
端粒DNA与非编码端粒RNA TERRA在体外形成分子间G-四链体复合物的研究
2020-08-19 02:48何苗,王涛
生物学杂志 2020年4期
何 苗, 王 涛
(1. 中国科学院强磁场科学中心, 合肥 230031; 2. 中国科学技术大学 科学岛分院, 合肥 230026)
端粒是真核细胞染色体末端的核酸蛋白复合体,维持染色体的稳定和完整。正常细胞的端粒长度随细胞分裂次数增长而逐渐变短,被称作细胞分裂的“有丝分裂钟”[1]。在大多数真核生物中,端粒DNA由5~8nt富含连续鸟嘌呤(G-rich)的串联重复序列组成[2-3],可以形成G-四链体结构。该结构由两层或两层以上的G-四分体通过π-π堆积形成,每个四分体又由Hoogsteen氢键将4个鸟嘌呤排列在一个环状平面上所构成。目前,已有研究表明真核生物端粒序列在体内、体外均可形成G-四链体结构[4-7]。该结构可以通过阻止端粒酶和相关蛋白与端粒结合来抑制端粒延伸[8-9],也可以作为端粒加帽结构维持端粒的稳定[6]。
Azzalin等报道端粒可以转录出具有GGGUUA重复序列的非编码端粒RNA TERRA(Telomeric repeat-containing RNA),并定位于端粒[10],推翻了端粒区不能转录的传统认知。 随后相继有NMR和晶体学研究证明,TERRA自身也可以在体内和体外形成G-四链体结构[11-15],其同样可以抑制端粒酶功能,并阻止染色体末端融合和细胞衰老[16]。
G-四链体可以由1条或多条独立的DNA(或RNA)链形成[17],所以在均含有连续鸟嘌呤串联重复序列的端粒DNA和TERRA之间,存在形成DNA-RNA G-四链体复合物的可能性(图1)。近年来已有报道通过点击化学[18]和CD[19]等方法证明人源端粒区DNA和TERRA可以形成(2+2)分子间G-四链体复合物,这种复合物为TERRA提供了一种与端粒DNA相互作用的方式[20]。因此,研究端粒DNA-RNA G-四链体复合物的结构和性质,可以为TERRA与端粒DNA的结合方式以及TERRA维持端粒稳定性的方式提供新的见解。
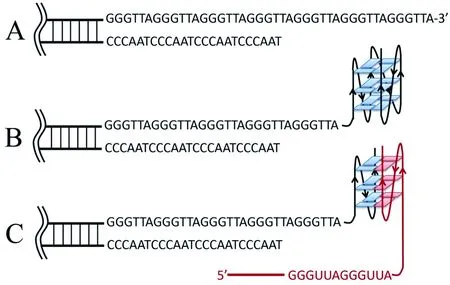
A:3′-悬垂单链; B:DNA分子内G-四链体; C: DNA-RNA G-四链体
除高等真核生物外,我们还以蓝氏贾第鞭毛虫(下称贾第虫)作为低等真核生物代表进行研究。贾第虫是最原始的单细胞真核生物,较其他低等真核生物有更高的进化地位,因此具有更高的研究价值。贾第虫端粒具有与人源端粒序列极为相似的GGGTA重复序列[21],另有生物信息学研究表明其端粒区也可以转录出具有GGGUA重复序列的TERRA[22]。
1 材料和方法
1.1 材料
实验中使用的DNA由Sangon Biotech合成,RNA由TAKARA合成,具体序列如表1。
1.2 方法
所有实验中,K+条件均为20 mmol/L磷酸钾缓冲液(pH 6.8);Na+条件均为含100 mmol/L NaCl 的20 mmol/L磷酸钠缓冲液(pH 6.8)。

表1 寡核苷酸序列
1.2.1 核磁共振实验
将待测试核酸样品溶于含10%重水的磷酸盐缓冲液(pH 6.8)中,95 ℃退火,在Bruker Ultrashield Plus 500 MHz核磁共振波谱仪上扫描一维氢谱。滴定实验时,每次向原体系中加入样品后立即进行一维氢谱记谱。
1.2.2 胶迁移率实验
将核酸样品溶于20 mmol/L 磷酸钾缓冲液(pH 6.8)中,95 ℃退火,用含20 mmol/L KCl的12%非变性胶进行电泳检测。电泳缓冲液为含20 mmol/L KCl的0.5×TBE,100 V恒压电泳2 h。
1.2.3 圆二色光谱测试
在磷酸盐缓冲液(pH 6.8)中配制核酸浓度为40 μmol/L的样品,95 ℃退火,在室温下扫描得到220~320 nm范围内的CD光谱。实验在JASCO J-810 CD谱仪上进行,比色皿光路长度为1 mm,最终谱图扣除相应缓冲液背景。
2 结果与分析
2.1 人源端粒序列形成DNA-RNA分子间G-四链体复合物
G-四链体结构核心部分的鸟嘌呤亚氨基质子(H1)参与形成氢键,在一维氢谱10~12 ppm化学位移范围内呈现出特征谱峰[23],所以通常通过该区域特征峰的出现来判断G-四链体结构的形成。NZ117是含1个重复序列的人源端粒DNA片段,NZR61是含3个串联重复序列的人源TERRA片段(表1)。K+条件下,NZ117在G-四链体特征峰区域内出现3个尖峰,表明其可以形成G-四链体结构(图2-B:Ⅰ);NZR61在该特征峰区域内的谱峰堆叠严重,无单独的尖峰出现,表明其形成多种G-四链体结构(图2-B:Ⅱ);而(NZ117+NZR61)等摩尔混合体系的特征峰区域内新出现了12个尖峰,与作为参照的NZ117和NZR61单独存在时的特征峰分布不同,且参照物特征峰完全消失,表明NZ117和NZR61结合形成了新的分子间G-四链体复合物(图2-B:Ⅲ)。以上NMR实验结果表明,人源端粒DNA和TERRA之间可以紧密结合,形成DNA-RNA G-四链体复合物。
胶迁移率实验发现,(NZ116+NZR61)混合体系的迁移率明显不同于NZ117和NZR61自身的迁移率,表明NZ117和NZR61之间可以形成分子间新结构(图2-A)。胶图中NZR61的条带较宽且边缘不清晰,是由于其形成多种结构而使迁移率不同步(图2-A:lane 1)。胶迁移率实验结果与NMR一维氢谱结果一致,表明人源端粒DNA和TERRA之间的确可以形成DNA-RNA G-四链体复合物。
(NZ117 + NZR61)混合体系的一维氢谱谱峰分布总体较为分散,非常有利于应用NMR进行结构解析,但还需进一步优化谱图。所以我们在288 K~320 K区间进行了变温实验,观察到谱峰的总体分布并未有明显变化但曾经重叠的谱峰可以在不同温度下呈现分离状态,说明NZ117与NZR61形成的DNA-RNA G-四链体复合物在变温区间依然保持着复合物折叠状态,具有一定温度范围内的生理热稳定性,以后的结构解析工作中可以通过调控实验温度降低谱峰指认难度(图2-C)。
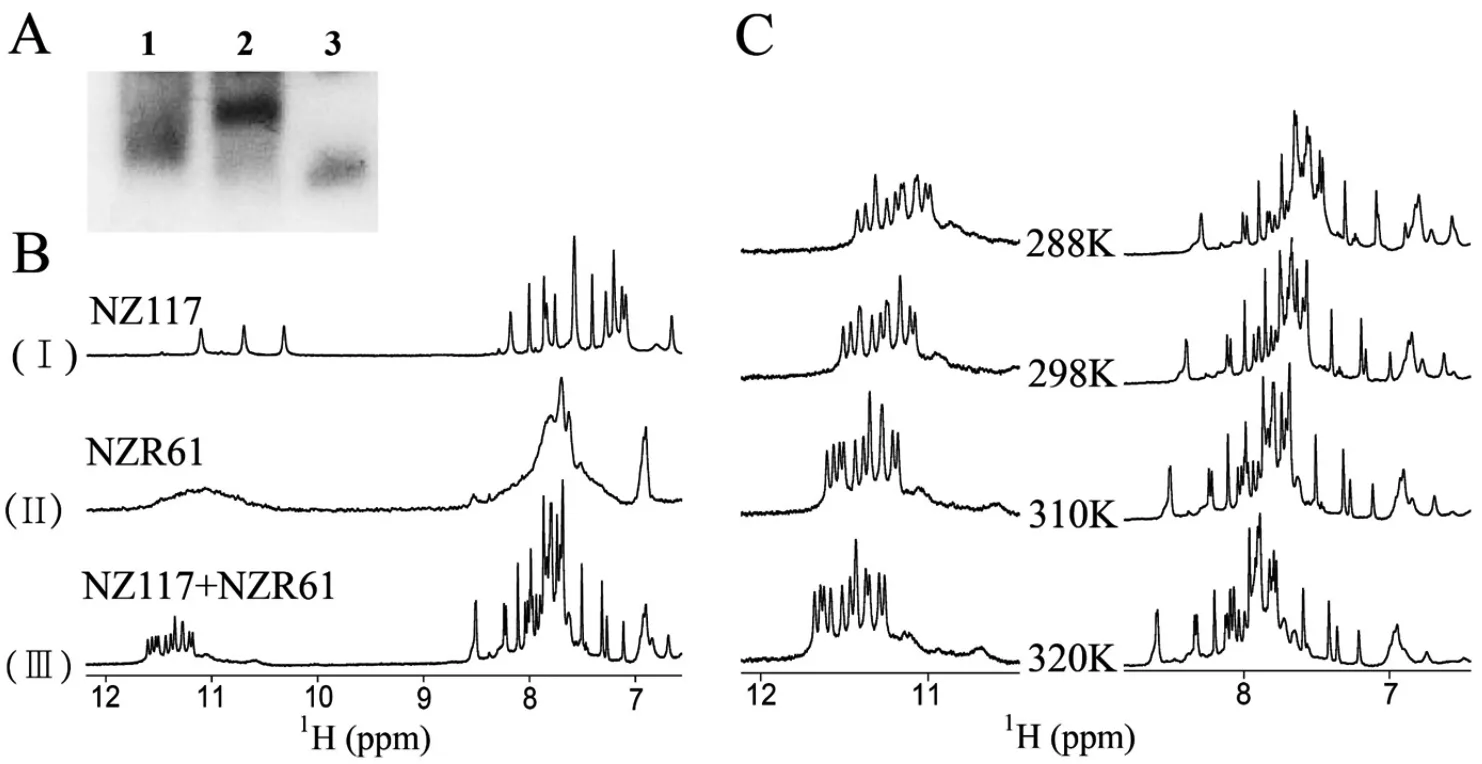
A:人源端粒序列胶迁移率实验结果[泳道1:NZR61;泳道2:(NZR61+NZ117)混合体系;泳道 3:NZ117]。B:NZ117、NZR61和(NZ117+NZR62)混合体系的一维氢谱(6.6~12.2 ppm), 310K。C:(NZ117+NZR61)混合体系的一维氢谱变温实验结果(6.4~9 ppm; 10.4~12.2 ppm)
2.2 贾第虫端粒序列形成DNA-RNA分子间G-四链体复合物
NZ484和NZ116分别是含3个和1个串联重复序列的贾第虫端粒DNA片段,NZR2是与NZ116序列相当的贾第虫TERRA片段(表1)。K+条件下,NZ484的G-四链体结构特征峰区域内无谱峰出现,表明其不形成G-四链体结构(图3-A:Ⅰ);NZ116和NZR2各自特征峰区域内都有明显的3个尖峰,但彼此分布的化学位移不同,表明二者各自均可形成G-四链体结构(图3-A:Ⅱ、Ⅳ)。而在(NZ484+NZ116)和(NZ484+NZR2)两个混合体系中,特征峰区域均有新的尖锐谱峰出现但谱峰分布不同,且与NZ116和NZR2单独存在时的谱峰分布也不相同,表明(NZ484+NZ116)和(NZ484+NZR2)两个混合体系可以分别形成不同的分子间G-四链体复合物,且不同于NZ116和NZR2单独的结构(图3-A:Ⅲ,Ⅴ)。
Na+条件下,NZ484和NZ116均无G-四链体特征峰出现,表明两者均不形成G-四链体结构(图3-B:Ⅰ、Ⅱ);而(NZ484+NZ116)混合体系出现了G-四链体特征峰,表明NZ484和NZ116之间可以形成DNA-DNA G-四链体复合物(图3-B:Ⅲ);NZR2单独存在时有4个特征峰分布,表明其自身则可以形成G-四链体结构(图3-B:Ⅳ);在(NZ484+NZR2)混合体系中,有除NZR2 G-四链体特征峰外的新峰出现,说明NZ484和NZR2之间可以形成DNA-RNA G-四链体复合物(图3-B:Ⅴ)。综上所述,贾第虫端粒序列既可以形成DNA-DNA G-四链体复合物,又可以形成DNA-RNA G-四链体复合物。
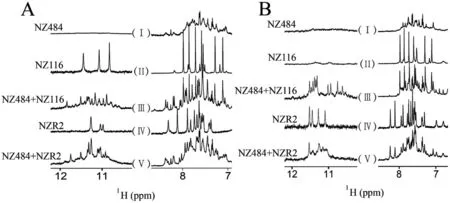
A:K+条件下贾第虫端粒序列片段的一维氢谱(6.9~8.7 ppm;10.3~12.2 ppm)。B:Na+条件下贾第虫端粒序列片段的一维氢谱(6.6~8.6 ppm;10.2~12.2 ppm)。298 K
2.3 RNA诱导平行-反平行混杂型G-四链体复合物向全平行结构转化
CD对核酸的构象变化非常敏感,可以通过220~320 nm范围内的特征峰分辨具有平行或反平行构象的G-四链体结构[24-25],所以我们用圆二色谱对上文中贾第虫端粒序列所形成的G-四链体结构中核酸糖-磷骨架链的相对取向进行研究。K+条件下,NZ116和NZR2样品以及(NZ484+NZ116)与(NZ484+NZR2)两个混合体系的CD谱图均呈现260 nm处的强正峰,而相同浓度的NZ484样品的CD谱图在260 nm处表现出相对弱的正峰。结合一维氢谱实验结果(图3-A),表明NZ484在K+条件下无折叠结构存在,而其它4个样品中形成的G-四链体结构均是采用了全平行的链取向(图4-A)。
Na+条件下,NZ484、NZ116和NZR2样品以及(NZ484+NZR2)混合体系的CD谱图均出现260 nm处的正峰。同样结合一维氢谱实验结果(图3-B)可知,NZ484和NZ116在Na+条件下均不形成G-四链体,NZR2和(NZ484+NZR2)混合体系均形成全平行的G-四链体结构。但是,(NZ484+NZ116)混合体系的CD谱图不仅显示出260 nm处的正峰,在295 nm处还出现了一个正峰,这表明该混合体系在Na+条件下形成了同时具有平行与反平行链取向的混杂型DNA-DNA G-四链体复合物。随后,向(NZ484+NZ116)混合体系中加入NZR2形成(NZ484+NZ116+NZR2)三元混合体系后,295 nm处的正峰消失,表明NZR2可以诱导(NZ484+NZ116)混合体系中已形成的平行-反平行混杂型DNA-DNA G-四链体复合物向全平行G-四链体结构转化(图4-B)。已知NZ484和NZR2可以形成具有全平行构象的G-四链体复合物,所以我们推测,在三元混合体系中, NZR2可以竞争替换掉NZ116与NZ484结合形成全平行DNA-RNA G-四链体复合物。
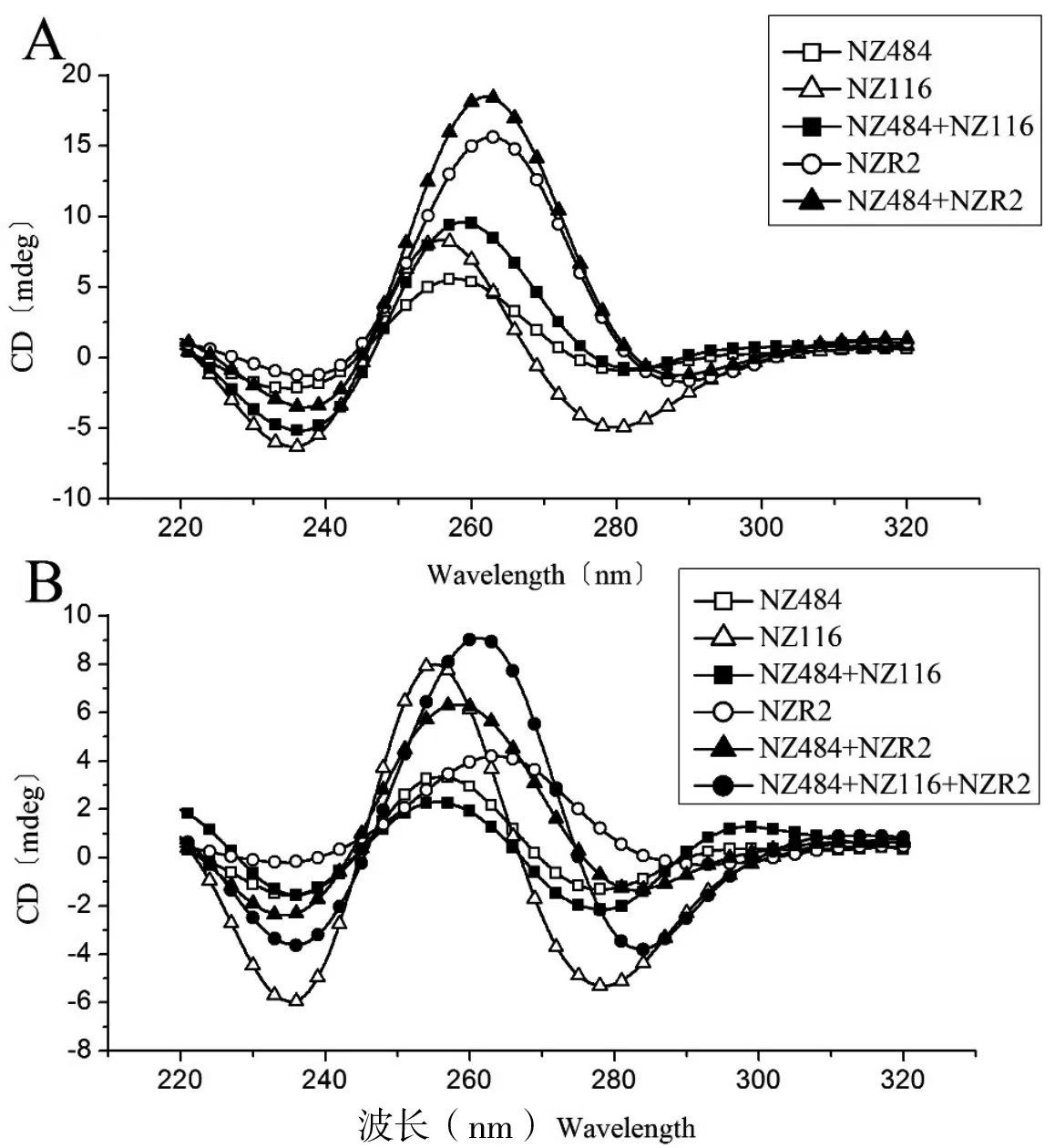
A:K+条件下贾第虫端粒序列片段的CD结果(220~320 nm)。B:Na+条件下贾第虫端粒序列片段的CD结果(220~320 nm)。 298K。□:NZ484;△:NZ116;■:(NZ484+NZ116)混合体系;○:NZR2;▲:(NZ484+NZR2)混合体系;●:(NZ484+NZ116+NZR2)三元混合体系〔在已完成测试的(NZ484+NZ116)混合体系中加入NZR2混匀后直接测量〕
2.4 RNA短链与DNA短链竞争形成更稳定的DNA-RNA分子间G-四链体复合物
为验证猜想,我们进行了滴定实验。在K+条件下,向(NZ484+NZ116)混合体系中逐渐滴加NZR2至过量,新形成的NZ484-NZR2 G-四链体复合物特征峰(图5中以▲标注)逐渐升高后保持不变(图5-A);而原本存在的NZ484-NZ116 G-四链体复合物特征峰(图5中以*标注)逐渐降低到快要消失(图5-A)。表明NZR2可以竞争替换NZ116与NZ484结合,形成新的NZ484-NZR2 G-四链体复合物,且该DNA-RNA G-四链体复合物在此三元混合体系中占多数(图5-A)。另一方面,向(NZ484+NZR2)混合体系中逐渐滴加NZ116至过量时,原本存在的NZ484-NZR2 G-四链体复合物特征峰只是稍有下降;而新形成的N Z484-NZ116 G-四链体复合物特征峰直到NZ116开始过量时才少量出现(图5-B)。表明NZ116滴入(NZ484+NZR2)混合体系后,只能使少量的DNA-RNA G-四链体复合物向DNA-DNA G-四链体复合物转化,过量滴加NZ116之后的三元混合体系中仍是DNA-RNA G-四链体复合物占主要成分(图5-B)。综上可知,NZR2比NZ116更容易结合NZ484形成更稳定的DNA-RNA G-四链体复合物,但两种G-四链体复合物的数量比例在一定程度上受到DNA和RNA相对浓度变化的影响。
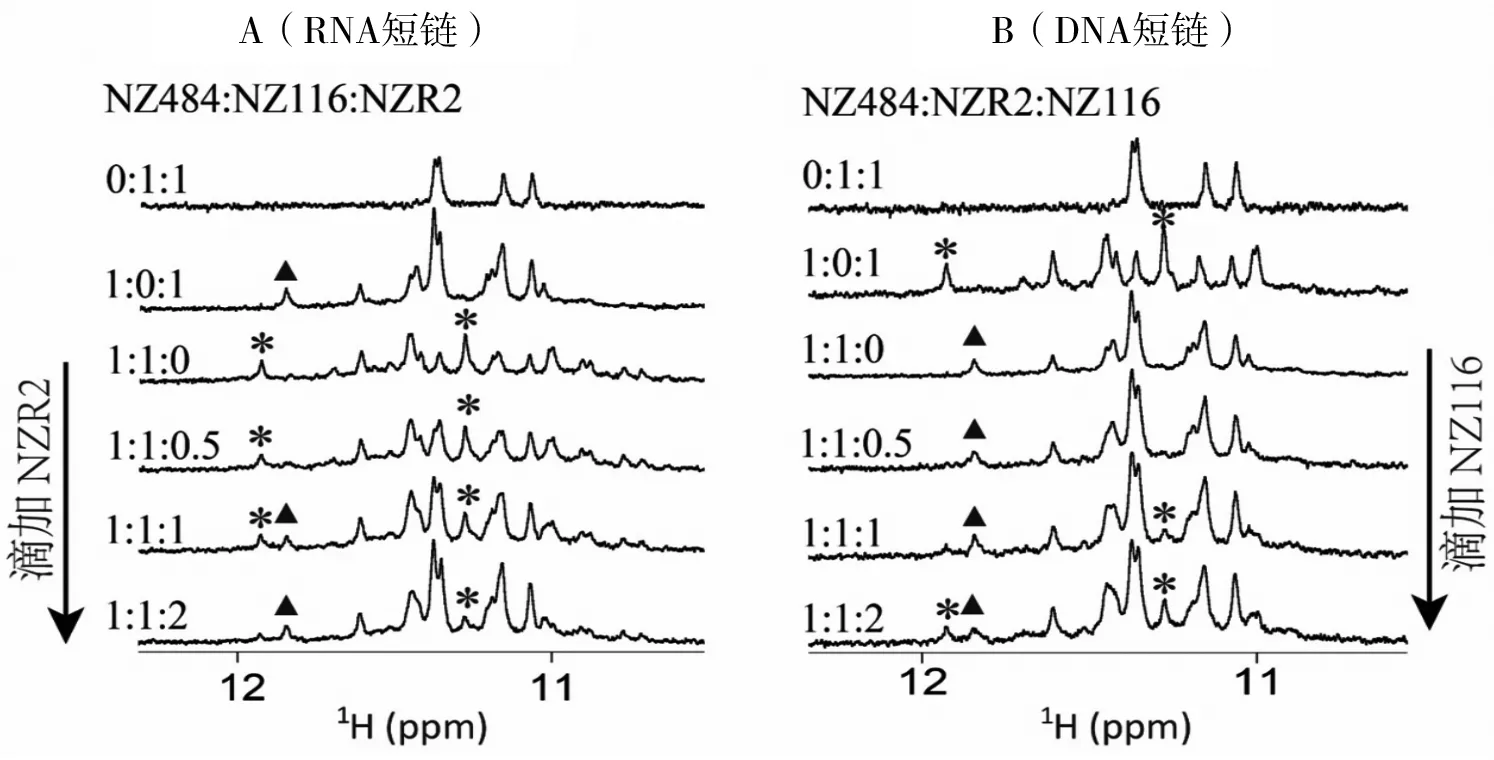
A:向(NZ484+NZ116)混合体系中滴加NZR2的一维氢谱跟踪结果(10.5~12.3 ppm)。B:向(NZ484+NZR2)混合体系中滴加NZ116的一维氢谱跟踪结构(10.5~12.3 ppm)。310K。(NZ116+NZR2)混合体系谱图为参照;▲:NZ484-NZR2 G-四链体复合物的代表性谱峰;*:NZ484-NZ116 G-四链体复合物的代表性谱峰
Na+条件下得到的滴定实验结果与K+条件下一致(图6-A、B),表明NZR2比NZ116更容易结合NZ484形成更稳定的DNA-RNA G-四链体复合物,但同样,两种G-四链体复合物的数量比例也在一定程度上受到DNA、RNA相对浓度变化的影响。结合文CD实验结果证明,在三元混合体系中, NZR2可以竞争替换掉NZ116与NZ484结合,诱导平行-反平行混杂型DNA-DNA G-四链体复合物转化为全平行的DNA-RNA G-四链体复合物。
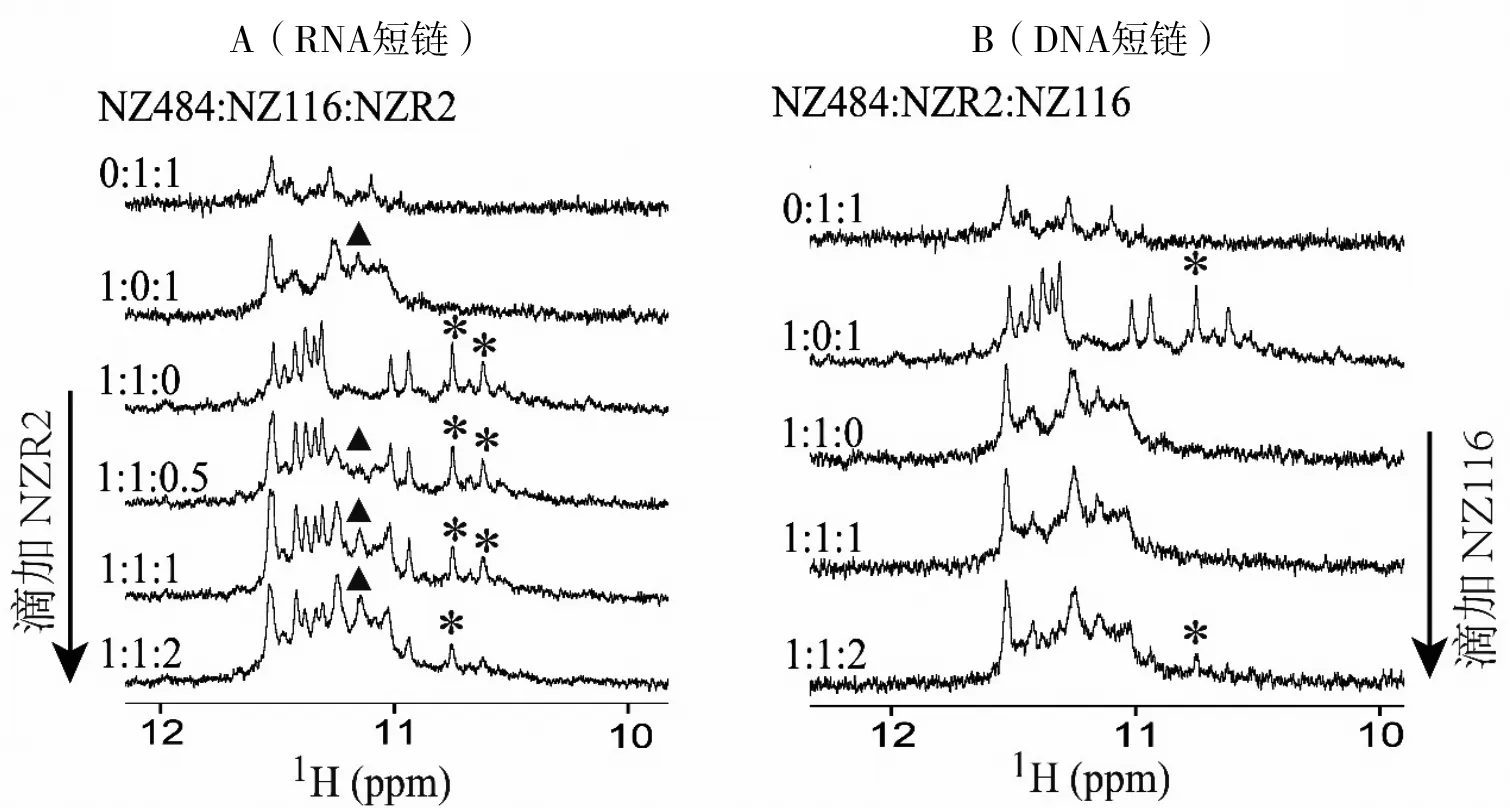
A:向(NZ484+NZ116)混合体系中滴加NZR2的一维氢谱跟踪结果(9.8~12.1 ppm)。B:向(NZ484+NZR2)混合体系中滴加NZ116的一维氢谱跟踪结果(9.9~12.2 ppm)。298K。(NZ116+NZR2)混合体系谱图为参照;▲:NZ484-NZR2 G-四链体复合物的代表性谱峰;*:NZ484-NZ116 G-四链体复合物的代表性谱峰
3 讨论与结论
本文通过NMR检测到了人源端粒序列形成的(1+3)DNA-RNA G-四链体复合物,以及贾第虫端粒序列形成的(3+1)DNA-RNA G-四链体复合物,表明在高等和低等真核生物的端粒序列中均可形成DNA-RNA G-四链体复合物,且此类复合物中DNA与RNA的序列长度可以自由组合,而这种DNA-RNA G-四链体复合物的形成可以阻止端粒酶接近端粒末端而抑制端粒酶对端粒的延伸。值得注意的是,文中人源端粒序列NZ117和NZR61序列形成的DNA-RNA G-四链体复合物(图2),其一维氢谱谱峰分散程度高且具有一定程度上的耐热性,可以通过改变温度使个别重叠的谱峰分散,非常有利于后续的结构解析工作。
此外,在CD实验中,我们发现平行-反平行混杂型DNA-DNA分子间G-四链体复合物可以被RNA诱导形成全平行的G-四链体结构,这可能与RNA G-四链体结构特性有关。由于RNA核糖上的2′-OH可以形成额外的相互作用而对整体结构施加空间限制,使得 RNA G-四链体主要表现出全平行型拓扑结构[13, 26]。因此,在RNA与DNA形成分子间G-四链体复合物时,RNA可能会诱导与之结合的DNA形成全平行的DNA-RNA G-四链体复合物。
最后,通过NMR滴定实验,确定了在混合体系中,RNA短链可以与碱基序列相当的DNA短链竞争性地结合另一DNA长链分子,自发形成更稳定的DNA-RNA G-四链体复合物,但其形成的数量比例在一定程度上受到DNA和RNA相对浓度变化的影响(图5和图6)。在生物体内,端粒RNA浓度随转录和降解过程而频繁变化,这种变化可能使端粒区的结构也随之不断变换:当端粒RNA转录水平较高时,端粒区主要形成DNA-RNA G-四链体复合物;而当端粒RNA降解时,DNA-RNA G-四链体复合物解体,导致与端粒RNA结合的端粒DNA链再被释放出来。这种RNA水平的变化过程可以使端粒DNA在多种结构状态中不断变换,有可能作为端粒在细胞周期不同时期产生状态差异的调控机制。
猜你喜欢
分析科学学报(2021年3期)2021-07-14
色谱(2021年6期)2021-05-06
科技资讯(2020年12期)2020-06-03
空天防御(2020年1期)2020-04-13
中成药(2018年7期)2018-08-04
中成药(2018年3期)2018-05-07
中国现代医学杂志(2018年30期)2018-01-23
中成药(2017年5期)2017-06-13
恋爱婚姻家庭·养生版(2016年9期)2016-09-07
恋爱婚姻家庭·养生版(2016年5期)2016-05-06
