
板栗PPO基因克隆及生物信息学分析
2020-08-28 08:51程丽莉程运河兰彦平曹庆昌胡广隆苏淑钗
西南农业学报 2020年7期
程丽莉,程运河,兰彦平,曹庆昌,胡广隆,苏淑钗
(1.北京林业大学省部共建森林培育学科与保护重点试验室,北京 100083;2.北京市林业果树科学研究院,北京 100093)
【研究意义】板栗为壳斗科(Fagacaea)栗属(Castanea)落叶果树,起源与我国,栽培历史悠久,是我国重要的经济树种。板栗种仁含有大量人体有益的次生代谢物质,如胡萝卜素、没食子酸、原儿茶酸等,其营养保健功效又重新被人们重视。但在加工环节中,这些次生物质伴随栗仁褐变问题发生,严重影响其产品的感官质量和营养价值。【前人研究进展】研究发现褐变与PPO之间的密切关系,国内外许多学者都围绕该酶在基因工程技术方面开展系列研究,试图通过修饰植株中的褐变相关酶类基因表达[1-2],为控制酶促褐变提供可行途径。PPO是由核基因编码的铜结合蛋白酶[3],表现多基因家族性,在生物体酶促褐变中起到重要的作用,体内色素合成的过程中起关键的作用。目前,番茄[4-6]、马铃薯[7-8]、甘薯[9]、烟草[10]、香蕉[11]、葡萄[12]、苹果[13-14]、中国梨[15]、杏[16]、茶树[17]等植物的PPO基因己被克隆,而板栗鲜见研究。【本研究切入点】为此,本项研究开辟新途径从分子水平着手,利用板栗转录组数据库,预测分析栗仁PPO基因结构、编码蛋白、跨膜区结构特性及编码蛋白保守结构域。【拟解决的关键问题】为克隆板栗PPO基因提供重要的序列信息,为深入研究果蔬褐变发生机制奠定基础。
1 材料与方法
1.1 板栗CmPPO基因克隆
取板栗‘YH’品种栗仁组织,立即用液氮冷冻,置于-80 ℃保存,用于RNA提取,植物总RNA提取试剂盒购于Omega公司,利用紫外分光光度计及凝胶电泳检测质量合格。
转录组测序获得长1969 bp的Contig序列,经Blast分析后发现其可能编码多酚氧化酶,根据该序列设计引物,利用Primer Premier5.0软件设计特异引物,并由上海英俊公司合成。正向引物PPO-1F:5′AACACCTTTACACCAAAACCAC3′,反向引物PPO-1R:5′ GGCAAAAGAAAGAACACAACAAT 3′。以RNA反转录cDNA为模板,通过PCR同源克隆PPO基因编码区序列。PCR反应体系为:2×PCR 反应缓冲液12.5 μl,dNTPs 5.0 μl,上下游引物各0.75 μl,cDNA模板1.0 μl,KOD长链高保真酶0.5 μl,ddH2O定容值25.0 μl。PCR反应条件为:94 ℃预变性2 min,98 ℃变性10 s,52 ℃退火30 s,68 ℃延伸2 min,35个循环后,72 ℃延伸8 min。PCR结果凝胶电泳检测,纯化回收,由上海生工测序。
1.2 CmPPO生物信息学分析
核酸及氨基酸序列组成分析、理化性质分析、开放阅读框(open reading frame,ORF)的查找和翻译,利用软件及 Prot-Param、p I/Mw、ORF Finder 等在线工具进行;核酸和氨基酸序列的同源性比对及多序列比对利用 Blast 和 Clustal W 在线工具完成;亲水性/疏水性的分析利用在线工具ProtScale完成;蛋白质二级及三级结构的预测利用SOPMA[18]和 SWISS MODEL[19]等在线工具完成。MEGA6.0软件[20]采用Neighborjoining 法(bootstrap值设定为 1 000)产生系统进化树。
1.3 不同品种不同组织CmPPO的表达
板栗Real-time PCR内参基因引物。扩增体系25 μl:cDNA模板1.0 μl,2×SuperReal PreMix Plus 12.5 μl,正向引物0.75 μl,反向引物0.75 μl,50×ROX×Reference Dye 0.5 μl,补水ddH2O 9.5 μl。Real-time PCR条件:95 ℃,15 min;95 ℃,15 s,60 ℃,60 s,40个循环。以安捷伦3005P检测PPO基因在板栗不同品种、不同组织中的相对表达量。
2 结果与分析
2.1 板栗果实多酚氧化酶序列的获得
以反转录得到的板栗果实cDNA第一链为模板,扩增获得1861 bp的基因序列,通过NCBI的BlastX比对,该片段与多种植物的多酚氧化酶有较高同源性,且片段包含了终止密码子,开放阅读框(ORF)长1794 bp,命名为CmPPO。经过ORF Finder在线分析,编码597个氨基酸(图1)。NCBI Blast p分析板栗PPO氨基酸序列结果显示(图2),本试验克隆的基因编码蛋白属于络氨酸酶基因家族蛋白,具有PPO蛋白典型特征,包含了PPO未知功能的保守结构域(468~594,PPO1_KFDV,pfam12143)、络氨酸普通中心区域(172~380,pfam00264)和多酚氧化酶中间保守域(386~437,pfam12142)。

图1 CmPPO基因cDNA序列及编码氨基酸Fig.1 cDNA sequence and amino acid sequence of CmPPO gene

图2 CmPPO编码氨基酸保守性分析Fig.2 Amino acids conservation analysis for CmPPO
2.2 板栗PPO基因的生物信息学分析
利用在线软件ExPASY预测CmPPO编码蛋白质的相对分子量66 335.38,理论等电点(pI)6.45,属于酸性蛋白总负电荷残基(Asp+Glu)数为72,总正电荷残基(Arg+Lys)数为67。其半衰期在体外哺乳动物网织红细胞为 30 h,在酵母体内大于 20 h,在大肠杆菌体内大于 10 h。不稳定系数为 40.99>40,为不稳定蛋白。脂肪系数77.47,总平均疏水指数(GRAVY)-0.434,为亲水性蛋白(图3)。板栗CmPPO蛋白跨膜区结构TMHMM软件在线分析显示蛋白全部在膜外(图4),不存在跨膜区,不属于膜结合蛋白。

图3 蛋白亲疏水性分析Fig.3 Analysis of hydrophobicity of CmPPO

图4 板栗CmPPO蛋白跨膜区结构预测Fig.4 Prediction of transmembrane structure of CmPPO
蛋白质二级结构是指蛋白质多肽链氨基酸残基借助氢键折叠和盘绕形成的α-螺旋、β-折叠、无规则卷曲以及模体等组件,其中,α-螺旋和β-折叠是最常见的蛋白质二级结构[21]。无规则卷曲和α-螺旋是板栗仁PPO蛋白二级结构的主要结构元件,其次是β-折叠和β-转角。CmPPO二级结构SOPMA预测分析结果表明,α-螺旋(α-helix,h)的比例最高21.22 %,β-折叠(Beta bridge,b)为0,无规则卷曲(Random coil,c)59.13 %,延伸主链(Extended strand,e)15.91 %,β-转角(Beta turn,t)3.85 %。
应用在线三级结构预测软件Swiss-Model (https://swissmodel.expasy.org/),选取其自动建模功能对板栗CmPPO蛋白质的空间结构模型进行同源建模分析(图5),结果发现与苹果络氨酸酶结构(MdPPO1)有72.75 %的序列一致度,53 %的序列相似度,从126~597位置的氨基酸84 %相匹配。

图5 板栗CmPPO蛋白质三级结构预测Fig.5 Tertiary strcture of CmPPO in chestnuts
2.3 板栗PPO基因家族保守结构域分析及系统进化树构建
板栗CmPPO基因编码的氨基酸序列与其他物种PPO基因编码的氨基酸序列进行Blast比对,发现CmPPO氨基酸序列与其他已知的9种植物的PPO氨基酸序列一致性在41.13 %~68.43 %(表1),其中与蔷薇科苹果PPO氨基酸序列一致性达到68.43 %,与菠菜PPO氨基酸序列的一致性最低,仅为41.13 %。
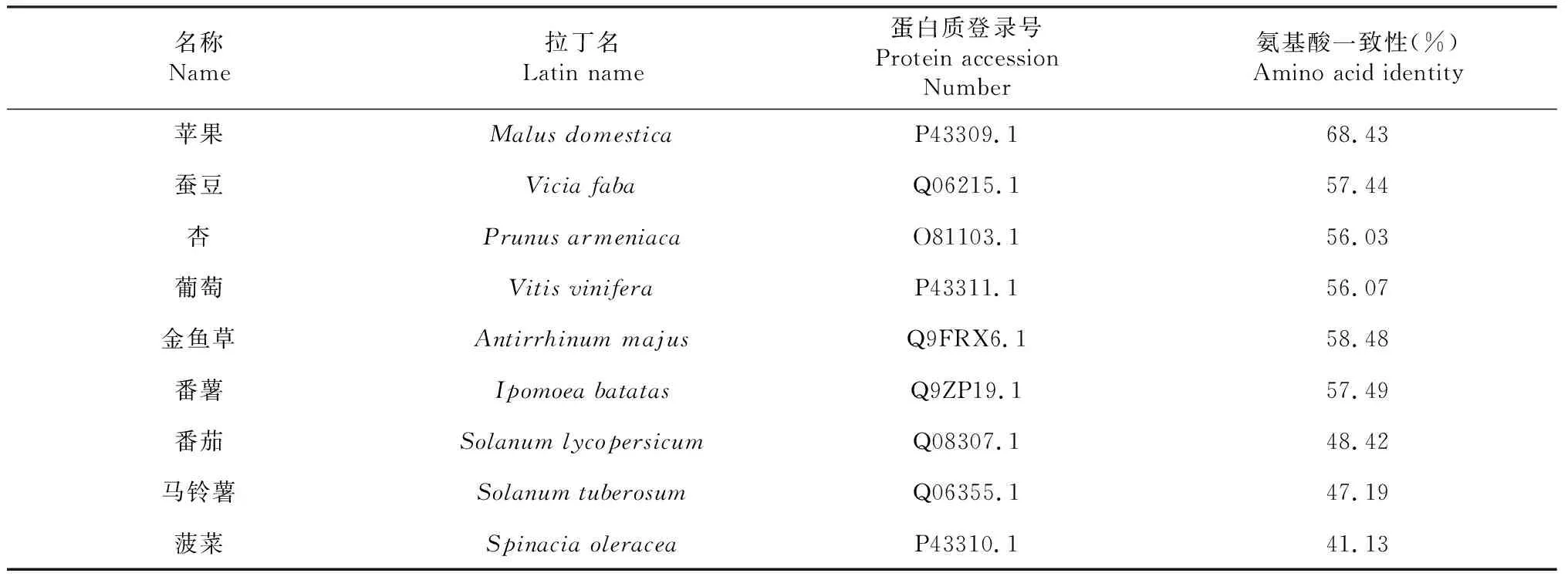
表1 板栗PPO基因与其他9种植物的PPO基因推导的氨基酸序列相似性比较
为进一步研究不同物种间的PPO蛋白进化关系,运用MEGA6.0软件对板栗等17种植物的PPO蛋白序列进行系统进化树分析(图6),聚类结果显示,板栗CmPPO与核桃先聚在一组,两者亲缘关系最近,可能具有相似的进化过程和生物学功能;后与龙眼、荔枝、橄榄一组聚在一起;而与山茶、葡萄、杏、番薯、番茄等物种的亲缘关系较远。
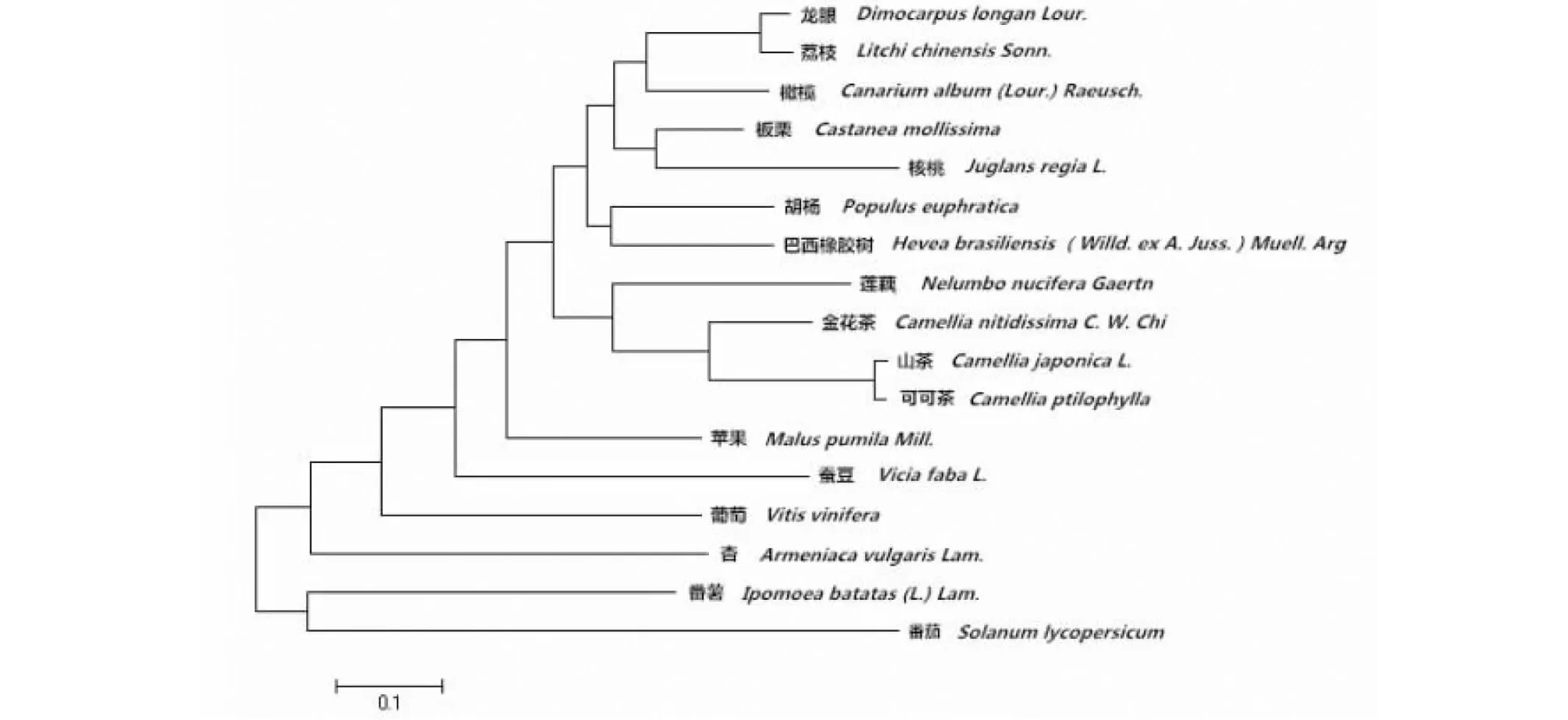
图6 PPO基因系统发育树Fig. 6 The phylogenetic tree of PPO gene
2.4 板栗CmPPO基因在不同品种中表达分析
为分析CmPPO基因在板栗不同品种中表达模式,选取褐变度从重到轻的‘YH’、‘YY’和‘YG’3个品种种子胚乳(B)和胚芽(P)进行qRT-PCR。结果表明,CmPPO基因相对表达量在不同的板栗品种间、果实不同组织器官间存在差异,‘YG’品种不管是在胚乳还是胚芽中表达都是最高,推测可能栗仁PPO基因与其代谢活跃程度、发育阶段、抗逆性等存在密切关系,而且栗仁褐变也不是单基因控制,而是多基因、多种代谢途径共同参与作用的复杂结果,有待后续试验验证。
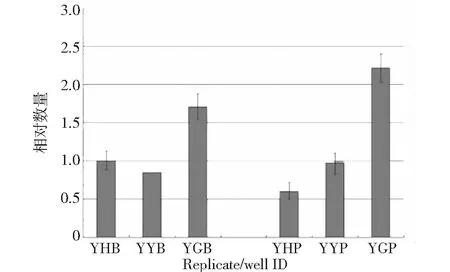
图7 PPO基因在板栗不同品种、不同组织中表达分析Fig.7 PPO gene expression level of different varieties and tissues
3 讨 论
多酚氧化酶PPO是板栗果实和其他果蔬加工时酶促褐变中酚类和单宁等成分氧化而产生的颜色变化酶源。PPO表现为多基因家族性,而本研究通过转录组测序只获得1个功能注释的PPO基因序列,则其PPO基因家族是否还存在其他成员,有待进一步研究发现。
对于板栗仁为材料获得的PPO全长序列,本研究利用多种生物信息学方法对其基因序列和编码蛋白进行预测和分析。保守区域CmPPO氨基酸序列中,172~380区域与络氨酸普通中心区域同源,在386~437和468~594区域分别具有PPO的2个结构域,具有PPO蛋白的典型特征。CmPPO基因编码的蛋白是非跨膜蛋白,属于亲水性,含有一个与2个铜离子相结合的中央域络氨酸酶等主要功能区,这与其他物种PPO蛋白特征高度相似[22]。PPO取自完整板栗仁,二级结构中α-螺旋含量较低,不含β-折叠,而无规则卷曲含量较高,表明酶活性不高,可能与板栗贮藏时间有关系。
在同源性分析与同科栓皮栎(GenBank登录号 XP_023923556.1)序列PPO一致性高达96.48 %,与蔷薇科同样存在褐变问题的苹果一致性68.43 %,与藜科草本植物菠菜一致性最低41.13 %。编码氨基酸序列比对进化树分析,与胡桃科核桃亲缘关系最近。通过qRT-PCR验证,CmPPO基因表达丰度存在品种差异,但在不同板栗品种、果实不同组织中表达差异与果实褐变度不一致,在弱褐变品种中表达却较高,可能是多酚氧化酶是抗性蛋白[23],参与种子贮藏过程其他代谢活动,表达量受淀粉、糖等代谢影响,由此也说明褐变不是单基因控制,而是多基因、多种代谢途径共同参与作用的结果,为在基因水平上研究果蔬褐变机制提供数据线索。
4 结 论
板栗果实多酚氧化酶基因开放阅读框(ORF)长1794 bp,编码597个氨基酸,属于络氨酸酶基因家族蛋白,是非跨膜蛋白,属于亲水性,含有一个与2个铜离子相结合的中央域络氨酸酶等主要功能区,与蔷薇科同样存在褐变问题的苹果一致性68.43 %,编码氨基酸序列比对进化树分析,与胡桃科核桃亲缘关系最近。
猜你喜欢
农产品加工(2022年15期)2022-09-26
茶业通报(2021年4期)2022-01-21
水产科学(2021年4期)2021-07-21
快乐作文(1.2年级)(2021年12期)2021-03-21
渔业致富指南(2020年19期)2020-11-10
农产品加工(2020年7期)2020-05-18
含笑花(2019年6期)2019-12-09
小学生作文(中高年级适用)(2018年5期)2018-06-11
漫画月刊·哈版(2016年5期)2016-07-11
农产品市场周刊(2015年36期)2015-10-30
