
甘蓝型油菜隐性核三系保持系新材料遗传机理初探
2020-08-31 01:36刘念蒙大庆祝捷
湖北农业科学 2020年10期
刘念 蒙大庆 祝捷

摘要:绵阳市农业科学研究院于1993年发现了油菜(Brassica napus L.)核三系保持系新材料绵7MB,在其自交以及兄妹交后代分离群体中,育性分离比例均不符合孟德尔遗传规律,呈现出偏分离现象。通过对绵7MB田间遗传分析和细胞学分析,对其遗传机理进行了初步的探索。结果表明,偏分离现象可能是由于绵7MB在雄配子传递过程中引起的。绵7MB偏分离现象的产生不是由于减数分裂异常,可能由于部分基因型的合子死亡,导致后代比例出现异常。
关键词:甘蓝型油菜(Brassica napus L.);偏分离;雄性不育;绵7MB
中图分类号:S565.4 文献标识码:A
文章编号:0439-8114(2020)10-0019-04
DOI:10.14088/j.cnki.issn0439-8114.2020.10.004 开放科学(资源服务)标识码(OSID):
Abstract: The new nuclear three-lines maintainer Mian 7MB was revealed by Mianyang Academy of Agricultural Sciences in 1993. In the segregating population of selfing and sibling offspring, the proportion of fertility segregation did not conform to Mendelian genetic law, showing a partial segregation phenomenon. Through the field genetic analysis and cytological analysis, the genetic mechanism of Mian 7MB was preliminarily explored. The results showed that the segregation distortion in Mian 7MB only happened in the male gamete transference. Mian 7MB segregation was not due to abnormal meiosis, but may be due to the death of zygotes of some genotypes, resulting in abnormal proportion of offspring.
Key words: Brassica napus L.; segregation distortion; male sterility; Mian 7MB
油菜(Brassica napus L.)是世界上四大油料作物之一。中国油菜播种的总面积和总产量都位于世界首位,约占世界的30%[1]。油菜具有杂种优势强、繁殖系数高、杂种制种技术简单等特点,因此在油菜上利用杂种优势具有重要的理论和实践意义[2]。目前,中国研究和利用杂种优势的主要途径有细胞质雄性不育、细胞核雄性不育、细胞核+细胞质雄性不育、生态型雄性不育、化学杀雄处理、自交不亲和、天然杂交等方法。在油菜杂种优势利用途径中,细胞核雄性不育具有不育性彻底、稳定,恢复源广,育种周期短,而且不受细胞质不育中恢-保关系的限制,不存在不育胞质的负效应和细胞质单一的风险,容易获得强优势的杂交组合。但是细胞核雄性不育很难获得100%的全不育群体,在繁殖和制种时必须用人工拔除不育系中50%的可育株,严重影响油菜种子生产的成本和质量[3]。如何在母本中经济有效地去除50%的可育株,是细胞核雄性不育可以广泛应用的瓶颈问题,也是世界各国油菜研究单位试图解决的问题。
绵阳市农业科学研究院于1993年起,在自育ms1、ms2类型隐性核两系不育材料转育过程中,发现了绵7MB这一雄性不育保持系,于1998年选育出较好的保持系绵7MB-1[4]。该材料与不育单株的杂交后代中,可以获得80%~90%的不育群体,绵7MB自交后代中不育株的比例可以达到50%~60%,不育株在不同的测交组合和自交后代群体中的比例明显高于1对等位基因的分离比例,不符合孟德尔遗传学规律。本研究通过对绵7MB田间遗传和细胞学分析,分析其育性控制基因及其在后代群体中的分布特点,并结合细胞学观察对绵7MB材料的遗传机理进行了初步探索,以期为这方面研究提供参考。
1 材料与方法
1.1 供试材料
甘蓝型油菜雄性不育两型系材料不育株绵7MA和可育株绵7MB由绵阳市农业科学研究院选育和提供。绵7MB自交后代中有少量单株的自交后代育性比例恢复到3∶1的情况,将这种后代育性比例恢复正常的可育株称作绵7MB*。
1.2 方法
1.2.1 田间育性调查 2014—2016年连续3年对绵7MA×S45B、绵7MB(人工去雄)×S45B、S45A×绵7MB、S45B×绵7MB 4个杂交组合F1代进行田间育性调查,进一步验证绵7MB与S45育性控制基因的遺传等位性。同时对绵7MB自交后代和绵7MA×绵7MB兄妹交后代群体进行育性调查,分析绵7MB育性偏分离现象的一般规律。
1.2.2 花粉活性观察 采用醋酸洋红染色法鉴定花粉育性,于油菜盛花期,上午8:00—10:00采集不同单株上、中、下不同位置即将开放的新鲜花蕾(3~5个),做好标记,用消毒的镊子将花药上的花粉粒置于载玻片上,用解剖针捣碎,加1~2滴醋酸洋红溶液染色,盖上盖玻片,用吸水纸将其周围多余的液体吸干,轻轻压片,置于显微镜下观察。根据花粉染色情况及形态判断其育性:圆形饱满且深红色的花粉粒表示其具有高生活力;低生活力或者不育的花粉粒表现为形态不规则、外形干瘪、畸形而且染色后不能着色或着色较浅。
1.2.3 花藥减数分裂的细胞学观察
1)材料准备。在油菜现蕾之后,花蕾直径为2~3 mm的时候,取整个花序。将材料置于卡诺固定液(乙醇、冰醋酸体积比为3∶1)中固定。隔3~4 h后换新鲜的卡诺固定液,换4~5次(卡诺固定液不再变黄为止),可4 ℃长期保存以备观察。
2)制片及观察。取出油菜花蕾,用去离子水漂洗1~2次。用解剖针或者尖头镊子将花蕾剥开,将花药取出置于1 mol/L HCl中60 ℃水浴2 min,取出后用去离子水浸泡1 min。将材料置于干净的载玻片上,用解剖针或镊子将其捣碎。加1滴卡宝品红染色液染色30 s,然后用解剖针将捣碎的材料均匀涂开,取干净的盖玻片盖上,用滤纸包住盖片,先用手指轻轻挤压盖片去除多余的染色液,然后用吸水纸将其周围多余的液体吸干。将玻片固定于显微镜下观察、拍照。
1.2.4 胚发育观察 分别选取绵7MB自交偏分离群体中植株生长发育状况基本一致的单株,人工套袋授粉,于授粉后7、14、21、28 d后取主花序相同部位的角果,从中间剖开观察。
2 结果与分析
2.1 田间育性调查分析
对绵7MB自交群体和兄妹交群体后代可育株和不育株育性以及花器官形态进行田间调查,发现绵7MB可育株开花期要比绵7MA早5 d左右;可育株花蕾较大,花丝较长,花药饱满,花粉量充足;不育株花蕾空瘪,雄蕊花丝较短,花药在花蕾盛开前就已经退化萎缩,败育彻底,无微粉现象。
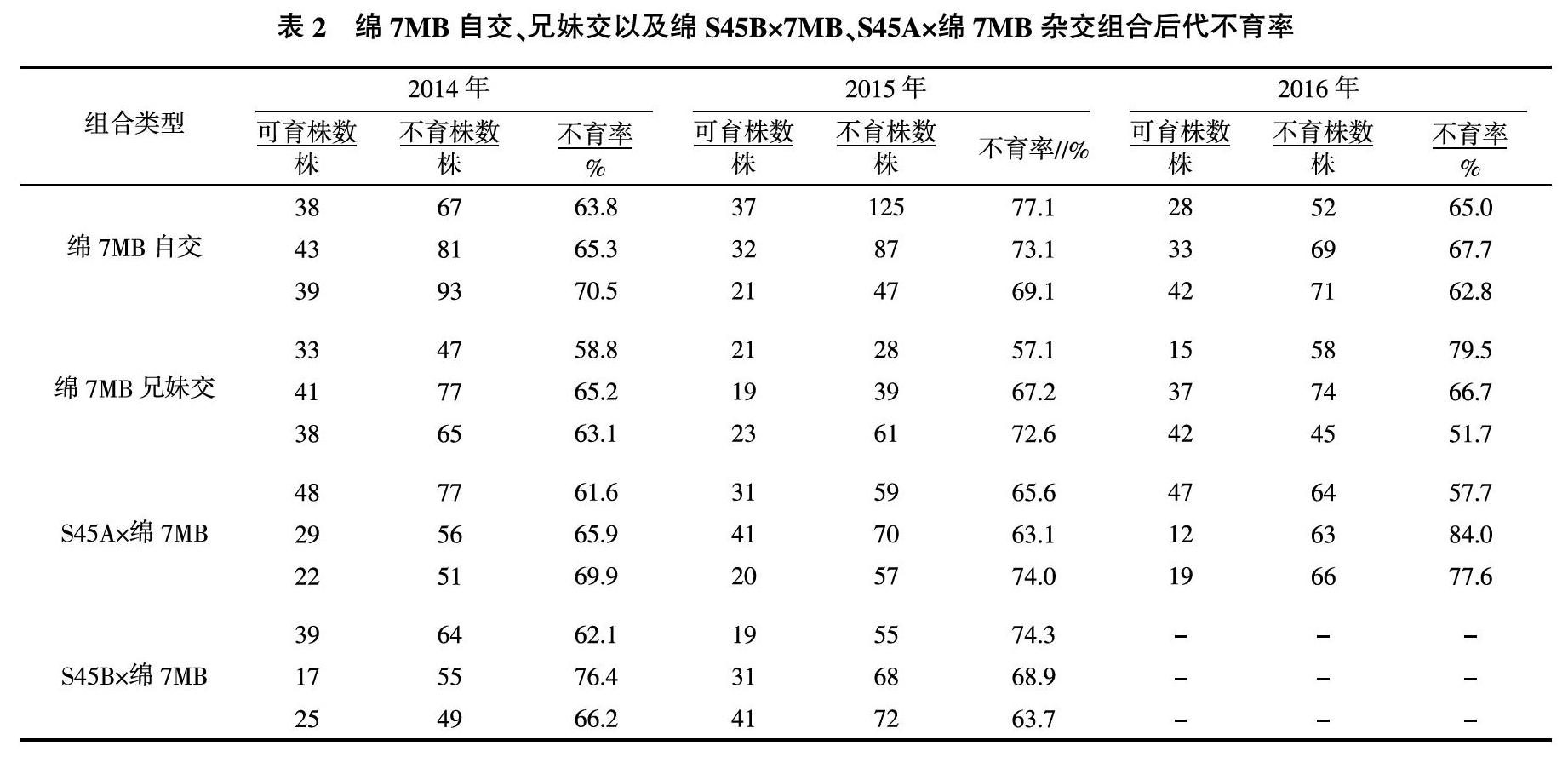
连续对绵7MB自交、绵7MB兄妹交、绵7MA×S45B、绵7MB(人工去雄)×S45B、 S45A×绵7MB、S45B×绵7MB,这些杂交组合的F1代进行育性调查统计。从表1可以看出,绵7MA×S45B、绵7MB(人工去雄)×S45B杂交F1代比例为1∶1和3∶1,符合孟德尔遗传规律,与蒙大庆等[5]的研究结果一致。田间调查结果表明,绵7MB的育性控制基因与S45的育性控制基因可能是由相同的等位基因控制。绵7MB自交、绵7MB兄妹交、S45A×绵7MB、S45B×绵7MB杂交F1代不育率较高,不符合孟德尔遗传学比例,育性遗传呈现出偏分离现象(表2)。
当绵7MB作为母本时,杂交组合后代均没有偏分离现象发生;而当绵7MB作为父本时,后代群体中就会发生育性偏分离现象。推测偏分离现象可能是由于绵7MB在雄配子传递过程中引起的。
2.2 花粉活性观察
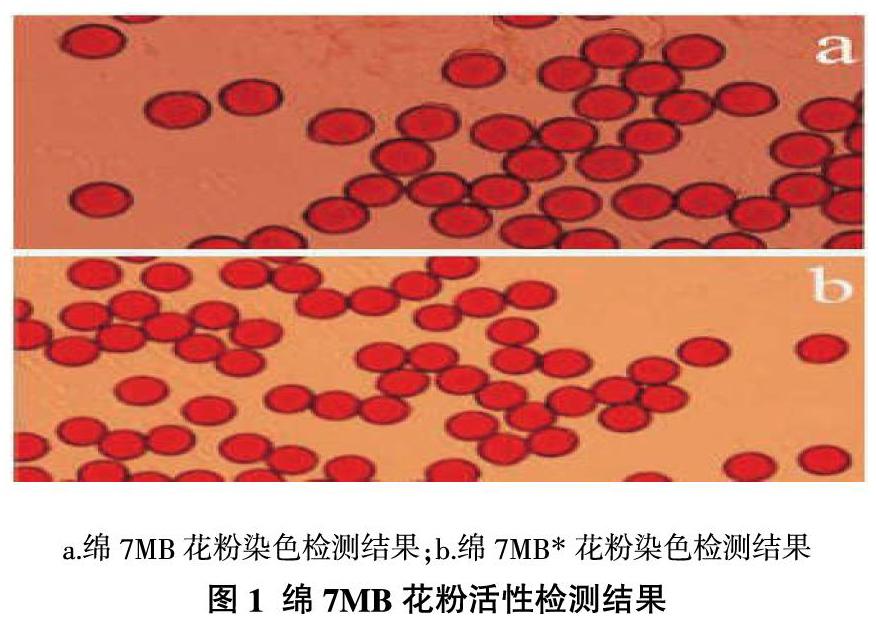
对偏分离材料绵7MB可育株花粉粒进行染色,从图1可以看出,绵7MB与正常植株花粉粒相比,无明显差异,均表现为花粉粒着色较深,花粉粒圆形且饱满。花粉活性检测说明,绵7MB偏分离现象并不是由于部分花粉活性异常引起的。
2.3 减数分裂细胞学观察
对绵7MB雄蕊花药进行减数分裂观察,发现绵7MB减数分裂中同源染色体联会、分离均表现正常(图2),减数分裂过程中无异常出现。说明绵7MB育性偏分离现象的产生不是由于减数分裂异常引起的。
2.4 胚发育观察
从图3可以看出,绵7MB所结角果有明显的死胚现象出现。在受精后14 d,绵7MB角果中部分合子发育明显缓慢;至受精后21 d,生长缓慢的合子开始脱水萎缩;受精28 d后彻底干瘪死亡。统计偏分离材料绵7MB的死胚率发现,与对照相比,绵7MB角果中死胚率较高(表3)。说明胚致死现象可能是导致绵7MB偏分离现象产生的原因之一。
3 讨论
偏分离现象是指在分离群体后代中,某一基因频率或者基因型频率明显偏离经典遗传学比例的现象[6]。偏分离现象的实质是一种选择的过程,包括对配子、合子生活力以及竞争力的选择。偏分离现象作为一种普遍存在的遗传学现象,在小鼠[7]、果蝇[8]、水稻[9]、玉米[10]、小麦[11]等植物中均有发现。但是由于物种来源、遗传作用方式等因素的不同,对偏分离现象的遗传机理研究有待深入。新型核不育保持系绵7MB也呈现出偏分离现象。本研究通过对绵7MB田间遗传进行分析,表明偏分离现象可能是由于绵7MB雄配子传递过程中引起的。对绵7MB细胞学进行分析,表明绵7MB偏分离现象的产生不是由于减数分裂异常,可能由于部分基因型的胚死亡导致后代比例出现异常。对于出现这种偏分离的分子作用机理,还需要进一步研究。
参考文献:
[1] 刘后利.实用油菜栽培学[M].上海:上海科学技术出版社,1987.
[2] 傅廷栋.杂交油菜的育种与利用[M].武汉:湖北科学技术出版社,1995.
[3] 侯国佐.油菜隐性核不育研究与利用[M].北京:科学技术文献出版社,2008.
[4] 蒙大庆,袁代斌,刘 念,等.甘蓝型油菜核三系育种研究与应用[J].技术应用,2012,9(71):51-54.
[5] 蒙大庆,袁代斌,郭子荣,等.双低甘蓝型油菜新型核不育系绵7AB-4的遗传研究Ⅰ.育性表现的初步研究[J].四川农业大学学报,2005,23(1):24-27.
[6] 刘海燕,崔金腾,高用明.遗传群体偏分离研究进展[J].植物遗传资源学报,2009,10(4):613-617.
[7] CASELLAS J,FARBER C R,VERDUGO R A,et al. Segregation analysis of a sex ratio distortion locus in congenic mice[J].J Hered,2010,101:351-359.
[8] MONTCHAMP C M,GINHOUX V,ATLAN A.The Y chromosomes of Drosophil simulansare highly polymorphic fortheir ability to suppress sex-ratio drive[J].Evolution,2001,55:728-737.
[9] REFLINUR,KIM B,JANG S M,et al.Analysis of segregation distortion and its relationship to hybrid barriers in rice[J].Rice,2014,7:3.
[10] WANG G,HE Q Q,XU Z K,et al. High segregation distortion in maize B73C × teosinte crosses[J].Genet Mol Res,2012,11:693-706.
[11] TAKUMI S,MOTOMURA Y,IEHISA J C,et al. Segregation distortion caused by weak hybrid necrosis in recombinant inbred lines of common wheat[J].Genetica,2013,141:463-470.
