
鸭疫里默杆菌表型相关基因在重庆、四川分离株中的分布及遗传多样性分析
2020-07-22 10:14高继业黎容红李德龙陈思怀陈学情
中国兽医学报 2020年7期
高继业,黎容红,黄 伟,*,李德龙,陈思怀,陈学情
(1.西南大学 动物科学学院,重庆 402460;2.重庆更尚科技有限公司,重庆 402460)
鸭疫里默杆菌感染(Riemerellaanatipestiferinfection)又称鸭传染性浆膜炎(infectious serositis),是当前危害我国养鸭业最为严重的疫病之一,发生率和致死率最高可达75%以上,其病原为鸭疫里默杆菌(Riemerellaanatipestifer,R.anatipestifer)[1-11]。R.anatipestifer在我国至少有25个血清型,不同地区或同一地区的不同时间流行的血清型存在较大差异,且各血清型间缺乏有效的交叉免疫保护抗原物质[6-11]。R.anatipestifer分布的广泛性、血清型的多样性、菌株间致病力的巨大差异性和不断增强的耐药性,给感染的防治工作带来严峻挑战,也对我国食品安全构成了巨大威胁[8-11]。虽然R.anatipestifer全基因组序列的测序工作已经完成,在GenBank登录序列多达33个,但很多基因的功能还没有被明确的注释。尽管关于R.anatipestifer毒力因子的研究不断增多,其中包括协同溶血素(CAMP)、血凝素(Hemaglutin,Hema)、外膜蛋白A(OmpA)、三磷酸甘油醛脱氢酶(GAPDH)等,但其致病力发生差异的分子基础和可能机制一直没有得到明确的解释[12-20]。
细菌的遗传学因素在其致病力方面发挥着巨大的作用,主要包括:细菌对外环境信号的感受和应答,基因转录的调控,毒力相关操纵子和调节子的组成,以及质粒、毒力岛等移动的遗传学因子[21-22]。因此,某些特定基因的存在或表达必将决定细菌的某些生物表型,如毒力、耐药性等,而从这些特定的基因或相关区域中找出特异的DNA片段具有重要价值,如幽门螺旋杆菌新的限制-修饰系统,沙门菌血清型特异性基因,奈瑟球菌属病原特异性基因等,即使找出的这些基因片段的功能不清楚,但也可作为诊断的分子标记[23-25]。为此,本研究以重庆、四川地区的R.anatipestifer流行菌株为研究对象,检测部分生物表型相关基因在其基因组中的分布,分析其出现的频率与保守性,将有助于探讨R.anatipestifer感染致病的分子基础,对阐明其致病力发生差异的分子基础及可能机制具有重要意义。
1 材料与方法
1.1 菌株及质粒22株R.anatipestifer均分离自重庆、四川地区的(隐性)感染鸭或鹅,纯化后经生化试验、16S rRNA编码基因序列测定和比对分析鉴定,由本研究室冻干保存(表1)。

表1 基因PCR检测菌株
1.2 主要试剂胰蛋白胨大豆琼脂(Trypticase Soy Agar,TSA),OXOID公司产品;Goldview核酸染料、X-gal、IPTG、氨苄青霉素,重庆鼎国生物试剂有限公司产品;MiniBEST Bacterial Genomic DNA Extraction Kit Ver.2.0、MiniBEST Plasmid Purification Kit Ver.4.0、MiniBEST Agarose Gel DNA Extraction Kit Ver.4.0、TaKaRaTaqTM、DNA molecular weight marker,宝生物工程(大连)有限公司产品。
1.3 基因的选择及引物设计参照文献[12,15,26-28],并根据GenBank登录的R.anatipestifer全基因组序列(表2)进行分析,选取18个生物表型相关基因,采用引物设计软件Primer 5.0设计引物,对目标基因进行PCR扩增和克隆测序分析,扩增的基因及引物见表3。

表3 PCR检测基因及引物序列和定位
1.4 PCR扩增取目标菌株的TSA新鲜培养物,参照试剂盒说明书提取细菌基因组DNA。以提取的基因组DNA为模板,进行PCR扩增,扩增基因及引物序列见表2,。采用25 μL反应体系:10×PCR Buffer 2.5 μL、MgCl2(5 mmol/L)2.5 μL、dNTP(2.5 mmol/L)2 μL、DNA模板2 μL、上下游引物(10 μmol)各1 μL、rTaq酶(5 U/μL)0.25 μL,无菌ddH2O补至终体积25 μL。反应参数:94℃预变性5 min,按94℃ 1 min、52℃ 1 min、72℃ 2 min循环30次,最后72℃延伸5 min,4℃保持5 min。同时设不加模板的阴性对照。

表2 参考基因组信息
1.5 PCR产物纯化及测序分析PCR产物经10 g/L 琼脂糖凝胶电泳分析、切胶回收、纯化后送北京华大基因公司测序,采用DNAStar软件对获得的DNA序列进行拼接和同源性的比较分析,并建立系统发育树。
1.6 RA、RS、RC菌株的全基因组序列测序分析取R.anatipestifer菌株RA、RS、RC对数生长期培养物,0.1%甲醛灭活后送杭州谷禾信息技术有限公司进行全基因组测序,使用CLC Genomics Workbench 6.5进行二代数据的基因组拼接。将获得的全基因组序列与GenBank登录序列进行遗传多样性比对分析(表2)。
2 结果
2.1R.anatipestifer生物表型相关基因的PCR扩增PCR扩增结果显示,在18个相关表型基因中,外膜蛋白A(OmpA)、类脂A生物合成酰基转移酶(LipidA)、血凝素(Hema)、协同溶血素(CAMP)的编码基因均能在22株R.anatipestifer中检测到,表明这些基因在不同血清型菌株或不同地区的分离株中具有良好的保守性。另外,铁离子转运及利用相关蛋白铁依赖受体1(TbdR1)、铁依赖受体2(TbdR2)、铁依赖抑制子(DtxR)的编码基因均能在22株R.anatipestifer中检测到(表4)。从检测结果中还发现,除细胞壁相关水解酶(CWAH)、嗜好模块毒素(AMT)的编码基因外,其他16个基因均能在鹅源分离株基因组中检测到,而不具有致病性的菌株DX1、YC1、YC2均未能扩增出转录调控因子1(LuxR1)、转录调控因子2(LuxR2)、热休克蛋白(GroEL)、三磷酸甘油醛脱氢酶(GAPDH)、转录调节子(TR)、AMT、的编码基因。本研究还对18个生物表型基因在RA、RS、RC菌株全基因组序列中的分布进行了检索分析,结果显示:除AMT的编码基因未能在RS菌株中检索到外,其他基因均能在3株菌中检索到。
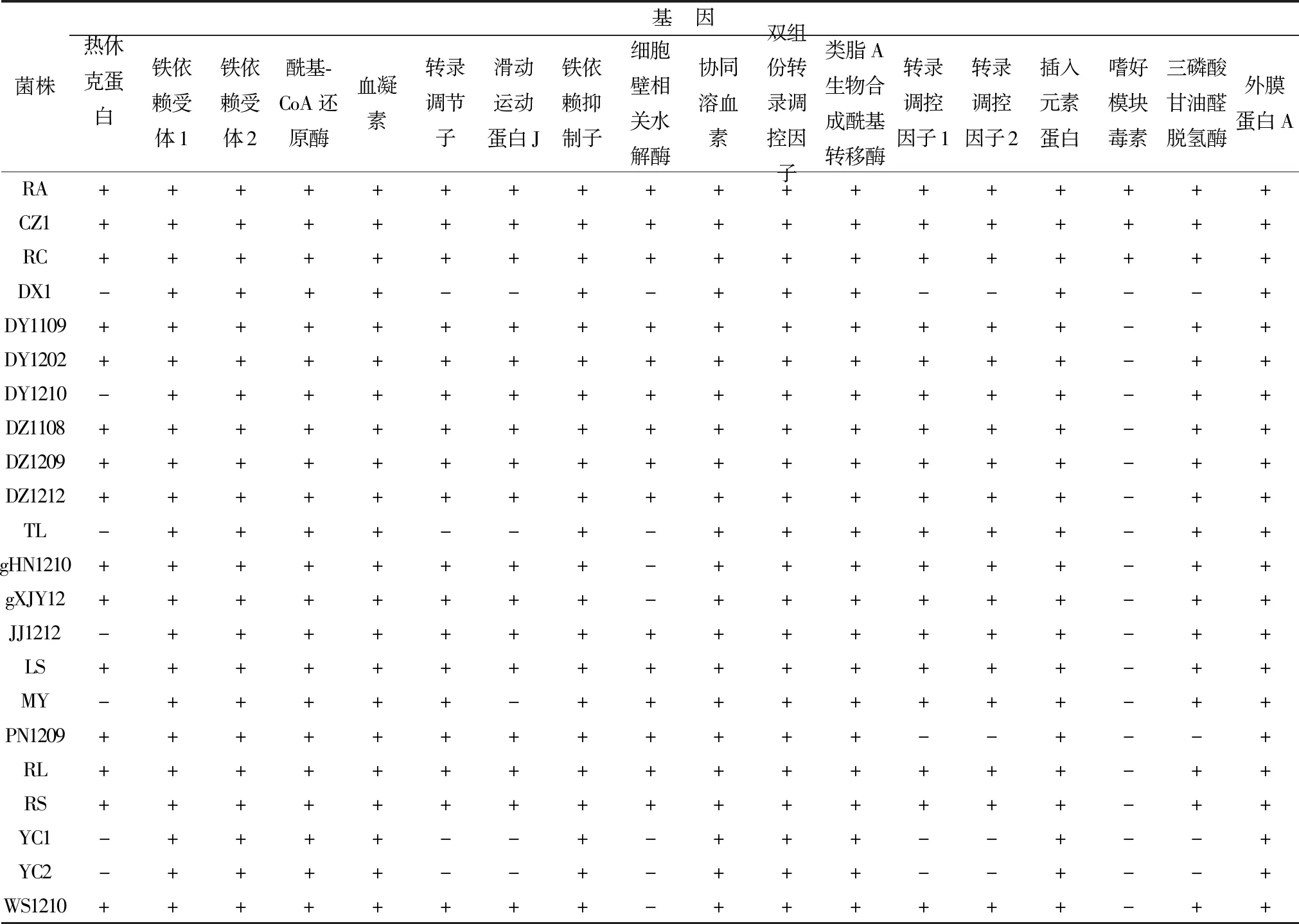
表4 18个表型基因在分离菌株中的PCR检测结果
2.2 阳性扩增基因的测序分析各基因的阳性PCR产物经纯化后送北京华大基因工程公司和南京金斯瑞生物工程公司进行测序。获得的序列采用MegAlign程序(DNAStar 5.01)进行遗传多样性比对分析(By Clustal W Method)。比对分析结果显示,阳性扩增基因序列与R.anatipestifer标准菌株ATCC1848相对应的序列的同源性均在93%以上,其中双组分转录调控因子(TCTR)、GroEL、LuxR1、插入素蛋白(IS1)、DtxR、TR、TbdR1、TbdR2的基因在阳性菌株中的同源性均在96%以上,而CAMP、Hema的编码基因在阳性菌株中存在较强的遗传多样性(图1);对3株非致病菌株DX1、YC1、YC2的阳性扩增基因进行序列比对分析发现,OmpA、LipidA、Hema、TbdR1、TbdR2、DtxR的同源性均在95%以上。
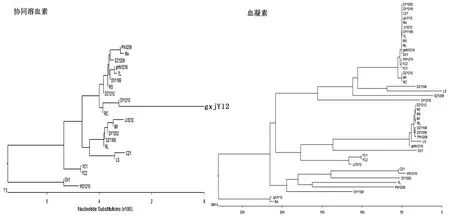
图1 协同溶血素和血凝素编码基因序列的遗传多样性分析
2.3 RA、RS、RC菌株的全基因组序列分析本研究还对RA、RS、RC 3株菌的全基因组进行了测序分析。分析结果显示,RA、RC、RS的基因组大小分别为2 129 862,2 172 619,2 067 830 bp,G+C含量分别为35.03%,35.09%,34.96%,蛋白编码基因数量分别为2 022,2 057,1 923,其中含多个抗生素耐受基因和毒力基因(表5)。获得序列与GenBank登录序列的进行遗传学多样性分析结果显示,血清Ⅰ型菌株RC与CH3、RA-CH-1处于同一分支,血清Ⅱ型菌株RA、RS与RA-GD、RA-SG、RA-YM属于同一分支,虽与ATCC11845菌株处于同一遗传节点,但遗传关系的密切程度存在差异。另外,RA、RS与同为血清Ⅱ型的菌株RA-CH-2处于不同的遗传节点上,其确切关系尚不清楚。

表5 RA、RS、RC菌株的全基因组特征
3 讨论
3.1R.anatipestifer基因组存在遗传多样性R.anatipestifer早在1932年就被分离获得,并于1997年被归于黄杆菌科第5核糖体RNA总科[1-3]。尽管GenBank上登录的R.anatipestifer全基因组序列已多达12个,但许多基因的功能都还没有明确的注释,其血清型、致病力等生物表型的多样性一直没有得到很好的解释。本研究将国内流行范围最广的血清Ⅰ、Ⅱ型菌株RA、RC、RS全基因组序列与GenBank登录序列进行了遗传多样性分析,分析结果表明:血清Ⅰ型菌株与血清Ⅱ型菌株分属于不同的分支,且血清Ⅰ型菌株间遗传关系非常接近,而血清Ⅱ型菌株间遗传关系的密切程度存在较大差异(图2)。结合生物表型相关基因的PCR检测结果猜测,这可能与菌株间存在不同程度的基因缺失有关,且血清Ⅱ型菌株发生缺失的频率显著高于血清Ⅰ型菌株。另外还发现,除RA-CH-2外的所有血清Ⅱ型菌株均与ATCC11845处于同一遗传节点,据此推测ATCC11845菌株可能属于血清Ⅱ型。另外,R.anatipestifer基因组出现这种遗传多样性和遗传学上相距甚远的可能原因有两个:一是通过疾病过程本身的突变和选择作用;二是优势血清型表型基因在R.anatipestifer群体中发生了水平传递或缺失。但无论是哪种情形,均可能存在一种与疾病相关的选择作用,这种选择作用将有利于增加那些遗传上相距很远但能表达优势血清抗原的菌株的出现频率与地理分布。

图2 鸭疫里默杆菌全基因组的遗传学多样性分析
3.2R.anatipestifer弱/无毒株的侵袭力相关基因存在缺失病原菌突破感染宿主机体的防御机能,在宿主体内的定居、繁殖及扩散和蔓延的能力是构成其侵袭力的主要因素,也是其毒力或致病性的重要组成部分。本研究对涉及R.anatipestifer在感染宿主体内定居、繁殖及扩散、蔓延的能力的相关蛋白的编码基因进行了PCR检测和测序分析,主要包括DtxR、TbdR1、TbdR2、CAMP、Hema、GldJ、GroEL。其中DtxR、CAMP、Hema与R.anatipestifer在感染宿主体内的铁/血红素代谢有关,GldJ被认为与细菌的运动密切相关,而作为真核生物热休克蛋白类似物的GroEL是一种高度保守的细胞内抗原,被认为可以保护细胞内病原体免受宿主吞噬细胞的恶劣环境的侵害,也被报道为多种感染中的有效免疫原[29-30]。分析结果显示,DtxR、TbdR1、TbdR2、GroEL的编码基因在阳性菌株中高度保守,同源性高达96%以上,而CAMP、Hema在阳性菌株中同源性存在较大差异,并形成3个相对独立的分支;另外,GroEL基因进在弱毒株MY、TL和无毒株DX1、YC1、YC2中呈阴性,GldJ基因在弱毒株MY、TL和无毒株DX1中呈阴性。据此推测,在感染宿主体内迁移和抗吞噬相关基因的缺失可能是R.anatipestifer无毒或弱毒株与强毒株表型差异的原因之一。
3.3R.anatipestifer毒力因子间可能存在协同作用和网络调控依据报道,可将R.anatipestifer的毒力因子分为3种类型:一是分泌型毒力因子,即分泌系统分泌的毒力因子;二是细菌表面毒力因子,即菌体细胞表面组成成分,包括夹膜、菌毛、生物膜,以及锚定在细菌表面的大分子蛋白等;三是代谢调控型毒力元件,这些调控元件包括能量代谢调控子、应激相关调控子、细菌群体感应信号分子、细菌单元与双元调控系统、细菌毒力因子翻译后修饰元件等[31]。在检测的18个表型中,CAMP、Hema、AMT、CWAH属于第1种类型,OmpA属于第2种类型,Luxr1、Luxr2、TR、TCTR、IS1属于第3种类型,而GAPDH存在于菌体细胞培养介质中和细胞膜外表面,故同时属于第1,2种类型。研究中发现,OmpA、CAMP几乎存在于包含无毒株在内的所有菌株中,且同源性高达95%以上,表明OmpA、CAMP为R.anatipestifer非必需毒力因子;而AMT,不仅在无毒或弱毒株中未被检测到,部分强毒或中强毒株中也未被检测到,其中包括RS菌株,表明R.anatipestifer可能存在可替代AMT的其他毒力因子。LuxR1、LuxR2是LuxR家族蛋白的重要成员,是细菌群体感应系统中一类具有转录调控作用的受体蛋白,主要通过与靶基因的结合或解离,在转录水平表现出激活或抑制的双向调节作用,而TR、TCTR、IS1则是细菌基因组中重要的转录调控元件。尽管本研究还未能找到以上基因在R.anatipestifer致病性方面可能发挥作用的直接证据,但这些基因控制细菌毒力基因的转录表达、细菌生物膜的形成、质粒的结合转移、色素的产生、细菌的运动等是确信无疑的。根据以上结果可推测,R.anatipestifer毒力因子间可能存在协同作用过程,并受到复杂的网络调控,其作用方式和调控机理还要期待在未来的研究中被一一揭示。
综上所述,R.anatipestifer基因组存在遗传多样性,某些菌株在遗传学上相距甚远,这可能与疾病过程本身的突变和选择作用有关。这种突变和选择作用导致某些优势血清型表型基因在R.anatipestifer群体中发生了水平传递或缺失,进而有利于增加那些遗传上相距很远但能表达优势血清抗原的菌株的出现频率与地理分布。同时,R.anatipestifer毒力因子间可能存在协同作用过程和网络调控,这使其致病的分子基础及机制变得更加复杂,单一的基因功能研究对阐明R.anatipestifer感染致病过程和分子机制的意义将变得十分有限。
猜你喜欢
云南化工(2021年6期)2021-12-21
今日农业(2021年11期)2021-08-13
农药科学与管理(2019年6期)2019-11-23
现代园艺(2017年21期)2018-01-03
中国康复理论与实践(2015年10期)2015-12-24
华南农业大学学报(2015年5期)2015-12-04
医学研究杂志(2015年5期)2015-06-10
现代检验医学杂志(2015年5期)2015-02-06
遗传(2014年3期)2014-02-28
世界科学(2014年8期)2014-02-28
