
花椰菜苗期黑腐病抗、感品种转录组差异表达分析
2020-08-31 08:44姚玉荣霍建飞郝永娟贲海燕王万立
山东农业科学 2020年7期
姚玉荣,霍建飞,郝永娟,贲海燕,王万立
(1.天津科润农业科技股份有限公司/蔬菜种质创新国家重点实验室,天津 300384;2.天津市植物保护研究所,天津 300384)
花椰菜(Brassica oleraceaL.var.botrytis)是十字花科芸薹属甘蓝的变种,以花球为食用器官的蔬菜。花椰菜除含有多种营养物质外,还含有类黄酮、硫代葡萄糖苷、黑子芥酶等生物活性成分[1],深受广大消费者喜爱。据2018年FAO(联合国粮食及农业组织)报告显示,全世界花椰菜的栽培面积约141.78万公顷,中国为54.79万公顷,占世界总栽培面积的38.64%[2],现已成为世界上花椰菜种植面积最大的国家。
黑腐病是由野油菜黄单胞菌野油菜致病变种Xanthomonas campestrispv.campestiss引起的细菌性病害[3],也是制约花椰菜生产的主要病害之一,发病率为20% ~30%,严重时可达50% ~60%[4]。目前,黑腐病防治方法主要有农事操作如轮作、化学防治和选育抗性品种等。其中,最经济有效的措施为培育和利用抗病品种。由于尚未找到有效抗原,前期研究对黑腐病抗病基因的挖掘和鉴定进展缓慢。随着分子生物学技术的发展和应用,RNA-Seq技术现已广泛应用于基因差异表达研究[5-7]。Chu等[8]利用RNA-Seq技术从抗、感白菜植株中鉴定出2 000多个呈现差异表达的基因,它们主要在茉莉酸和乙烯信号通路、胼胝质沉淀及吲哚类物质合成等过程中发挥作用,且均在抗病植株中表达水平显著上调。Chen等[9]通过对大白菜根肿病抗病和感病品种转录组进行测序和分析发现,抗、感品种间存在151个抗病相关差异基因,主要参与PAMPs、效应子识别、激素信号转导和细胞壁修饰等过程。Zhang等[10]对不同根肿菌侵染时期的青花菜转录组进行比较分析发现,抗、感品种间在NBS-LRR基因、SA代谢相关基因、JA代谢相关基因、细胞壁重塑基因、几丁质酶基因等存在表达差异,这些基因参与寄主对病原菌抗病响应途径,从而使抗、感品种呈现不同抗病性。郭珍等[11]对接菌后油菜根肿病抗、感品种基因进行差异表达分析发现,它们分别存在6 607个和2 499个差异表达基因。雷阳等[12]对114份辣椒群体进行抗疫病鉴定,并对筛选得到的1份抗病材料和3份感病材料进行RNA-Seq测序分析,发现三组材料中有331个差异表达基因,包括68个上调基因和263个下调基因。
目前,多基因控制的抗性机制的研究在拟南芥中较为成熟[13],且相关研究发现,十字花科黑腐病抗性是多基因控制的数量性状[14]。因此,利用转录组分析来研究十字花科黑腐病抗性机制具有可行性。基于此,本研究利用RNA-Seq技术测定并分析花椰菜黑腐病抗、感品种间抗病相关基因的差异表达,以期为后续抗病基因的挖掘及花椰菜抗病育种提供参考。
1 材料与方法
1.1 供试材料
供试品种:花椰菜黑腐病感病品种Y1-2和抗病品种EC-247,均由天津科润农业科技股份有限公司蔬菜研究所提供。
供试菌株:黑腐病菌为野油菜黄单胞菌野油菜致病变种Xanthomonas campestrispv.campestiss,由天津市植物保护研究所蔬菜病害实验室分离保存。
1.2 样品制备
试验于2019年4月在天津市植物保护研究所实验室进行。育苗基质为蛭石∶草炭∶土壤=1∶1∶2(体积比),灭菌后备用。将花椰菜黑腐病抗、感品种的种子分别用50℃热水处理10 min后播于育苗穴盘内,置于防虫网室培养。加强栽培管理,使幼苗生长健壮、整齐一致。
接种菌液的准备:将黑腐病菌菌株转接于LB液体培养基上,置于28℃摇床恒温培养24 h,加适量无菌水稀释,并用细菌浊度仪调整菌液浓度至每毫升1×108个菌体,备用。
于各幼苗长至4~5片真叶时移到人工气候室内保湿24 h,使其叶缘出现露珠。参照姚星伟等[15]的方法进行接种:将制备接种菌液用小型喷雾器进行喷雾接种,均匀喷至幼苗叶片。幼苗接种后培养条件为28℃、相对湿度80%,每天光照14 h。
接种5 d后观察发病情况,同时用无菌剪刀剪下叶片并迅速置于液氮中,再将其移至-80℃保存备用。每品种设置3次重复,每重复15株幼苗。
1.3 RNA提取、测序及组装
将抗、感品种的每个重复混合磨样,用Trizol试剂提取总RNA,利用琼脂糖凝胶电泳检测RNA的完整性和纯度,NanoDrop 2000检测RNA浓度,合格后送至上海美吉生物科技有限公司进行Illunmina测序。
利用Trinity软件对6个样本的clean reads从头组装,用于后续分析。
1.4 差异表达基因分析及注释
利用RSEM 软件将每个花椰菜样本clean reads比对到参考序列,每个基因表达水平用FPKM值计算,有多个转录本的基因,选择最长转录本计算FPKM值。使用DESeq2软件筛选2个样本间的差异表达基因(differential expressed genes,DEGs)。为减少假阳性,对P-value值进行矫正,矫正后P-value<0.05,|log2FC|>1设为基因差异显著表达阈值。
所有表达基因基于6个数据库进行注释,包括:NR(non-redundant protein sequence database)、Swiss-Prot、Pfam、COG(clusters of orthologous groups)、GO(gene ontology)、KEGG(kyoto encyclopedia of genes and genomes)。对于具有多个匹配蛋白序列的基因,选取相似性最高的蛋白质作为最优注释。
1.5 数据分析
利用RSEM软件计算抗、感品种生物学重复之间的Pearson相关系数,并绘制基因表达矩阵图对样本间基因表达相关性进行分析。利用DESeq2软件绘制差异表达基因火山图。
2 结果与分析
2.1 接种黑腐病病菌后叶片发病情况
由图1可以看出,黑腐病病原菌悬浮液接种5 d后,感病品种Y1-2叶片表现出黑腐病典型的“V”形黄褐色病斑,而抗病品种EC-247叶片未呈现该典型病斑。
2.2 转录组质量分析
对花椰菜感病品种Y1-2及抗病品种EC-247接种5 d后的样本进行测序后,本研究构建了6个cDNA文库。过滤后获得49.01×106~55.39×106个平均片段为150 bp的clean reads,GC含量为47.15%~47.80%,Q≥30占比为93.63%~95.16%(表1),表明测序数据质量较好,可以用于后续分析。
2.3 样本间基因表达量分析
花椰菜抗、感品种间基因表达量相关性的聚类分析结果(图2A)显示,抗病品种EC-247和感病品种Y1-2样本各自聚在一起,且表达差异明显。试验中EC-247 5DAI_1~3(E_5DAI)和Y1-2 5DAI_1~3(Y_5DAI)的unigenge数量分别为24 227、25 436个,6个样本共表达的unigene数量为22 570个(图2B)。
2.4 基因表达差异分析
由图3可以看出,花椰菜黑腐病抗、感品种差异表达基因共3 195个,其中上调表达基因1 424个,占差异表达基因的44.57%;下调表达基因1 771个,占差异表达基因的55.43%。因此,抗、感品种差异显著性强的基因可能与花椰菜黑腐病抗病性相关。

图2 抗、感病花椰菜品种间基因表达量相关性分析(A)和基因表达韦恩图(B)

图3 花椰菜黑腐病抗、感品种转录组表达量差异统计(A)和差异表达基因火山图(B)
2.5 差异表达基因的功能注释与富集分析
由图4可以看出,GO数据库中,抗、感品种在接种黑腐病菌5 d时,差异表达基因被富集到3个GO分类,分别是细胞组成、分子功能和生物代谢。差异表达基因富集数目最多的功能是细胞组成,其中以细胞膜(membrane)、细胞(cell)和细胞膜部分(membrane part)数量最多,占比为55.92%;生物代谢分类中基因富集数目最多的功能是细胞过程、代谢过程以及单有机体过程;分子功能注释基因主要包括结合(binding)749个基因,占比46.55%、催化活性(catalytic activity)606个基因,占比37.66%。

图4 花椰菜黑腐病抗、感品种差异表达基因GO功能注释
将花椰菜黑腐病抗、感品种差异表达基因与KEGG数据库进行比对,挖掘抗病和感病品种中差异基因显著富集的KEGG通路(图5)。其中上调表达基因富集到94条通路,显著富集的主要包括苯丙素生物合成(phenylpropanoid biosynthesis),N-聚糖生物合成(N-glycan biosynthesis),亚油酸代谢(linoleic acid metabolism),二苯乙烯、二芳庚烯和姜辣素的生物合成(stilbenoid,diarylheptanoid and gingerol biosynthesis),苯丙氨酸代谢(phenylalanine metabolism),角质、软木脂和蜡质生物合成(cutin,suberine and wax biosynthesis),氨基糖和核苷酸糖代谢(amino sugar and nucleotide sugar metabolism),类固醇生物合成(steroid biosynthesis)和光合作用(photosynthesis)共9条通路(图5A)。下调基因富集到87条通路,显著富集的有错配修复(mismatch repair)、DNA复制(DNA replicatioin)、核苷酸切除修复(nucleotide excision repair)、同源重组(homologous recombination)、醚脂类代谢(ether lipid metabolism)、苯丙素生物合成(phenylpropanoid biosynthesis)、植物昼夜节律(circadian rhythm plant)、类黄酮生物合成(flavonoid biosynthesis)、植物-病原互作(plant-pathogen interaction)、植物MAPK信号通路(MAPK signaling pathway-plant)、核黄素代谢(riboflavin metabolism)11条代谢通路(图5B)。
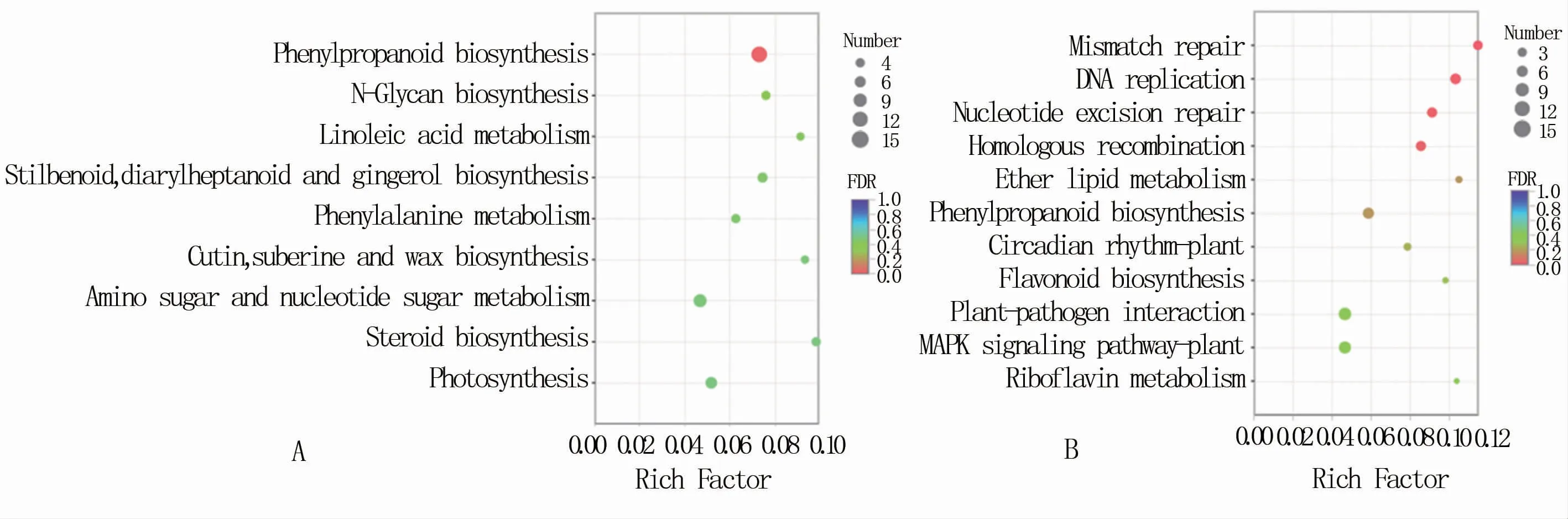
图5 花椰菜黑腐病抗、感品种差异表达基因KEGG通路
3 讨论与结论
本研究利用RNA-Seq技术分析了黑腐病菌野油菜黄单胞菌野油菜致病变种接种5 d后的花椰菜抗、感病品种间基因转录差异,得出抗病品种E_5DAI和感病品种Y_5DAI的unigenge数量分别为24 227、25 436个,6个样本共表达的unigene数量为22 570个;花椰菜抗、感病品种间共有差异表达基因3 195个,其中上调表达基因1 424个,下调表达基因1 771个。通过GO和KEGG进行基因功能注释和显著性富集分析发现,上调表达的基因主要富集在苯丙素生物合成,N-聚糖生物合成,亚油酸代谢,二苯乙烯、二芳庚烯和姜辣素的生物合成,苯丙氨酸代谢,角质、软木脂和蜡质生物合成,氨基糖和核苷酸糖代谢,以及类固醇生物合成和光合作用等途径;下调表达基因主要富集在错配修复、DNA复制、核苷酸切除修复、同源重组、醚脂类代谢、苯丙素生物合成、植物昼夜节律、类黄酮生物合成、植物-病原互作、植物MAPK信号通路和核黄素代谢等代谢通路。该类差异基因主要集中于苯丙烷类代谢、次级代谢产物合成、逆境响应、MAPK信号通路等途径。
苯丙烷类代谢是植物防御反应的主要次级代谢路径之一,参与植保素、木质素、类黄酮、生物碱等与植物防御反应相关物质的合成[16,17],这些物质对真菌、细菌有抑制活性[18-21]。本研究中花椰菜黑腐病抗、感品种差异表达基因在类黄酮生物合成通路显著富集。因此,推测类黄酮途径与花椰菜黑腐病抗病性相关。核黄素能够提高植物抗病性,彭建令[22]发现核黄素能够启动番茄Pto介导的Ptis级联反应,提高对细菌的抗性。此外,蛋白酶类可通过磷酸化和去磷酸化参与调节各种细胞反应,应答环境胁迫[23,24]。
本研究通过高通量测序对花椰菜抗病和感病品种接种黑腐病后基因转录本差异进行了分析,可为在转录水平上全面了解花椰菜抗黑腐病的分子基础提供参考。
猜你喜欢
今日农业(2022年4期)2022-06-01
西南农业学报(2022年4期)2022-05-28
文萃报·周二版(2022年19期)2022-05-13
文萃报·周二版(2022年1期)2022-01-22
今日农业(2021年13期)2021-11-26
中国蔬菜(2021年3期)2021-03-29
贵州农业科学(2020年1期)2020-03-25
发明与创新·大科技(2019年5期)2019-07-31
创新作文(1-2年级)(2016年5期)2016-05-14
北方果树(2016年5期)2016-03-07
