
荒漠森林开垦成农田前后土壤呼吸速率的变化及其影响因素
2020-06-15 09:33杨晓东吕光辉
水土保持通报 2020年2期
杨 军, 杨晓东, 吕光辉, 崔 海
(1.银川能源学院, 宁夏 银川 750199; 2.宁波大学 地理与空间信息技术系,浙江 宁波 315211; 3.新疆大学 资源与环境科学学院, 新疆 乌鲁木齐 830046)
干旱荒漠面积约占全球陆地面积的1/4,其土壤碳库在全球碳循环中扮演着十分重要作用[1]。在严酷环境的限制下,干旱荒漠的土壤呼吸对全球变化最为敏感[2-3]。新疆维吾尔自治区是中国荒漠分布面积最多的省,荒漠生态系统在保障当地社会发展和维护生态安全等方面,都具有十分重要的意义。但近几十年来,荒漠森林被大规模开垦并转变为绿洲[4]。这一过程在改变地表植被的同时,也改变了土壤碳储量和土壤呼吸,进而对当地土壤碳收支和全球范围的碳循环产生影响[5-6]。但在目前,对荒漠森林转变为绿洲过程中土壤碳排放的变化并不清晰。
荒漠森林转变为绿洲后,大多数被用来从事农业生产。众所周知,水分和“随水而来随水而去”的土壤盐分是干旱区荒漠及荒漠基底下农田系统的主要生态限制因子[7-8]。相对荒漠森林,耕作活动能使农田的土壤水分和盐分含量发生变化,进而对农作物根系生物量、微生物活性和有机质分解速率产生影响[5,9]。由于农田系统中土壤呼吸主要来源于农作物的根际呼吸、微生物呼吸和土壤中有机质的分解[10]。因而,农田耕作活动可能会改变土壤呼吸的数值[11-12]。此外,耕作活动对土壤呼吸的影响大小可能与耕作年限有关[13]。这是因为干旱荒漠森林土壤十分贫瘠,开垦为农田后,施肥、灌溉、轮作等管理方式都能减小土壤盐分、增加水分和有机质含量[14-16]。因而,在荒漠森林开垦为农田的种植年限上,土壤呼吸可能存在差别。
本文在新疆维吾尔自治区艾比湖荒漠地区的托托乡内,选择不同连续耕作年限的农田、以及未开垦的荒漠森林为对象,通过观测不同样地的土壤呼吸速率以及影响土壤呼吸速率的水盐、地下细根生物量和微生物菌落数量等因子,旨在回答以下2个科学问题:①将荒漠开垦后为农田后,土壤呼吸速率是否随农田连作年限发生变化。②水盐作为干旱区荒漠的限制因子,它们在连作年限棉田间的差别是否引起了土壤呼吸速率的变化。
1 研究区概况
新疆维吾尔自治区的艾比湖地区处于亚欧大陆腹地,在远离海洋和三面高山围绕的综合影响下,属典型温带大陆性气候。气候极端干燥,降水稀少,冬、夏季漫长,春、秋季短暂。全年日照时数约2 800 h,年降水不超过100 mm,蒸发量1 600 mm以上,年均无霜期为160 d,年平均气温6~8 ℃,极端最高气温44 ℃,极端最低气温-33 ℃[17]。西北部阿拉山口是全国著名的风口,盛行西北风,全年8级以上大风达165 d,多集中在4—6月[17]。
本文所选择的样地均在艾比湖地区的托托乡内(44°33′—44°37′N,83°33′—83°32′E),其位于精河县东部婆罗科努尔北麓的山前冲积扇绿洲上,位置在艾比湖的西南部。土壤为荒漠土,盐渍化明显,土壤0—20 cm表层含盐量高达4%~8%。植被区域属于新疆荒漠植被区中北疆荒漠植被亚区,旱生荒漠植物是当地主要植被类型[12,17]。从21世纪80年代开始,托托乡周边大量以梭梭(Haloxylonammodendron)和胡杨(Populuseuphratica)为建群种的荒漠森林开垦为农田,其上种植棉花、玉米、油葵等,其中棉花是主营农作物。
2 试验设计和研究方法
2.1 试验设计
基于1984—2014年绿洲边缘扩展位置和询问当地住户结果,沿托托乡至艾比湖的方向上,我们选择开垦后连续种植30 a,16 a和3 a的农田,以及未开垦的荒漠林地(胡杨+梭梭林)各3块。胡杨+梭梭林和30 a,16 a和3 a农田的距离大致分别为7,5和3.5 km。胡杨+梭梭林和30 a,16 a和3 a的海拔高度分别为298,377,333和397 m。在每块上设置了10 m×10 m样地各1个,共设置样地12个。托托乡是一个以农业为主的乡,属于兵团统一企业式管理,耕作模式、种植密度、施肥水平、灌溉方式等田间管理方式一致。所选择试验农田开垦之前均为自然发育的原始荒漠,土壤为沙质土壤,土壤容重在0.98~1.25 g/cm3之间,土壤总孔隙度在40%~43%之间。开垦后,土壤容重降低,大致在0.78~0.93 g/cm之间,相反土壤总孔隙度增加,数值在45%~49%之间。农田种植作物均为棉花,一年种植1季,前茬也是棉花(当地多数农田连作种植棉花,甚至有连作棉花几十年以上的农田)。棉花在4月中下旬播种,在9月中旬收获。收获后地上茎叶被收割机直接从田中带走,不实施秸杆还田。棉田耕作栽培方式为拌种、深耕(20 cm)地膜。基肥品种为美国二胺和过磷酸钙,年均用量约每亩20 kg,不施有机肥,在播种时将基肥的80%施到田中,后分别在7和8月下旬追肥二次,追肥量约为总量20%。播种前期10 d左右进行春灌,在7月中旬、8月上旬、8月下旬的棉花生长期间进行人工灌溉,其他时间不灌溉。
2.2 土壤呼吸速率和环境因子测定
艾比湖地区棉花的生长季为4—10月,其中6和9月分别是棉花的花期和吐絮期,是棉花生理最为旺盛的两个时期。本文为分析土壤呼吸速率在不同耕作年限上的差别,在6和9月,分别测定了棉田的土壤呼吸速率和环境因子。6和9月是时间梯度上对试验的重复。农田系统土壤呼吸的日变化及其影响因素,当前在艾比湖地区的研究较多,本文不做探讨。
本文利用开路式土壤碳通量测量系统Li-8100(Li-CorInc,Lincoln, NE, USA)测定土壤呼吸速率。具体观测时,我们在连续种植30 a,16 a和3 a的农田,以及未开垦的荒漠胡杨+梭梭林的各样地中各随机设定3监测点,在每观测点上,在土壤中插入内径和高度分别为20和10 cm的PVC环,PVC环插入土壤深度约3 cm。2 d后在当地时间7:00至翌日5:00间测定土壤呼吸速率。日观测频度为每间隔2 h测定1次,具体时点为7:00,9:00,11:00,13:00,15:00,17:00,19:00,21:00,23:00,1:00,3:00,5:00时。共连续观测两天。在每个时间点测定土壤呼吸速率的同时,用Li-8100自带的温度探头和水分探头(Echo EC-5, Decagon Devices Inc, Pullman, WA, USA)分别测定土壤10 cm处土壤温度和土壤表层(0—10 cm)体积含水量。另外,用手持式气象仪(Kestrel 4000, Nielsen-Kellerman, Boothwyn, PA, USA)测定距离地表150 cm处的气温和大气相对湿度。
每样地土壤呼吸速率测定结束后,在每个观测点附近,采用内径2.5 cm的标准土钻采集0—20 cm深的土壤样品。土壤样品采集时首先除去土层上面的枯枝落叶层,其次为避免其它微生物混入土壤,土壤样品采集前用土钻预采集土壤3次。每观测点土钻采样2次。待所有观测点土壤样品采集完成后,将同一样地中土钻第1次采集土壤样品混合,装入自封袋中用于测定细根生物量。同时,将土钻第2次采集属于同一样地的土壤样品混合,并将其分为2份,1份装入自封袋中用于土壤理化性质的测定,另1份放入无菌纸袋进一步保存在4 ℃冰箱用于测定土壤微生物的菌落数量。
本文所选择的土壤理化性质有pH值、土壤总盐和土壤有机质。它们测定分别采用玻璃电极法、重量法和重铬酸钾法[18]。分别用牛肉膏蛋白胨、高氏和马丁氏培养基分别在37 ℃和28 ℃培养箱中培养细菌、放线菌和真菌,用菌落形成单位(colony-forming units, CFU)对微生物菌落计数的方法测定了微生物的菌落总数[19,20]。此外,我们将测定细根生物量的土壤样品取出,使用2 mm筛子将直径≤2 mm的细根从其内分离出来,冲洗后在80 ℃下烘干48 h至恒重,称重后重量(g)和取样面积(3×2.52×π/4=14.72 cm2)综合计算结果为细根生物量(g/m2)[21]。
2.3 数据处理
采用单因素方差分析比较了种植30 a,16 a和3 a的农田、以及未开垦的荒漠胡杨+梭梭林间土壤呼吸速率、细根生物量、土壤理化性质和微生物数量的差别。方差分析中,不同处理间(胡杨+梭梭林,30 a农田,16 a农田和3 a农田)方差齐性采用Tukey HSD进行检验;相反,方差不齐性采用Tamhane’sT3检验。采用多元线性逐步回归分析了土壤呼吸速率与细根生物量、土壤理化性质和微生物数量间的关系。多元线性回归时,土壤呼吸、土壤水分、土壤温度、气温和大气相对湿度在2 d共测定了24次,每个监测点共有24个数据点。相反,每个样地的土壤理化因子和微生物菌落总数在土壤呼吸测定结束后采样,每个监测点仅有1个数据点。两者数据不对应。考虑到土壤理化因子(pH值、土壤总盐和土壤有机质)和微生物菌落总数在24 h内变化不大,在回归时,将24个数据点的理化因子和微生物总数用1个数据点的数据进行补充,即:24个点具有相同的土壤理化因子和微生物菌落总数。数据处理均在SPSS 19.0中完成,图在Oringin 8.0中制作。
3 结果分析
3.1 荒漠森林开垦前后土壤呼吸速率的变化
在棉花的花期(6月)和吐絮期(9月),30 a的土壤呼吸速率>3 a=16 a>胡杨+梭梭林(林地),这说明土壤呼吸速率随农田耕作年限增长显著增加(p<0.001)(图1)。另外,开垦农田各时间段上的土壤呼吸速率均高于胡杨+梭梭林(p<0.001)。将6月和9月数据合并后计算平均值发现,30 a,16 a和3 a的土壤呼吸分别是胡杨+梭梭林的4.44,2.78,3.69倍。
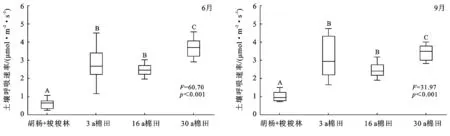
注:F和P为单因素方差分析的检验参数。每一个子图中,相同大写字母表示不同处理间没有显著性差异,不同大写字母表示有显著性差异。
图1 荒漠森林、不同耕作年限农田之间土壤呼吸速率的差异
3.2 荒漠森林开垦前后土壤理化性质的变化
在棉花的花期(6月),随连作农田耕作年限增长,土壤有机质、含水量、微生物菌落总数和细根生物量都显著增长(p<0.05)(表1);相反,土壤总盐和pH值显著减小(p<0.05),空气相对湿度未有变化(p>0.05),连作3 a和16 a的气温和10 cm深地表温度均高于连作30 a农田和胡杨+梭梭林(林地)(p<0.05)(表1)。同样,在棉花的吐絮期(9月),土壤有机质、含水量、微生物菌落总数、细根生物量、总盐和pH值在农田耕作年限上的变化和6月一致(表1)。相反,空气湿度、气温和10 cm深地表温度与6月表现明显不同。具体地:随农田耕作年限增长,空气湿度显著增加(p<0.05),10 cm深地表温度显著减小(p<0.05),气温未有显著变化(p>0.05)(表1)。

表1 土壤呼吸的影响因子在荒漠森林、不同农田连作年限间的差别
注:F和p为单因素方差分析的检验参数。表格同一行内,数字后面相同大写字母表示不同处理间没有显著性差异,不同大写字母表示有显著性差异。
3.3 荒漠森林和农田的土壤呼吸与理化性质之间的关系
多元线性逐步回归结果中,最终方程显示自变量是引起因变量变化的主要因素。根据模型回归标准系数的大小可确定自变量对因变量变化的影响程度。标准系数的绝对值越大,表明自变量对因变量变化的影响越大。将能引起土壤呼吸变化的因子对其多元逐步回归后发现(表2): ①在棉花的花期(6月),土壤呼吸速率的变化主要由土壤含水量和总盐决定,其各自对土壤呼吸速率变化的模型回归标准系数分别为0.60和-0.52; ②在棉花的吐絮期(9月),土壤呼吸速率的变化主要由土壤含水量和地下细根生物量决定,其各自对土壤呼吸速率变化的模型回归标准系数分别为0.49,0.48; ③将棉花的花期和吐絮期数据合并整体回归后发现,土壤呼吸速率的变化主要由土壤含水量和土壤总盐决定,其各自对土壤呼吸速率变化的模型回归标准系数分别为0.67和-0.42。这3个结果中,土壤总盐和含水量是最主要决定土壤呼吸速率变化的因素。

表2 土壤呼吸速率与土壤理化因子、微生物菌落总数、地下细根生物量之间的多元线性逐步回归关系
注:Y为土壤呼吸速率。
4 讨 论
以往研究[22-23]表明,森林和草原等植被开垦成农田后,由于植被类型的改变,土壤呼吸速率减小。与之相反,本文得出,荒漠森林开垦为农田后,土壤呼吸显著增加(p<0.001)(图1)。这可能是因为在土壤十分贫瘠、本身所含有机质和无机碳等十分稀少的干旱荒漠中,原生森林开垦为农田后,耕作、施肥和灌溉等人为措施会改变土壤的水分、有机质和矿质离子等的含量,改善了土壤环境,进而使土壤中细根生物量和微生物数量增加,最终导致土壤呼吸速率增加[9,24]。但在当前研究中,另有观点认为,在不同种植年限的农田之间,农作物类型、灌溉方式、施肥品种、耕作模式(轮作与连作)和田间管理方式等也可以引起土壤呼吸的改变,使得它们有可能抵消土壤中的细根生物量、微生物多样性和数量和有机物底质对土壤呼吸速率的影响[25-27]。因而,在种植年限梯度上,可能出现土壤呼吸速率随种植年限增长降低或在年际间波动不大的结果[27-28]。但本研究结果与这些研究完全相反。这可能是因为,20世纪80年代后,伴随西北经济快速发展,新疆绿洲周边的荒漠森林逐渐被开垦为农田,用以种植棉花、小麦、油葵和玉米等作物。众多农业作物中,棉花作为新疆绿洲区的主要经济作物,在耕地有限条件下,经济价值和棉区规划使得轮作制度在新疆棉花种植地区难以实施,连作已成为该区域作物的主要种植模式[16,24]。但另一方面,连作过程中农作物种植类型、灌溉方式、耕作方式、施肥品种、田间管理方式等都大体一致[12]。因而,连作棉田中,农作物类型、灌溉方式、施肥品种、耕作模式和田间管理方式不是引起不同耕作时间农田中土壤呼吸变化的主要原因。这一点也可以通过土壤理化因子在不同耕作时间农田中的差异进行佐证。本研究发现,土壤中的细根生物量、微生物菌落总数和有机物含量也随棉花连作年限显著增加(p<0.05)(表1)。说明在新疆地区虽然用荒漠森林开垦的农田存有土壤盐渍化的天然缺陷,但如果在农业生产中,合理利用排盐渠等措施,并持续向土壤中施肥,会使得荒漠森林在开垦后的农田,逐渐出现土壤熟化过程,即:人为农业活动能够逐渐改善土壤质地,减小土壤容重,提高土壤中有机质含量和营养物质,增加土壤空隙和保水能力,并同时减少盐分含量等,使得土壤从不适合农田耕作逐渐转化为理化性质稳定、适合耕种并能够保证作物高产的土壤[9,27]。我们的结论还可以通过桂东伟和张晓东等人的研究进行核实。他们发现在干旱区绿洲,随耕作年限增加,土壤容重减小,土壤质地改变(黏粒比例增加,沙粒比例减小),土壤逐渐由不适合耕作向适耕土壤转变[15,29]。
本研究在艾比湖托托乡跨度30 a的农田上观测时,6月和9月土壤温度和气温在不同耕作年限农田间未有显著差别(p>0.05)(表1)。相反,水分和盐分含量存在显著差别(p<0.05)(表1)。此外,多元逐步回归结果显示,水盐是土壤呼吸速率模型中回归标准系数最高的两个因子。说明在干旱荒漠区的农田系统中,水分和“随水而来随水而去”的盐分可能是引起土壤呼吸变化的主要原因[11,33]。这一结果与前人研究土壤呼吸影响因素的成果不同[30-32]。以往的研究发现,由于温度能促进有机物分解酶的活性,温度被认为是引起土壤呼吸在不同土地利用类型之间变化的主要原因[30-31]。
本研究与这些研究呈现不同的结果可能是因为: ①土壤含水量能对土壤溶解性有机质、植物和微生物能量的分配、土壤通透性、植物根系与微生物活动等产生影响。较低含水量会造成微生物活性和细根生物量下降,对土壤呼吸产生抑制作用[34,35]。尤其对土壤微生物,极低水分使得土壤微生物很难繁殖生长[11]。本文中,土壤含水量在荒漠森林最低,可能造成土壤微生物活性、细根生物量均数值较小,由此导致荒漠森林拥有较低的土壤呼吸速率。将荒漠森林开垦成农田后,耕作活动显著改善了土壤水分含量,进而改善了微生物的生存环境。它们开始大量繁殖生长,代谢消耗有机质,最终导致开垦后农田的土壤呼吸显著提高。②盐渍土中,盐分能限制土壤结构更新,降低团聚体的稳定性和有机质的含量,因而对土壤呼吸产生负作用[36-37]。在干旱荒漠的农田中,排盐渠、漫灌压盐、施肥等人为耕作活动能够降低土壤含盐量,进而引起土壤中的细根生物量、微生物数量和有机物含量随耕作年限不断增加,最终造成了土壤碳释放量在耕作年限间的差别。③本研究区域范围较小,从荒漠森林样地到30 a耕作农田的距离在7 km左右,加上当地属于山前冲积平原,海拔差异较小[11,17]。由此使得土壤温度和气温在荒漠森林、3种不同耕作时间农田之间未有显著性差别。
5 结 论
在梭梭+胡杨为建群种的荒漠森林被开垦为农田后,随棉田耕作年限增长,土壤呼吸速率显著增加。相对荒漠森林,在6月和9月,耕作30 a,16 a和3 a农田的土壤呼吸速率分别增加了4.44,2.78,3.69倍。说明在干旱区荒漠,荒漠森林开垦以及其后的耕作活动增加了土壤碳释放量。另外,在土壤呼吸作为因变量的多元逐步回归方程中,水盐的标准系数分别为0.67和-0.42,是回归方程中仅剩的因子。这说明土壤水盐是引起土壤呼吸在不同耕作年限的土壤之间变化的主要原因。综合可知,将以梭梭+胡杨为建群种的原生荒漠森林开垦为农田后,人为耕作活动改善了土壤的水盐情况,使土壤含水量增加而含盐量不断减小,由此引起了碳释放量在不同耕作时间农田间的差别。
猜你喜欢
小哥白尼(趣味科学)(2022年5期)2022-08-15
东北林业大学学报(2022年6期)2022-07-25
绿色科技(2021年5期)2021-04-09
绿色中国(2019年14期)2019-11-26
四川农业大学学报(2018年3期)2018-09-04
儿童故事画报·智力大王(2016年7期)2017-02-08
中国工程咨询(2017年7期)2017-01-31
文史春秋(2016年3期)2016-12-01
中国农业信息(2016年10期)2016-02-06
土壤与作物(2015年3期)2015-12-08
