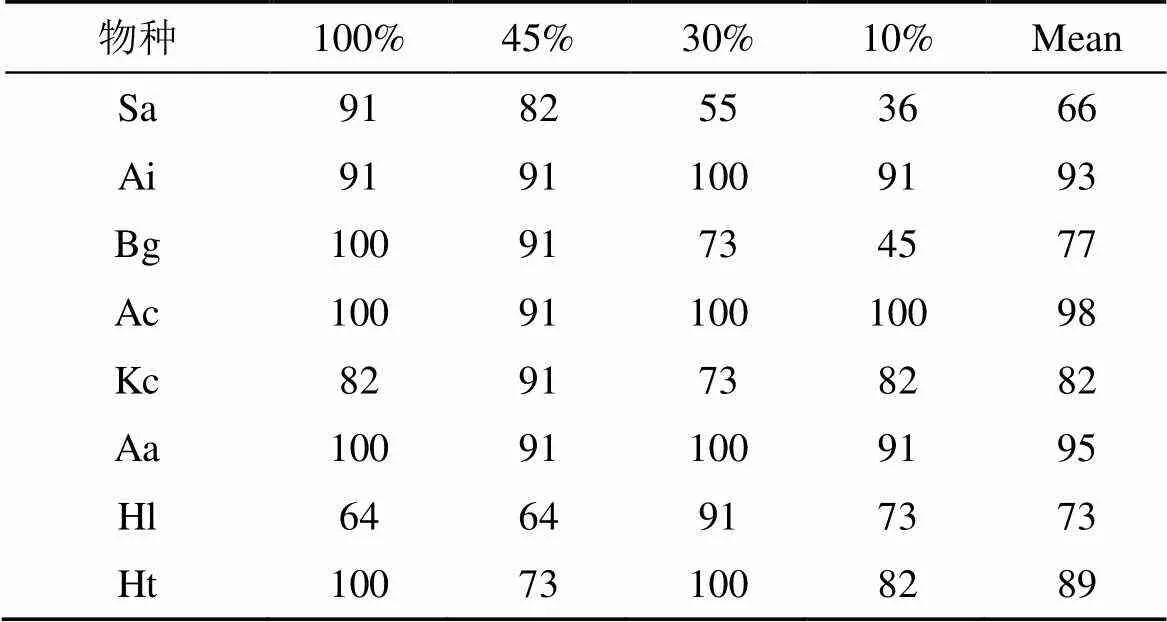
表4 8种红树幼苗成活率
注: 物种缩写见表1。
2.2 光强对8种红树幼苗茎高的影响
各红树幼苗在不同遮荫处理之间茎高增长量差异明显, 在全光照处理下, 无瓣海桑和黄槿幼苗苗高的增长较快, 增长量大于3个遮荫处理, 与之相反, 老鼠簕和桐花树幼苗在遮荫处理下, 苗高增长速率较快。遮荫对老鼠簕幼苗茎高增长有显著的促进作用, 对无瓣海桑幼苗茎高增长有显著的抑制作用。
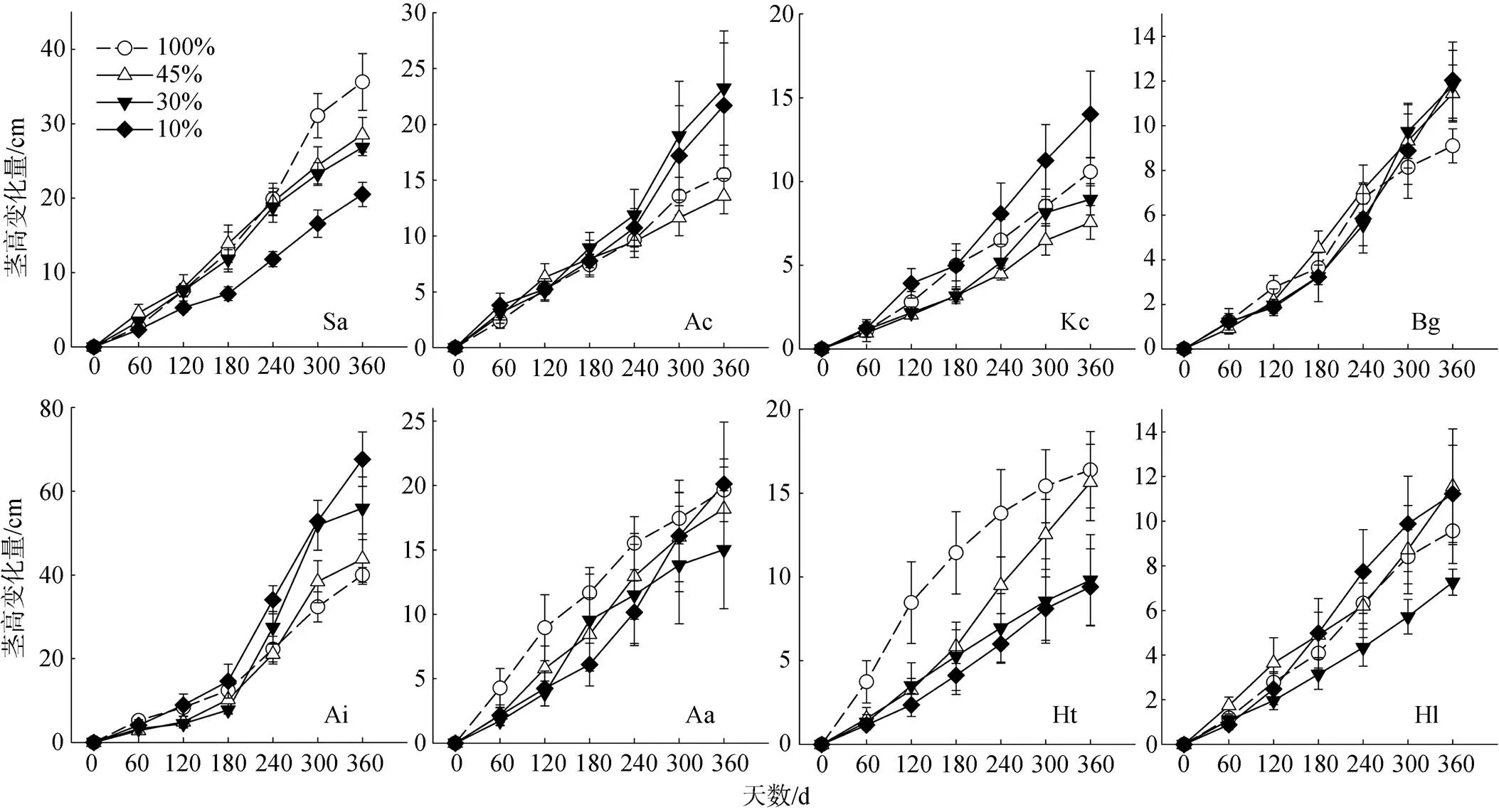
注: 物种缩写见表1。
Figure 1 Stem height growth dynamics of seedlings of the eight mangrove species under four light treatments
2.3 光强对8种红树幼苗基径和冠幅的影响
遮荫对秋茄、无瓣海桑和银叶树幼苗基径的增长量有显著影响, 对黄槿、卤蕨、老鼠簕、木榄、桐花树幼苗基径的增长量影响则不显著; 遮荫仅对老鼠簕和秋茄幼苗冠幅的增长量有显著影响, 对黄槿、卤蕨、木榄、桐花树、无瓣海桑和银叶树幼苗冠幅的增长量均未产生显著影响。在遮荫处理下, 无瓣海桑幼苗基径的年增长量在遮荫处理下显著低于全光照处(图2-A); 秋茄和老鼠簕幼苗冠幅的增长显著高于全光照处理, 其中老鼠簕冠幅增长在10%光照处理下达到最大值(图2-B)。
2.4 光强对8种红树幼苗叶片数和分枝数的影响
遮荫对卤蕨和无瓣海桑幼苗叶片数量变化及卤蕨和老鼠簕幼苗分枝数变化产生显著影响, 对其他6种红树植物幼苗叶片数量变化和分枝数变化影响不显著。无瓣海桑、卤蕨幼苗叶片数随光照强度的下降而显著下降, 其中无瓣海桑幼苗叶片数在4个处理下较处理前数量下降(图2-C); 老鼠簕和卤蕨在遮荫处理下幼苗的分枝数量显著少于全光照处理下的幼苗分枝数量, 其中卤蕨幼苗分枝数在45%和10%光照处理下较处理前减少(图2-D)。
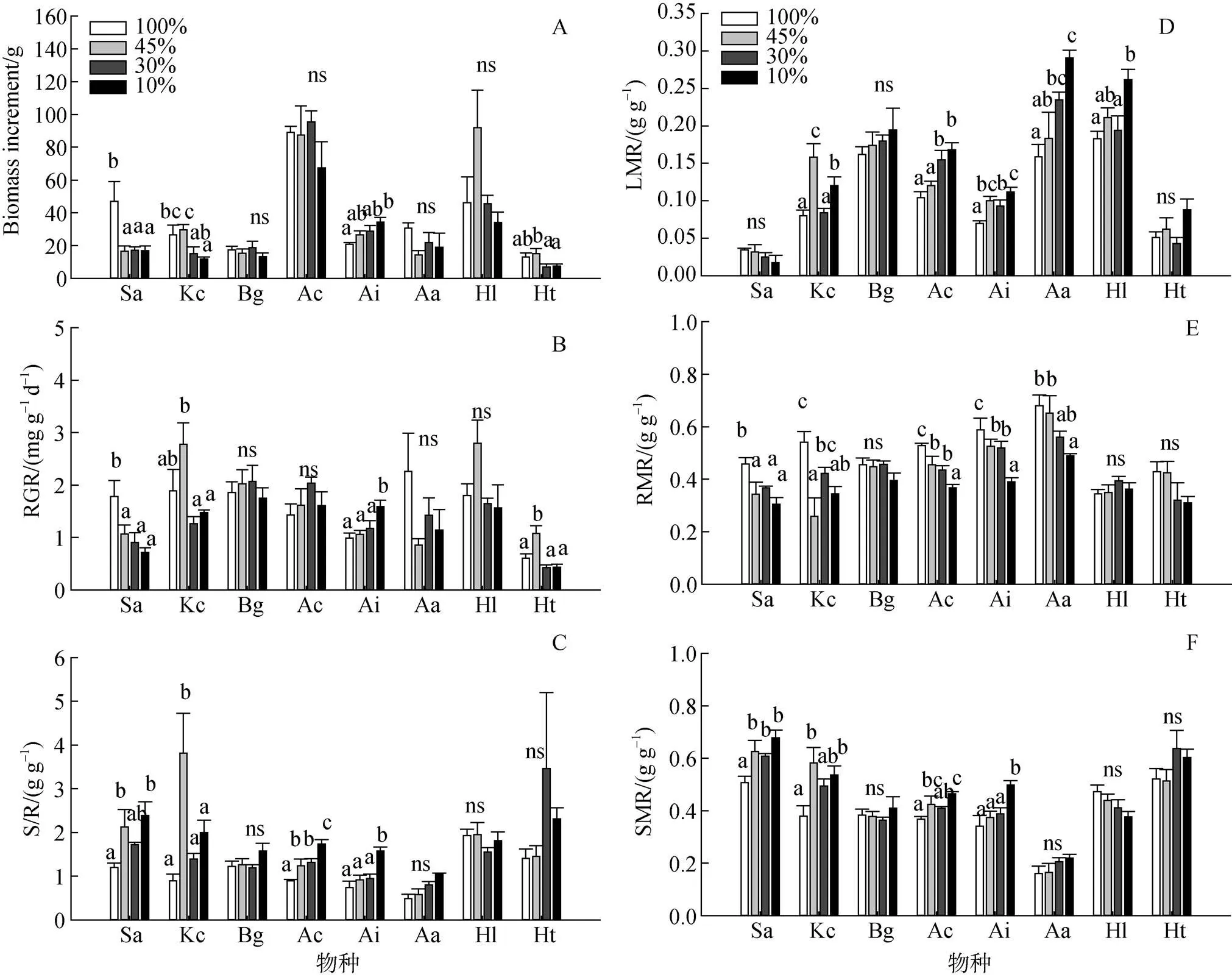
2.5 光强对8种红树幼苗相对生长速率和生物量变化及分配的影响
单因素方差分析结果(图3)表明: 遮荫对黄槿、老鼠簕、秋茄、无瓣海桑幼苗的生物量增长和RGR有显著影响; 遮荫对老鼠簕、秋茄、桐花树幼苗的S/R、LMR、RMR、SMR均有显著影响, 对无瓣海桑幼苗的S/R、LMR和RMR、卤蕨幼苗的LMR和RMR以及银叶树幼苗的LMR有显著影响。无瓣海桑幼苗生物量的积累和相对生长速率在遮荫处理下均有显著下降(图3-A)、与之相反老鼠簕幼苗生物量增长和相对生长速率在随光照强度的下降而增加, 在10%光照处理下达到最大值, 秋茄和黄槿幼苗生物量增长和相对生长速率则在45%光照处理下达到最大值(图3-B); 在遮荫处理下, 无瓣海桑、秋茄、桐花树和老鼠簕幼苗S/R值均有显著上升, 其中无瓣海桑幼苗S/R值在45%和10%光照处理下显著高于全光照处理组, 秋茄幼苗S/R值在45%光照处理下显著高于其他处理组, 老鼠簕幼苗S/R值在10%光照处理下显著高于其他处理组, 桐花幼苗S/R值随光照强度的下降显著升高(图3-C), 这意味着遮荫导致各幼苗地上部分生物量比重有不同程度的升高。
无瓣海桑幼苗在遮荫处理下, 茎生物量比显著升高, 根生物量比显著下降, 在3个遮荫处理间则不存在显著差异, 其叶生物量比相对其他红树植物较低; 秋茄、桐花树和老鼠簕幼苗在遮荫处理下茎和叶的生物量比重均有所增加, 根生物量比显著下降; 卤蕨幼苗在遮荫处理下叶生物量比上升, 根生物量比下降; 木榄、银叶树和黄槿各部分生物量分配在各处理间变化不大, 受遮荫的影响程度较小(图3-D、3-E和3-F)。
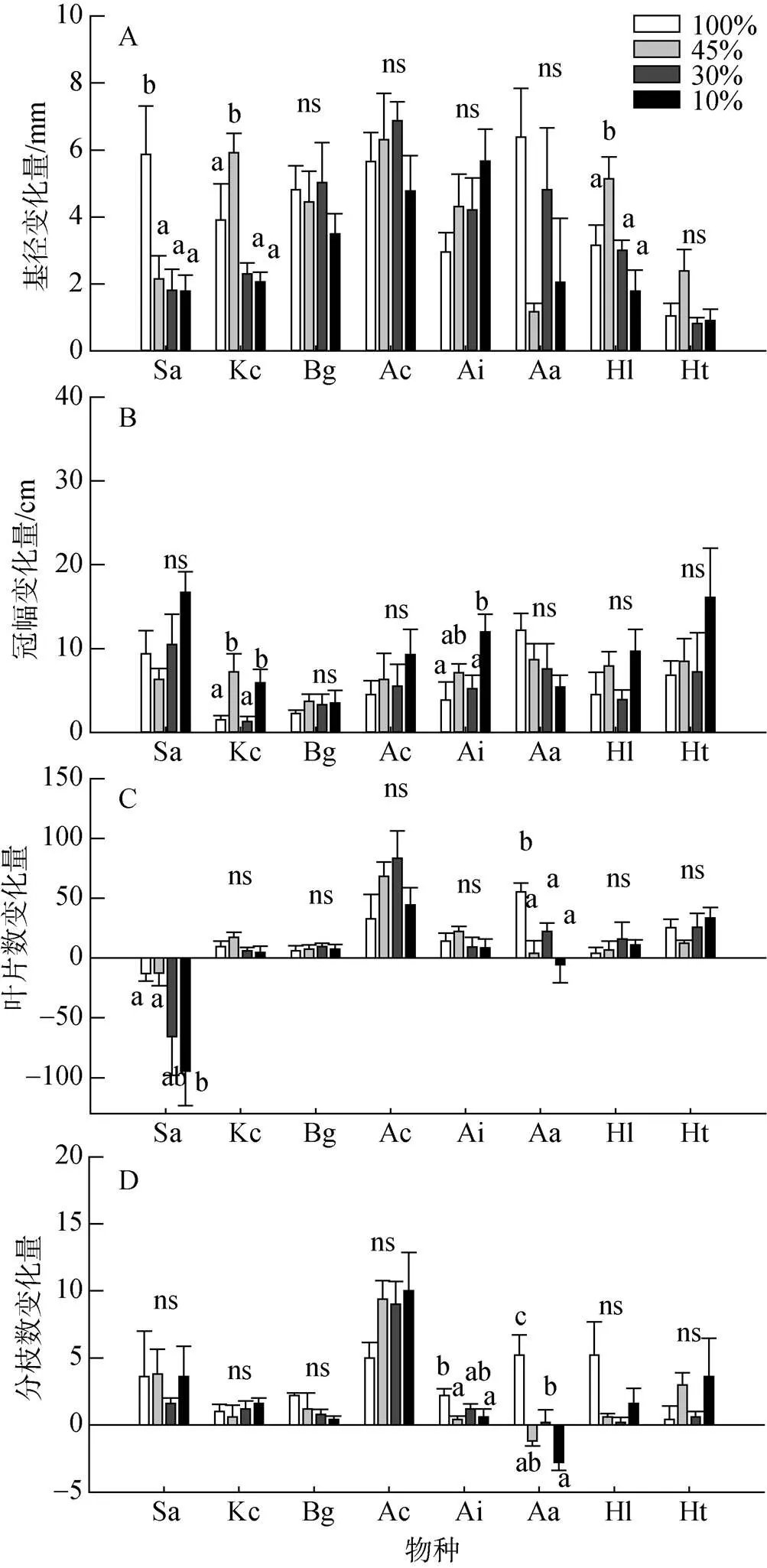
注: 物种缩写见表1 ; ns和相同字母表示在P=0.05水平差异不显著。
Figure 2 Changes in stem basal diameter, crown breadth, leaves number and branches number of seedlings of the eight mangrove species under four light treatments
3 讨论
有关人工遮荫或自然遮荫(不同光照强度)对植物生长发育影响的研究报道已有很多[22-25], 不同光照强度对一些植物的株高、生物量、光合特性等都有较显著的影响[26]。有研究表明, 生长在弱光下的植物, 相对生长率较低, 叶面积减少, 生物量降低[27]。一般情况下, 植物为了适应荫蔽的环境, 会增加植株高度向以获得更多的光能[28], 刁俊明和陈桂珠的研究表明在相同盐度处理下盆栽桐花树植株的茎高年增量随光照强度减弱而增大, 而基径年增量则随光照强度减弱而减小[17]。但在本实验中, 除老鼠簕外其他红树植物的植株高度并未显著增加,这可能是由于红树植物本身生长速度相对缓慢导致。本实验结果表明遮荫对无瓣海桑幼苗成活率、年茎高增长量、年基茎增长量、平均叶片数、年生物量积累和相对生长速率均有显著的抑制作用, 这与刁俊明等人的结果一致[29]。但在3个遮荫处理间, 无瓣海桑幼苗各生长指标不存在显著差异, 说明当光强下降到45%时, 无瓣海桑幼苗的生长即会受到抑制, 但这种抑制作用并不随光照强度的进一步下降而增强。同时, 光照条件的不同会导致植物生物量分配会增加, 而强光照下, 根的生物量分配会增加[30]。本研究表明: 当光照强度降低时, 无瓣海桑、秋茄、老鼠簕、桐花树的幼苗都对地上部分投入了更多的生物量, 其中秋茄、老鼠簕、桐花树通过提高叶生物量来获取更大的光能捕获能力, 弥补光能的减少。本研究在在以往针对单个红树树种(如无瓣海桑、桐花树)的遮荫试验研究基础上, 而本研究系统地针对常见的8种红树植物(包含外来种和本土种、真红树和半红树等各类红树植物)系统开展其幼苗对光照的生长和生理生态适应研究, 旨在综合评价这8种红树植物幼苗的耐阴性, 结果可直接应用于红树林造林实践。
注: 物种缩写见表1 ; ns和相同字母表示在P=0.05水平差异不显著。
Figure 3 Changes of biomass, relative growth rate(RGR) , shoot/root dry matter ratio(S/R) , lLeaf dry matter ratio (LMR), root dry matter ratio (RMR) and stem dry matter ratio (SMR) of seedlings of the eight mangrove species under four light treatments
仅从存活率的角度看, 7种本土红树植物都能够在林下种植。但是从生长适应的角度考虑, 桐花树和老鼠簕幼苗在遮荫处理下生长状况较好, 遮荫甚至对老鼠簕幼苗茎高、冠幅、生物量的增长以及相对生长速率均有显著的促进作用; 卤蕨幼苗在不同遮荫处理下都能够保持90%以上的存活率, 并且其生长状况没有受到显著影响。因此, 桐花、老鼠簕和卤蕨都比较适合在林下种植。这与刘滨尔研究调查发现淇澳岛无瓣海桑人工林内自然更新的主要植物是老鼠簕和卤蕨的现象符合[19]。秋茄幼苗在弱光条件下生长状况较好, 比较适合在郁闭度较低的林下种植。而半红树黄槿幼苗虽然在遮荫处理下存活率较高, 但其茎高生长量、基径变化量、分枝数变化量和生长速率在低光处理下都有所降低, 说明遮荫不利于黄槿的正常生长。在本研究中, 木榄幼苗耐阴性低于其他红树植物, 这与刘滨尔在淇澳岛进行无瓣海桑林分改造实践中得出的木榄与无瓣海桑混交林最成功的结果不一致[19], 原因可能是木榄的年龄级和本研究中的试验材料不同, 幼苗高度较高时利用光照的能力也较强。另外, 在造林实践中还需要考虑其他综合因素, 例如秋茄虽然耐阴性较强, 但是枝干脆弱容易受到上层林木压折, 因此林下存活率不高。而木榄茎干比较粗壮、抗压能力强且在45%和30%遮荫处理下仍有较高的存活率, 因此在郁闭度相对较低的无瓣海桑林下混交也有较好的效果, 而在裸滩的本土红树林群落营造则不存在这种情况。因此, 在红树林造林实践中应根据不同的造林目的选择适宜的造林树种, 而光照作为影响红树幼苗生长的关键因子, 其对红树幼苗生长和生物量分配的影响结果是合理混交造林、林分改造和优化红树群落配置的重要参考依据。今后的研究还需关注盐度、潮汐、光照以及土壤质地对红树幼苗的综合交互作用, 从而为造林实践提供更科学的指导。
红树林生态系统由于其自身环境条件限制, 其物种多样性远远低于陆地森林生态系统[31], 而我国在红树林人工恢复的过程中以种植纯林为主[32], 导致大部分红树林林分单一, 群落结构简单, 生态系统相对脆弱, 容易爆发大面积的病虫害, 降低红树林生态系统的功能和质量。因此, 如何合理地进行红树林混交造林是提高红树林造林存活率和造林质量至关重要的环节。本研究结果反映了不同红树植物耐阴能力的差异, 一方面有助于在无瓣海桑等速生林林分改造时更好地选择适宜的改造树种, 另一方面在本土红树群落营造时, 以此为依据对红树植物物种进行优化组合, 并在此基础上实现乔-灌-草合理配置模式。例如, 根据耐阴能力的差异采用秋茄、木榄作为上层乔木层, 桐花、老鼠簕作为中层灌木层以及卤蕨作为下层草本层。通过优化红树林的群落配置结构, 不仅可提高红树林的生物多样性, 而且能更好地恢复和提升红树林湿地生态系统结构和生态服务功能。同时, 有研究表明遮荫对入侵植物互花米草的生长具有显著的抑制作用, 并有学者提出通过无瓣海桑对互花米草进行生态控制[33-34], 本研究下一步将探讨无瓣海桑与耐阴性较强的桐花树和老鼠簕、卤蕨混合种植是否达到对互花米草更好的生态控制效果。结合竞争替代的生物手段实现互花米草的彻底清除, 实现真正意义上的“退草还林”, 从而更好地维持红树林生态系统的生态平衡和丰富生物多样性。
4 结论
本研究表明: 遮荫对无瓣海桑生长的抑制作用与其他红树植物相比更显著; 而桐花和老鼠簕在遮荫处理下生长状况较好, 光强的降低甚至对老鼠簕的茎高、冠幅、生物量的增长以及相对生长速率有显著的促进作用; 卤蕨在低光条件下存活率高且生长状况不受显著影响。因此, 从红树植物对光强的生长适应的角度来看, 桐花、老鼠簕和卤蕨都比较适宜种植在郁闭度较高的林下; 秋茄耐荫能力相对较弱, 适宜种植在郁闭度较低的林下或者作为中上层树种; 真红树无瓣海桑和木榄以及半红树黄槿和银叶树则不适宜直接在林下种植, 适合作为上层树种或者以纯林方式种植。根据不同红树植物耐阴性差异进行群落优化配置和林分改造将进一步丰富红树林生态系统的生物多样性, 更好地提升红树林生态服务功能。
[1] 林鹏. 中国红树林研究进展[J]. 厦门大学学报(自然科学版), 2001, 40(2): 592–603.
[2] 穆大刚. 五种红树植物光合产量模型及对环境因子的响应[D]. 青岛: 中国科学院海洋研究所, 2008.
[3] 廖宝文, 张乔民. 中国红树林的分布、面积和树种组成. 湿地科学, 2014, 12(4): 435–440.
[4] DUKE N, MEYNECKE JO, DITTMANN S, et al. A world without mangroves?[J]. Science, 2007, 317(5834): 41–42.
[5] GILMAN EL, ELLISON J, DUKE NC, et al. Threats to mangroves from climate change and adaptation options: a review[J]. Aquatic Botany, 2008, 89(2): 237–250.
[6] PARIDA AK, JHA B. Salt tolerance mechanisms in mangroves: a review[J]. Trees, 2010, 249(2): 199–217.
[7] TAKEMURA T, HANAGATA N, SUGIHARA K, et al. Physiological and biochemical responses to salt stress in the mangrove,[J]. Aquatic Botany, 2000, 68(1): 15–28.
[8] KRAUSS KW, TWILLEY RR, DOYLE TW, et al. Leaf gas exchange characteristics of three neotropical mangrove species in response to varying hydroperiod[J]. Tree physiology, 2006, 26(7): 959–968.
[9] 陈鹭真, 王文卿, 林鹏. 潮汐淹水时间对秋茄幼苗生长的影响[J]. 海洋学报, 2005, 27(2): 141–147.
[10] MEEKINS JF, MCCARTHY BC. Responses of the biennial forest herbto variation in population density, nutrient addition and light availability[J]. Journal of Ecology, 2000, 88(3): 447–463.
[11] CUSHMAN J, LORTIE CJ, CHRISTIAN CE. Native herbivores and plant facilitation mediate the performance and distribution of an invasive exotic grass[J]. Journal of Ecology, 2011, 99(2): 524–531.
[12] MONTGOMERY R, CHAZDON R. Light gradient partitioning by tropical tree seedlings in the absence of canopy gaps[J]. Oecologia, 2002, 131(2): 165–174.
[13] HOLSTE EK, KOBE RK, VRIESENDORP CF. Seedling growth responses to soil resources in the understory of a wet tropical forest[J]. Ecology, 2011, 92(9): 1828–1838.
[14] 刁俊明, 彭逸生, 郑明轩, 等. 红树植物对低光照水平的生长及生理生态响应研究进展[J]. 嘉应学院学报, 2009, 27(3): 69–74.
[15] 刁俊明, 陈桂珠. 盆栽桐花树对不同遮光度的生理生态响应[J]. 生态学杂志, 2011, 30(4): 656–663.
[16] SMITH SM, LEE DW. Effects of light quantity and quality on early seedling development in the red mangrove,L. [J]. Bulletin of Marine Science, 1999, 65(3): 795–806.
[17] 叶勇, 谭凤仪, 卢昌叉. 土壤结构与光照水平对秋茄某些生长和生理参数的影响[J]. 植物生态学报, 2001, 25(1): 42–49.
[18] 廖宝文, 郑松发, 陈玉军, 等. 外来红树植物无瓣海桑生物学特性与生态适应性分析[J]. 生态学杂志, 2004, 23(1): 10–15.
[19] 刘滨尔. 淇澳岛无瓣海桑人工林的自然更新特征及林分改造效果[D], 广州: 中国林业科学研究院热带林业研究所, 2013:2–3.
[20] 廖宝文, 田广红, 杨雄邦, 等. 珠海淇澳岛无瓣海桑种苗天然更新与扩散分析[J]. 生态科学, 2006, 25(6): 485– 488.
[21] 黄陵, 詹潮安. 粤东沿海引种无瓣海桑试验研究[J]. 林业实用技术, 2003, 5(2): 7–8.
[22] BALL M, CHOW W, ANDERSON J. Salinity-induced potassium deficiency causes loss of functional photosystem II in leaves of the grey mangrove,, through depletion of the atrazine-binding polypeptide[J]. Functional Plant Biology, 1987, 14(3): 351–361.
[23] BALL MC, CRITCHLEY C. Photosynthetic responses to irradiance by the grey mangrove,, grown under different light regimes[J]. Plant Physiology, 1982, 70(4): 1101–1106.
[24] BJÖRKMAN O, DEMMIG B, ANDREWS TJ. Mangrove photosynthesis: response to high-irradiance stress[J]. Functional Plant Biology, 1988, 15(1-2): 43–61.
[25] CHEESEMAN J, CLOUGH B, CARTER D, et al. The analysis of photosynthetic performance in leaves under field conditions: A case study using Bruguiera mangroves[J]. Photosynthesis Research, 1991, 29(1): 11–22.
[26] THOMPSON W, HUANG L, KRIEDEMANN P. Photosynthetic response to light and nutrients in sun-tolerant and shade-tolerant rainforest trees. Ⅱ. Leaf gas exchange and component processes of photosynthesis[J]. Functional Plant Biology, 1992, 19(1): 19–42.
[27] ELLISON AM, FARNSWORTH EJ. Seedling survivorship, growth, and response to disturbance in[J]. American Journal of Botany, 1993, 80(10): 1137–1145.
[28] 王俊峰, 冯玉龙. 光强对两种入侵植物生物量分配, 叶片形态和相对生长速率的影响[J]. 植物生态学报, 2004, 28(6): 781–786.
[29] 刁俊明, 曾宪录, 陈桂珠. 无瓣海桑幼苗对不同遮光度的生理生态响应[J]. 生态学杂志, 2010, 29(7): 1289– 1294.
[30] 刘滨尔, 廖宝文. 老鼠簕幼苗在潮汐环境下对不同光强的生理生态响应[J]. 林业科学研究, 2013, 26(2): 192– 199.
[31] 林益明, 林鹏. 中国红树林生态系统的植物种类、多样性、功能及其保护[J]. 海洋湖沼通报, 2001, 3(2): 8–16.
[32] 曾雯珺, 廖宝文, 陈先仁等. 无瓣海桑与三种乡土红树植物混交的生态效应[J]. 生态科学, 2008, 27(1): 31–37
[33] 李贺鹏, 张利权. 外来植物互花米草的物理控制实验研究[J]. 华东师范大学学报(自然科学版), 2007, 11(6): 44–55.
[34] 唐国玲, 沈禄恒, 翁伟花, 等. 无瓣海桑对互花米草的生态控制效果[J]. 华南农业大学学报, 2007, 1(28): 10–13.
Effects of light intensity on growth and biomass allocation of seedlings of the eight mangrove species
TAN Shujuan1,2, LI Ting1,2,5, YU Surui4, CAI Shuihua1,2, YE Wanhui1,3, SHEN Hao1,3,*
1. Key Laboratory of Vegetation Restoration and Management of Degraded Ecosystems/Guangdong Provincial Key Laboratory of Applied Botany, South China Botanical Garden, Chinese Academy of Sciences, Guangzhou 510650, China 2. University of Chinese Academy of Sciences, Beijing 100049, China 3. Southern Marine Science and Engineering Guangdong Laboratory (Guangzhou), Guangzhou 511458, China 4. College of Horticulture and Landscape Architecture, Zhongkai University of Agriculture and Engineering, Guangzhou 510225, China 5. South China Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Guangzhou 510300, China
The characteristics of growth and biomass allocation in seedlings of eight mangrove plant species(Linn.,Blanco.,Linn.,Lam.,Druce.,Ait.,L.) under different light intensities (100%, 45%, 30%, 10% of full sunlight) were studied using shading control experiment. The results showed that: (1) Shading significantly inhibited the relative growth rate, biomass accumulation, basal diameter growth and leaf number accumulation ofseedlings. (2) Seedlings ofandgrew well under shading treatments. Shading had no significant effect on the growth ofseedlings, but promoted the growth ofseedlings.(3) With the decrease of light intensities, seedlings of,,andallocated more biomass to their aboveground growth, and among them, seedlings of the latter three species increased biomass allocation to leaves to improve the ability of light capture. The results indicate that from the perspective of growth adaptation of light intensities,,andare suitable for planting under forests with high canopy density, andcan be planted under forests with low canopy density, while,,andare not suitable for planting under forests.
mangrove forest; light intensity; relative growth rate; biomass; biomass allocation
10.14108/j.cnki.1008-8873.2020.03.019
Q945.3; Q945.79
A
1008-8873(2020)03-139-08
2019-06-05;
2019-08-09
广东省科技计划项目(2017A020217001); 广东省林业科技创新项目(2017KJCX036, 2019KJCX015); 南方海洋科学与工程广东省实验室(广州)人才团队引进重大专项(GML2019ZD0408); 科技基础资源调查专项(2017FY100702); 国家科技支撑计划项目(2012BAC07B04, 2017YFC0505802); 国家重点研发计划项目(2017YFC0505802)
谭淑娟(1994—), 女, 湖北宜昌人, 硕士, 主要从事植物生理生态学研究, E-mail: tanshujuan@ scbg.ac.cn
沈浩, 博士, 副研究员, 主要从事保护生态学和植物生理生态学研究, E-mail: shenhao@ scbg.ac.cn
谭淑娟, 李婷, 余素睿, 等. 光照强度对8种红树植物幼苗生长和生物量分配的影响[J]. 生态科学, 2020, 39(3): 139–146.
TAN Shujuan, LI Ting, YU Surui, et al. Effects of light intensity on growth and biomass allocation of eight mangrove seedlings[J]. Ecological Science, 2020, 39(3): 139–146.
猜你喜欢
小猕猴学习画刊(2022年4期)2022-05-05
儿童故事画报·自然探秘(2022年3期)2022-04-27
老年博览·上半月(2021年10期)2021-11-02
幼儿园(2021年4期)2021-07-28
海洋通报(2020年5期)2021-01-14
中学生百科·小文艺(2020年6期)2020-01-13
儿童故事画报(2019年12期)2019-01-07
小天使·二年级语数英综合(2018年3期)2018-06-29
歌海(2018年4期)2018-05-14
天然产物研究与开发(2016年6期)2016-06-05
