
田中碘泡虫的重描述及系统发育分析
2020-08-07 01:24张金叶刘小花吴明秋赵元莙
水生生物学报 2020年4期
张金叶 刘小花 吴明秋 赵元莙
(重庆师范大学生命科学学院,动物生物学重庆市重点实验室,重庆 401331)
碘泡虫属Myxobolus是碘泡虫科最大的属,迄今已报到900多种[1—4],绝大部分碘泡虫属种类均基于形态学为主体的经典分类学定种,其中许多黏孢子虫种之间拥有极其相似的形态特征[5—8]。然而,随着不断深入的研究显示,尽管一些物种的形态特征已出现一定的差异,但是它们的分子数据实际上依然显示为同种[5,9]。这些结果给黏孢子虫物种的识别和鉴定提出了挑战。因此,迄今分子生物学结合形态学、宿主特异性、组织向性和地理分布的现代分类学方法用于黏孢子虫种鉴定已经被广泛推荐和接受[10—17]。
种群分化在微生物、植物、动物等生物类群相关领域已有广泛的研究[18—20]。关于黏孢子虫的种群研究也有相关报道,例如,2006年Whipps和Kent[9]报道分别在日本、澳大利亚、东太平洋和东大西洋四个主要区域发现的蛇鲭库道虫Kudao thyrsites在形态学和遗传学上已经出现差异。来自美国加利福尼亚州和英国哥伦比亚的小双角小囊虫Parvicapsula minibicornis核糖体小亚基有大于98.9%的相似性,且可以分为15个表型[21]。Gen-Bank中北美和欧洲的脑碘泡虫Myxobolus cerebralis的ITS (Internal Transcribed Spacer) 也有7个碱基的变异[22]。2012年Karlsbakk和Koie[23]报道了来自丹麦和挪威的球形反曲孢虫Sigmomyxas phaerica两个地理种群 (感染相同宿主颌针鱼Belone belone)与来源于丹麦的种群 (感染游沙蚕Nereis pelagic)有着较高的亲缘关系。王茂等[24]对中国重庆、湖北和意大利的透镜碘泡虫Myxobolus lentisuturalis不同种群进行了遗传变异、地理位点、宿主和寄生部位分析,结果表明宿主因素对透镜碘泡虫种群分化的影响大于地理隔离。
田中碘泡虫Myxobolus tanakaiKato,et al.,2017由日本学者Kato等[25]首次从黑川地区鲤Cyprinus carpioLinnaeus,1758 的鳃检获并描述,且提供了18S rDNA分子数据和系统发生学的研究结果。近期我们从中国重庆江津地区金鱼Carassiusauratus auratusLinnaeus,1758 的鳃部检获了田中碘泡虫。由于田中碘泡虫的两个种群地理分布不同,宿主类型也不相同,故这是又一例研究地理隔离和宿主种类差异对黏孢子虫种群分化影响的素材。基于此,本文对其进行了形态学、分子系统学和种群分化的研究,旨在丰富该种的形态和分子数据,以及黏孢子虫种内分化和系统进化研究的基础数据。
1 材料与方法
1.1 样本收集和鉴定
15条金鱼样本于2017年6月从中国重庆江津地区长冲热带鱼良种场 (106°16′E,N29°29′N) 购买,活鱼样本带回实验室后缺氧致死、解剖;而后将从鱼鳃部检获的孢囊转移到载玻片并戳破获得游离孢子。新鲜孢子用蒸馏水冲洗三遍,离心(2000×g)沉淀。虫种鉴定、样本处理和测量均遵循Lom等[26]和赵元莙等[4,27]的方法。
1.2 基因组DNA提取、目的基因扩增、克隆和测序
首先用蒸馏水洗涤从金鱼鳃部获得并保存于95%乙醇中的黏孢子虫样本两次,每次10000×g离心10min除去多余酒精成分;利用DNeasy血液和组织试剂盒 (QIAGEN,Düsseldorf,Germany) 提取洗涤好的样本基因组,提取过程严格按照说明书执行。18S rDNA 引物包括正向引物18E: 5′-CTGGTTGATCCTGCCAGT-3′;反向引物18R: 5′-CTACGGAAACCTTGTTACG-3′。PCR体系25 μL,包括2.7 μL 10× Buffer,2 mmol/L MgCl22.7 μL,0.25 mmol/L dNTP 2.8 μL,每对15 pmol 18S rDNA引物各0.5 μL,15 ng基因组DNA,Taq酶0.2 μL (TaKaRa,Otsu,Japan) 和双蒸水。PCR程序如下: 94℃预变性5min,然后执行35个循环94℃变性45s,60℃退火50s,72℃延伸2min,最后72℃延伸10min。PCR产物通过1.0%琼脂糖电泳和DNA琼脂糖胶试剂盒 (Omega Bio-Tek,Norcross City,GA) 电泳分离和纯化,随后插入到pMD18-T载体 (TaKaRa,Otsu,Japan)。克隆通过ABI Prism 377 DNA Sequencer (Applied Biosystems Inc.,Foster City,California) 完成测序。

图1 田中碘泡虫的成熟孢子及其宿主Fig.1 Mature spores of Myxobolus tanakai and its host
1.3 18S rDNA的二级结构和系统学分析
通过GenBank中BLAST对本研究所获的黏孢子虫序列进行序列同源性比对;18S rDNA使用BioEdit进行手动编辑[28];Muscle3.6[29]软件进行比对;Paup 4.0和Modeltest 3.6用于模型计算;碱基频率为0.2441、0.1858、0.2768、0.2934,碱基替换率为0.9607、3.9297、1.6333、0.5750、5.0662和1.0000,γ分布形状参数为0.4178,不变位点比例为0.2895;使用DNA Star计算遗传距离和变异度;利用RNA Structure 5.2基于自由能最小化进行18S rRNA二级结构预测,参选European Ribosomal RNA database的二级结构模型,使用RNAViz 2.0[30]进行手动调整;选取了与本研究相似黏孢子虫的18S rDNA的三个高变区 (V2、V4和V7区)分析它们的变异度。
系统学分析选用了65条黏孢子虫18S rDNA序列,包括本研究获得的3条和选自NCBI数据库的感染鲤科鱼且与本研究遗传和形态相似黏孢子虫的62条有效18S rDNA序列[4,31—33],用四角库道虫Kudoa quadricomis(GenBank登录号: FJ792721) 和蒜芥库道虫Kudoa alliaria(GenBank登录号: DQ182561)作外群。MrBayes和PhyML 4.0用于构建Bayes树和ML树;Bayes程序共执行1000000代,每100代收集结果,ML树运算100代。最后用MEGA6.0[34]和Adobe illustrator CS6完成系统树的绘制。最后BI和ML树构建所用序列的不一致性利用consel[35]进行最大程度无偏 (AU) 检测。
2 结果
2.1 田中碘泡虫 (Myxobolus tanakai Kato et al.,2017) 重庆种群形态学描述 (图 1和表 1)
孢囊为近圆形,直径约为0.89—92 mm。成熟孢子呈长梨形,壳面观前尖后盾,缝面观未见,孢子壳瓣光滑。两个梨形极囊等大,约占据整个孢子腔体长的2/3;极丝圈数为8—10圈,垂直于极囊纵轴缠绕;两极囊之间的突起位于孢子前端呈大三角形。在孢质中有嗜碘泡,未见黏液泡。孢子的量度
(n=25): 孢子长(14.58±0.54) μm (13.48—15.63 μm),
孢子宽(6.09±0.37) μm (5.01—6.71 μm);极囊长(7.64±0.55) μm (6.20—8.67 μm),极囊宽(2.17±0.33) μm(1.56—3.45 μm)。未见明显病理性特征。
宿主: 金鱼 (Carassius auratus auratusLinnaeus,1758)
感染部位: 鳃丝
感染率: 6.7% (15尾鱼中仅1尾被感染)
采集地: 中国重庆市江津区热带鱼良种场(106°16′E,N29°29′N)
描述标本: No.CQJj-Caa-201707是用甘油 :70%乙醇∶福尔马林 (G∶A∶F= 12∶108∶5) 混合液固定的孢囊,现存于重庆师范大学动物生物学重庆市重点实验室标本收藏中心。

表 1 田中碘泡虫与其他形态相似种的孢子测量比较Tab.1 Comparison of spore measurements of M. tanakai and the morphologically similar species
2.2 田中碘泡虫18S rDNA分子生物学特征
本研究共获得3条分别来自同一尾金鱼鳃丝的3个不同孢囊的田中碘泡虫18S rDNA序列,长度分别是1923、1925和1934 nt (GenBank登录号依次为:MK552413、MH196559和MK552414)。它们之间的序列一致性为 100% 和 99.9%,分歧度为 0.0 和0.1.与日本报道的田中碘泡虫(LC228235和LC228236)一致性为 99.3%—99.6% (表2),分歧度为0.4、0.5、0.6、0.7 (表2)。与其他形态相似种及其种群/株系的一致性和分歧度表现为: 与野鲤碘泡虫(FJ841887、KJ725077、KT240127和FJ710800)的一致性均在 97% 以上 (表2),分歧度为1.8和1.9。与GenBank数据库中形态相似且18S rDNA一致性在 90% 以上的碘泡虫种还包括拟野鲤碘泡虫Mxyobolus parakoi(MH196558)、奥利萨碘泡虫Myxobolus orissae(KF448527)、茄形碘泡虫Mxyobolus toyamai(LC010116)、长孢碘泡虫Myxobolus longisporus(AY364637)、拟茄形碘泡虫Myxobolusparatoyamai(LC228237)、葡萄碘泡虫Myxobolus acinosus(KX810022)、塔形碘泡虫Myxobolus pyramidis(LC228239)、射阳碘泡虫Myxobolus sheyangensis(KU313684)、金布纳碘泡虫Myxobolus ginbuna(LC228238)、吴李碘泡虫Myxobolus wulii(HQ613412)、瓶囊碘泡虫Myxbolus ampullicapsulatus(KC425225) 和洪湖碘泡虫Myxobolus honghuensis(KJ725074),一致性和分歧度详见表2。
田中碘泡虫、野鲤碘泡虫和奥利萨碘泡虫的18S rRNA序列V2、V4和V7可变区的二级结构比较结果显示: 田中碘泡虫MH196559的V2区E10-1较其他田中碘泡虫株系增加了一个内径环;田中碘泡虫LC228236的V7区E43-3与重庆种群及田中碘泡虫LC228235相比缺少了内径环,且该株系的V7区E43-4右端内径环增大。此外,野鲤碘泡虫FJ710800的V4区E23-15、V7区E43-3和E43-4与其他野鲤碘泡虫株系相比,也出现了种内二级结构的差异 (图2)。综上所述,不同种或同种之间在V2区E10-1和V4区E23-15、V7区E43-3和E43-4出现差异。

表 2 田中碘泡虫与其他形态相似种18S rDNA序列一致性和分化度(%)Tab.2 Identity and divergence of 18S rDNA for M. tanakai with its morphologically similar species (%)
2.3 基于18S rDNA的系统学分析
以四角库道虫和蒜芥库道虫为外群分别构建的田中碘泡虫与相关碘泡虫种18S rDNA的ML和BI两棵树的整体趋势基本相一致 (图3),且consel分析建树的最大无偏差分析结果为P<0.05,表明本研究所构建的系统树可用。系统树首先分为两枝: 一枝碘泡虫种的孢子形态呈梨形且前端稍尖,另一枝则为球形或卵圆形且前端钝圆。本研究虫种位于具有梨形孢子虫种的分枝中,3条18S rDNA序列独立聚为一枝,然后与日本报道的2条田中碘泡虫18S rDNA聚枝,最后与野鲤碘泡虫 (FJ710800) 和拟野鲤碘泡虫 (MH196558) 的聚枝形成姊妹枝;而另外的野鲤碘泡虫 (KT240127、KJ725077和FJ841887)株系独立聚枝,并与奥利萨碘泡虫 (KF448527) 形成姊妹枝;而后这两个姊妹枝再聚为一类。
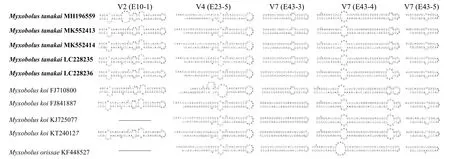
图2 田中碘泡虫及相关种18S rDNA V2、V4、V7二级结构差异区Fig.2 The different secondary structure section of 18S rDNA V2,V4 and V7 for M. tanakai with related species
3 讨论
随着分子生物学技术在黏孢子虫分类学的广泛应用,黏孢子虫的现代分类学相比其经典分类学而言更加精准。近年来已有不少学者利用黏孢子虫18S rDNA进行种内相似度和种间相似度的变化趋势研究黏孢子虫分类[6,36,37]。2006年Fiala[31]的研究结果表明黏孢子虫18S rDNA序列的种内分歧度为0—2%,2008年Ferguson等[38]的研究结果则进一步表明碘泡虫属种内的变异度应低于0.2%,2013年Zhao等[6]认为碘泡虫属种内的变异位点应少于10个。
就形态、宿主、寄生部位而言,本研究获得的3个样本的碘泡虫完全相同,18S rDNA的序列一致性为99.9%和100%,分歧度为0和0.1%,符合Ferguson等[38]和Zhao等[6]报道的碘泡虫属种内变化范围,故确认为是同一物种。形态学特征比较显示,本研究所涉虫种与田中碘泡虫为同种,除田中碘泡虫中国重庆种群的极囊长、宽略小于日本报道的田中碘泡虫原始种群外,其孢子长宽比和极囊长宽比几乎无差别;分子数据分析结果显示,虽然18S rDNA的一级结构出现了碱基差异,一致性为99.3%—99.6%,分歧度为0.4%—0.7%,但依然符合已报道的碘泡虫属18S rDNA种内变化范围[38]。此外,本研究的田中碘泡虫与野鲤碘泡虫、拟野鲤碘泡虫、奥利萨碘泡虫、茄形碘泡虫、长孢碘泡虫、拟茄形碘泡虫、葡萄碘泡虫、塔形碘泡虫、射阳碘泡虫、金布纳碘泡虫、吴李碘泡虫、瓶囊碘泡虫以及洪湖碘泡虫相比,形态上很相似,但均已出现了或多或少的差异,例如田中碘泡虫中国重庆种群的孢子宽明显小于野鲤碘泡虫的日本东京种群、中国湖北种群和美国乔治亚种群 (表1);并且18S rDNA一致性均低于99%,分歧度大于1% (表2),故属于不同物种。采自中国重庆的田中碘泡虫寄主为金鱼(起源于中国的野生鲫驯化形成的观赏鱼类品种,且有丰富的多样性[39]),而检获自日本的田中碘泡虫寄主为鲤;表明两者为不同宿主的地理种群。就二级结构的分析结果而言,虽然本研究所涉的黏孢子虫和来自日本的田中碘泡虫为同一物种,但是无论是中国重庆种群还是日本种群都已经发生了种内分化,且此差异尚未达到新种发生的水平。因此,随后关于田中碘泡虫种内变异驱动力的研究还需使用ITS等高变异度序列进一步分析,其他有关种内变异的驱动力,如宿主和地理分布也需要考虑在内。综上所述,金鱼为田中碘泡虫的新宿主纪录;田中碘泡虫在中国为首次报道,并已发生了可能因宿主和地理分布同时所致的明显的种内分化。
此外,本研究结果还显示GenBank中的野鲤碘泡虫FJ710800与其他野鲤碘泡虫种群相比,18S rDNA序列一致性仅为97.1%和96.7%,分歧度超过2.8 (表2),这已经超出碘泡虫属种内变化幅度[38];且它的18S rDNA的V2、V4、V7区和其他野鲤碘泡虫种群相比均出现了明显差异 (图2),系统发生分析也显示与其他野鲤碘泡虫种群明显分离 (图3),因此,GenBank中报道的野鲤碘泡虫 (FJ710800) 与野鲤碘泡虫(KT240127、KJ725077和FJ841887)可能为不同种,这尚待更多数据进一步研究予以证实。

图3 利用PhyML/MrBayes构建的田中碘泡虫与相关碘泡虫最大似然树/贝叶斯树 (ML/BI树)Fig.3 ML/BI tree of M. tanakai with relative species generated by the software PhyML/MrBayes
猜你喜欢
意林·少年版(2021年6期)2021-05-07
科学(2020年3期)2020-11-26
当代水产(2020年3期)2020-06-15
儿童故事画报(2019年6期)2019-08-01
科技视界(2017年12期)2017-09-11
中国感染控制杂志(2015年7期)2015-12-13
小星星·阅读100分(高年级)(2015年11期)2015-11-28
科学大众(中学)(2015年9期)2015-10-12
少儿科学周刊·少年版(2015年3期)2015-07-07
医学研究杂志(2015年12期)2015-06-10
