
彩叶桂新品种‘紫嫣公主’遗传多样性的AFLP和SSR分析
2020-08-03 02:26陶秀花罗永松
江西农业学报 2020年7期
陶秀花,罗永松,江 军
(1.江西省农业科学院 蔬菜花卉研究所,江西 南昌 330200;2.江西省赣州市林业有害生物防治检疫局,江西 赣州 341000;3.江西省全南县林业技术推广站,江西 全南 341800)
桂花(OsmanthusfragransLour.)原产于我国西南部,为木犀科(Oleaceae)木犀属(OsmanthusLour)常绿灌木或小乔木,为中国十大传统名花之一[1]。桂花的栽培历史悠久,观赏价值高,应用广泛。桂花在长期的进化过程中受不同环境及栽培方式等的影响,发生了丰富的遗传变异,拥有众多遗传资源。对桂花品种的分类大多是根据形态特征将其分为4个品种群:四季桂品种群、银桂品种群、金桂品种群和丹桂品种群[2-5]。随着长期的自然杂交及人工选育,各品种群形成了丰富多样的栽培种,迄今共记载了170多个桂花品种[6]。利用分子标记技术对桂花品种资源进行分类已经有一些研究报道[7-9]。
‘紫嫣公主’[10]是全南厚朴生态林业有限公司近年来选育的彩叶桂花新品种,通过了木犀属栽培植物品种国际登录中心组织的鉴定,并获得了木犀属新品种国际登录。 国家林业和草原局授予‘紫嫣公主’植物新品种权(2018第17号);2018年10月江西省林木品种审定委员会颁发了品种‘紫嫣公主’良种证书。‘紫嫣公主’的嫩枝、幼叶至成年叶期依次呈现紫红、浅黄、墨绿等多种颜色的变化,彩色观赏期达半年之久,具有很高的观赏价值,可在江西、广西、广东、福建等地作园林观赏栽培。
本研究利用AFLP和SSR标记对彩色桂花新品种“紫嫣公主”的遗传关系进行了鉴定,旨在为种质资源保护和新品种选育提供新的分子依据,并为今后彩叶桂的品种鉴定、保护及新品种开发和资源合理利用提供理论依据。
1 材料与方法
1.1 供试材料
本研究供试材料共有12份(见表1),包括:由全南厚朴生态林业公司选育的两个新的彩叶桂花品种虔南桂妃[11]和紫嫣公主;对照种长叶木犀;由江西农业大学选育的7个栽培品种(丹桂2个、四季桂2个、金桂2个、银桂1个);由江西林业科学院彩叶桂组选育的2份材料(银碧双辉[12]、云田彩桂)。采集各个供试材料的新鲜幼叶,于-20 ℃保存备用。

表1 供试材料的信息
1.2 DNA提取及AFLP分析
根据制造商的建议,使用DNeasy Plant Mini Kit (Takara)从冷冻的叶片组织中提取基因组DNA。AFLP实验过程如Vos P, et al.[13]所述,并做了一些修改;同时用Ecori和msei(Takara)分别对DNA样品进行限制。用双链Ecori-和msei-适配器连接限制性片段。为了减少由实验因素造成的差异,同时进行了消化和结扎。在含300 ng基因组DNA样品、20 μL Ecori、20 μL msei、350 μL T4 DNA连接酶(Takara)、5 pmol Ecori接合器、50 pmol msei接合器和5 μL 10×T4连接酶缓冲液的50 μL体积中,在37 ℃下进行消化连接酶反应,在65 ℃下灭活10 min,并保存于-20 ℃。用上述4 μL digestion-ligation产品与10 μL 2×Taq PCR混合液(Sangon Biotech, Shanghai, China)、20 μL 10 μmol E+0/M+0引物(图1)进行反应。反应进行了35个循环(94 ℃反应60 s, 56 ℃反应60 s, 72 ℃反应1 min),最后在72 ℃下延长5 min。
在用琼脂糖凝胶电泳检测片段涂片存在的前提下,将PCR产物用DDH2O稀释至1∶20,用2 μL进行2个引物的最终扩增,每个引物具有3个选择性核苷酸(E00+3选择基和M00+3选择基)。PCR扩增反应采用原始AFLP协议[13]中描述的触控循环进行。变性PCR产物在6%聚丙烯酰胺凝胶电泳(PAGE)上分离,用标准尺寸标记物在凝胶的第一泳道以120 V加载约3.0 h,用Sanguinetti C J, et al.描述的银染色法[14]进行检测。
1.3 Microsatellite分析
20个Microsatellite先前在O.fragransLour中显示出明显的多态性,故在本研究中使用。PCR扩增10 μL反应体积,包含50 ng基因组DNA、2×taq-PCR主混合物(中国上海桑贡生物科技有限公司)、每种引物0.2 μL。用热循环仪进行放大,样品先在94 ℃变性5 min;然后在94 ℃变性50 s,在55 ℃变性30 s,在72 ℃变性45 s,共35个循环;最后在72 ℃延长8 min,在4 ℃延长。用2.0%琼脂糖凝胶电泳分离PCR产物。
1.4 数据分析
为了描述遗传变异的特征,在物种水平上计算了等位基因的观察数(NA)、等位基因的有效数(NE)、杂合度(HO)、基因多样性(He)和Shannon信息指数(I);还进行了分子方差分析(AMOVA)和主配位分析。使用Genalex 6.1软件,利用遗传距离值进行E分析(PCA)。以相似矩阵为基础,利用NTSYS PC软件中的算术平均(UPGMA)聚类的非加权对群方法构造了树状图。用Dnaman软件进行了核苷酸序列分析。
2 结果与分析
2.1 AFLP和SSR标记的鉴别能力
AFLP通常是1个显性标记。将没有条带扩增出来的记为“0”,扩增出目的条带的记为“1”。在本研究材料中共筛选出55个AFLP引物组合,用这些引物组合进行遗传分析。共发现了1103个片段,分布在100~500 bp范围内(图1),其中多态性片段1018个(占比92.29%,表2);AFLP片段总数从10个(EAT/M-CAG)到34个(EAT/M-CTC)不等,平均18.69个;多态片段从9个(EAT/M-CAG)到32个(EAT/M-CTC)不等,平均为17.79个,其中EAT/M-CTA组合的多态比例最高,达95.8%(表2)。

M:2000marker;1:朱砂丹;2:鄂橙;3:四季桂;4:大叶佛顶珠;5:麸金;6:波叶金桂;7:紫梗籽银桂;8:长叶木犀;9:虔南桂妃;10:紫嫣公主;11:银碧双辉;12:云田彩桂。

表2 桂花中55个扩增AFLP的特性
所调查的17个多态性较高的SSR共鉴定了12份材料中的36个等位基因。生成扩增产物45个,其中多态性占17个SSR的62.2%。PCR产物的大小主要分布在100~300 bp(图2);每个位点的等位基因数平均为2.12个,范围为2~3个(表3)。图2显示了作为代表的清晰的DNA指纹。

表3 桂花17个扩增SSRs的特点

M:2000marker;1:朱砂丹;2:鄂橙;3:四季桂;4:大叶佛顶珠;5:麸金;6:波叶金桂;7:紫梗籽银桂;8:长叶木犀;9:虔南桂妃;10:紫嫣公主;11:银碧双辉;12:云田彩桂。
2.2 遗传群体的特征
基于AFLP标记,将Jaccard系数的两两遗传距离转化为相似系数,结果相似系数的变化范围为0.55~0.82,平均为0.68(表4)。其中银碧双辉与云田彩桂的遗传相似性最大(相似系数为0.82);其次是朱砂丹和鄂橙(0.81)、波叶金桂和鄂橙(0.81)。长叶木犀(木犀科)与其它木犀的配对遗传相似系数为0.55~0.59,平均0.57。‘紫嫣公主’和其他桂花的遗传相似性变化在0.58~0.80之间,其中‘紫嫣公主’和银碧双辉(彩桂)的遗传相似系数最大(0.80)。

表4 12种供试材料间的遗传相似系数
在群体水平上,有效等位基因的数量从0.76到1.11不等,每个位点的平均值为0.91;Shannon指数(I)从0到0.11不等,平均值为0.05(表5)。6个群体间的遗传分化系数(Hst)和基因流(Nm)分别为0.56和0.31(表6),表明在群体间存在遗传分化和基因交换。

表5 不同群体的遗传多样性

表6 5组遗传多样性分析结果
2.3 分子方差分析(AMOVA)
对收集桂花的地点、海拔高度和品种类型的变异模式进行分子方差分析(AMOVA),分析结果表明,总变异量的73%来自种群内部的变异,27%来自种群之间的变异(表7)。外交种通常具有更高水平的遗传多样性和较低水平的种群之间的变异,这与桂花兼容。上述的Hst指数表明,在桂花种群内部存在高度分化,种群内的遗传多样性远远大于地理遗传多样性。

表7 AMOVA分析11个桂花样品的结果
2.4 桂花品种间的遗传关系
为了进一步分析桂花种群间的遗传关系,基于AFLP和SSR的内力系数矩阵构建了UPGMA树状图(图3)。图3显示了基于NEI和LI相似性估计(NEI和LI,1979)的12个样本间的关系,其中AFLP的系数范围为0.57~0.82, SSR的系数范围为0.34~0.93。在该树状图中,所有的材料首先被分为2组,其中长叶木犀(Oleaceae)的材料明显分离,可以认为是离群的。
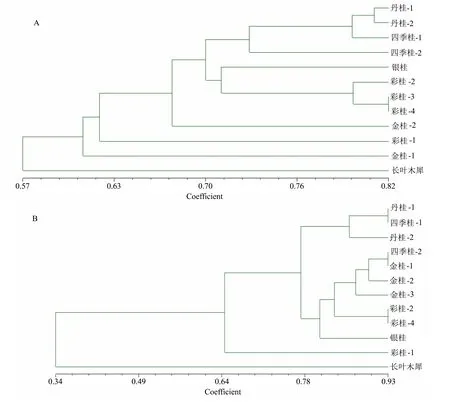
A:基于AFLP数据; B:基于SSR数据。
在AFLP的树状图中,金桂1(麸金)和‘虔南桂妃’分别形成一个类群,另一组在0.67位。另外,当系数为0.72时,将其他材料分为2组:一组由银桂和彩桂组成,包括‘紫嫣公主’;另一组由丹桂和四季桂组成。在SSR的树状图中,银桂、彩桂、金桂、‘紫嫣公主’为一组。
主成分分析结果(图4)显示了11个桂花样本与木犀属之间的遗传关系,与上述聚类分析结果基本一致,表明长叶木犀植物是从木犀属植物中分离出来的。

pop1:丹桂种群; pop2:四季桂种群; pop3:金桂种群; pop4:银桂种群; pop5:木犀属; pop6:彩桂种群。
3 小结与讨论
AFLP被认为是高通量的制造者,结合DNA扩增方法来研究物种水平和群体水平上的遗传多样性(基于地理群体或遗传背景),并在个体的DNA水平上以高分辨率识别变异。我们通过AFLP观察到了92.29%的多态性,这高于Li M, et al.[15]的研究结果;使用SRAP检测了桂花的遗传多态性(83.78%),这略高于Zhang W R, et al.[16]的研究结果;发现了4个桂花物种的遗传多样性(多态性86.8%),表明所有的采样桂花有高水平的遗传多样性。各品种间的片段分布规律表明, ETG/M-CAA引物组合在Albus和黄体之间至少产生了1个独特的片段,该引物组合被认为是非常独特的,因为它可以区分品种。该引物组合在特定品种中产生独特的片段,可用于开发序列标记位点(STS)标记,以识别特定品种。
遗传多样性可以通过检测到的标记数来表达,可以促进种质资源的保护和育种。在本研究中,桂花有效等位基因的数量从0.76到1.11不等,平均每个位点0.91个,低于棉花的[17](范围2~5,平均2.26个);预期的杂合性(He)从0到0.07不等,平均值0.031,明显低于水稻的[18](0.061~0.287,平均0.166)和玉米的[19](0.00~0.35,平均值为0.08)。与这些传统作物相比,桂花在物种水平上的遗传多样性较低,表明通过营养繁殖培育桂花品种,具有较窄的遗传基础,是一个值得关注的问题。因此,迫切需要扩大桂花品种的遗传基础,寻找优质桂花种质资源,培育优良品种。
遗传距离和树形图分析为确定彩叶桂花新品种的起源提供了明确的基础[20]。根据AFLP数据,‘紫嫣公主’和银碧双辉(彩桂组)的遗传相似性系数最大(0.80),说明这两个物种存在遗传相似性。在树形图中,‘紫嫣公主’与‘银碧双辉’(彩桂)、紫梗籽银桂(银桂)相近,表明‘紫嫣公主’属于银桂群。
猜你喜欢
今日农业(2022年15期)2022-09-20
中国农学通报(2022年12期)2022-06-01
中国糖料(2022年2期)2022-04-06
北京联合大学学报(2022年1期)2022-02-12
中国种业(2021年11期)2021-11-25
中学生物学(2019年7期)2019-10-17
生物学教学(2018年3期)2018-08-08
中学生物学(2018年8期)2018-03-01
西江月(2017年4期)2017-11-22
红豆(2017年1期)2017-01-14
