
黑龙江地区水稻糙米中γ-谷维素含量和阿魏酸酯组成
2020-10-28 07:14李洪亮张东杰
食品科学 2020年20期
张 超,李 丹,张 昌,李洪亮,殷 坤,张 雪,张东杰,2,
(1.黑龙江八一农垦大学食品学院,黑龙江 大庆 163319;2.黑龙江八一农垦大学 国家杂粮工程技术研究中心,黑龙江 大庆 163319)
大米(Oryza sativa)被全世界三分之二的人作为主食消费,随着世界人口的不断增加,国际稻米研究机构预测到2025年全世界大米市场的消费量将达到8亿 t[1-2]。而在我国,大米的食用人口已超出总人口的三分之二,并且这一比例还在不断上升[3]。
稻米中含有许多对人体有益的天然营养物,如γ-谷维素、普利醇、VE(生育酚、生育三烯酚)、硫胺素、核黄素、烟酸和叶酸等[4]。其中γ-谷维素因最早在米糠中被发现,后得其名“γ-oryzanol”。γ-谷维素是由阿魏酸与三烯萜及甾醇酯化而成的一种天然植物营养物,在米糠油中起着重要的抗氧化作用,目前已有30多种单体结构被确认[5-6]。在天然稻米中γ-谷维素主要由环木菠萝烯醇阿魏酸酯、24-亚甲基环木菠萝烯醇阿魏酸酯、菜油甾醇阿魏酸酯、β-谷甾醇阿魏酸酯组成(结构式如图1所示),约占90%以上。糙米和米糠是γ-谷维素的重要获取来源,其主要作用有抗氧化、抗炎、抗肿瘤、降低胆固醇,并且对神经和更年期起到一定的调节作用[7-8]。我国每年米糠的产量巨大,但只有小部分被开发利用,若在γ-谷维素提取和性质方面加以重视和利用,将会给稻米副产物精深加工带来巨大收益。在体外抗氧化实验中发现,γ-谷维素可以减少植物油在煎炸过程中由高温氧化造成的损失,同时在冷藏过程中可以抑制牛肉胆固醇氧化物的形成[9-10],事实上γ-谷维素能有效避免胆固醇氧化产物对机体造成的损害[11-12]。

图1 阿魏酸酯的结构式Fig.1 Structures of ferulic acid esters studied in this work
稻米中γ-谷维素含量变化受到环境、品种、产地因素的影响,除考察以上因素对稻米中γ-谷维素含量影响外,还有文献报道了不同加工程度对稻米γ-谷维素总含量的影响[13]。不同阿魏酸酯单体的活性也存在明显差别,但是对稻米γ-谷维素单体组成成分的分析相对较少;同时对米糠油中的γ-谷维素含量研究较多,对糙米的研究很少。此外,不同地区开展研究的水稻样本量差异很大,Kim等[14]对韩国不同品种的稻米作了分析,发现γ-谷维素含量差异与基因型有关,另外有文献报道欧洲地区稻米间的γ-谷维素含量,发现存在明显差异[15]。而对我国稻米间γ-谷维素含量的研究几乎为空白,仅徐冉等[16]报道了湖南75 份米糠油中γ-谷维素含量的变化。本实验通过高效液相色谱检测技术得出黑龙江省五大主产区糙米间γ-谷维素含量差异以及分析阿魏酸酯单体组成在自然条件下变化的关系,旨在以实验数据填补对黑龙江区域内糙米中谷维素含量研究的不足,为米制品及稻米深加工提供理论指导。
1 材料与方法
1.1 材料与试剂
实验稻米样品2018年9—10月采自于黑龙江5 个地区农田(方正、五常、响水、查哈阳、建三江);不同成熟期的水稻籽粒2018年7—9月采自宝泉岭农场,每隔7 d采收1 次,采收5 次,品种为龙粳-31(在黑龙江省种植面积最广)。
甲醇、乙腈、醋酸(均为色谱纯) 上海安谱实验科技股份有限公司;γ-谷维素标准品(环木菠萝烯醇阿魏酸酯23%、24-亚甲基环木菠萝烯醇阿魏酸酯51%、菜油甾醇阿魏酸酯15%、β-谷甾醇阿魏酸酯10%) 日本Wako公司;去离子水为GB/T 6682—2008《分析实验室用水规格和试验方法》推荐用水;提取用甲醇为国产分析级。
1.2 仪器与设备
1260 高效液相色谱仪(配有二极管阵列紫外检测器) 美国安捷伦公司;SMART-N纯水机 上海Canrex公司;4N电子分析天平 上海森信实验仪器有限公司;DGG-9123A型电热恒温鼓风干燥箱 上海精桥精密仪器有限公司;CT193旋风式粉碎机 福斯分析仪器(苏州)有限公司;JHSFY-60A水分快速测定仪 无锡市剑辉机电设备有限公司;CPX-3800HC型医用数控超声波清洗器(100 W) 美国超声仪器有限公司;TDZ5-WS台式离心机 湖南湘仪实验仪器开发有限公司;RE-2000B旋转蒸发仪 上海亚荣生化仪器厂。
1.3 方法
1.3.1 样品处理
对采获的样品进行编号,以便区分地区,并对稻米品种进行记录:“地区+具体地点+品种名(龙粳/绥粳/稻花香)+序列编号”。取回实验室后储藏于黑暗阴凉通风处。实验前在烘干箱中45 ℃烘12 h后去壳,挑去带有青色未成熟籽粒后于旋风粉碎机中粉碎过40 目筛收集样品于密封袋中,储藏于4 ℃冰箱中。不同成熟期的水稻,每次采获后,及时用液氮处理后置于-40 ℃冰箱中,冷冻干燥再去壳,粉碎方法同上。
水分的测定:取0.5 g粉于水分快速测定仪中测定水分含量。
γ-谷维素提取[17]:称取约(3±0.10)g样品于50 mL离心管中,加入10 mL甲醇,室温条件下超声60 min后,3 000 r/min离心10 min,取上清液于250 mL旋转蒸发瓶中,50 ℃旋蒸50 r/min至近干,准确加入6 mL甲醇定容,涡旋5 min,将旋转蒸发瓶中的甲醇过0.45 μm有机微孔滤膜装进样瓶,待上机检测。
1.3.2 标准品的制备
取10 mgγ-谷维素标准品用甲醇溶解于10 mL棕色容量瓶中,定容至刻度线处,作为贮存液置于-20 ℃冰箱中;实验时将贮存液稀释成以下不同质量浓度梯度10、50、100、150、200 μg/mL作标准曲线。
1.3.3 色谱条件[18]
色谱柱:C18色谱柱(250 mm×4.6 mm,5 μm);流动相:乙酸-乙腈-甲醇(3∶44∶53,V/V),等度洗脱,流速1.4 mL/min;进样量10 μL;检测波长325 nm。
1.3.4 阿魏酸酯含量计算
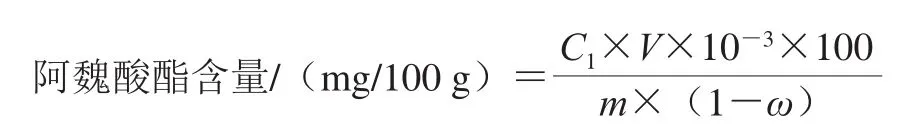
式中:C1为样品的检测质量浓度/(μg/mL);V为甲醇回溶体积/mL;m为样品质量/g;ω为样品含水量/%。
1.4 数据处理与分析
实验所统计的数据利用SPSS分析软件进行处理,采用单因素方差分析(one-way ANOVA)、两两比较均值(Duncan)、相关性分析。P<0.05,差异显著。各组数据均采用±s表示,n=3。
2 结果与分析
2.1 土壤有机质测定结果
为了确定土壤有机质对糙米γ-谷维素含量造成的影响,按照NY/T 1377—2007《土壤pH的测定》测定,采用GB 9834—1988《土壤有机质测定》测定土壤中有机质的含量,不同地区土壤中有机质和pH值有所差别,结果如表1所示,土壤中有机质均值最高的为五常地区,为(4.00±0.57)%;有机质均值最低为响水,为(2.74±0.76)%。
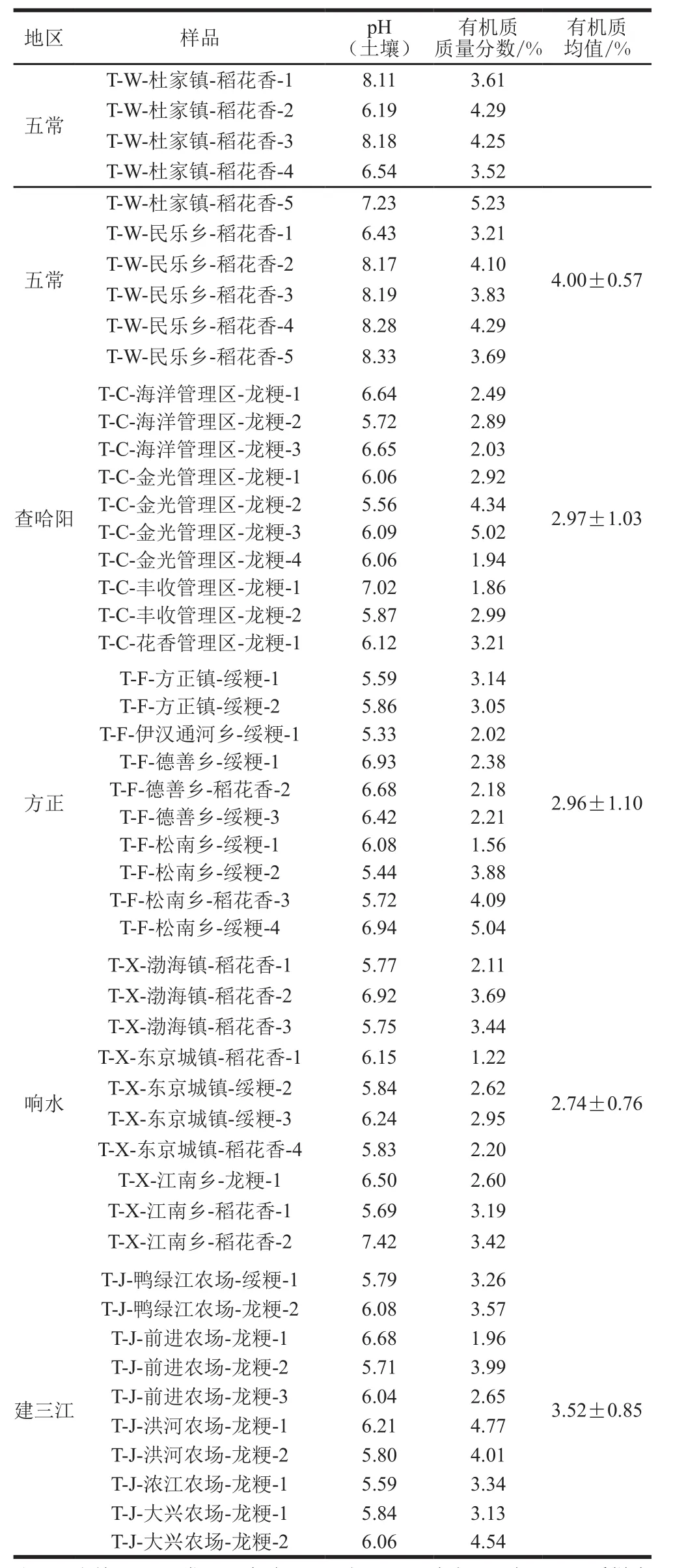
表1 土壤有机质含量和pH值Table 1 Organic matter contents and pH values of soil samples from five rice-producing regions
2.2 糙米中γ-谷维素的含量
所有糙米样品经45 ℃烘干12 h后,水分质量分数变化范围在14%~19%之间,计算样品干基中γ-谷维素含量。糙米中的γ-谷维素含量变化显著,五常地区的糙米均值含量最高(21.48±5.04)mg/100 g,响水地区的含量最低(16.56±6.29)mg/100 g,5 个区域糙米的γ-谷维素含量变化如表2所示。

表2 糙米样品中γ-谷维素含量和阿魏酸酯组成Table 2 γ-Oryzanol contents and ferulic acid ester composition in brown rice samples

注:同一地区同列不同字母表示差异显著(P<0.05);γ-谷维素均值为地区平均值±变化范围值。
糙米中γ-谷维素含量的变化一直受到研究者的关注,欧州地区糙米γ-谷维素含量为23~63 mg/100 g[15],韩国地区糙米的γ-谷维素含量为26.7~61.6 mg/100 g[14]。此外,有色糙米的γ-谷维素含量明显高于普通糙米,Kim等[19]的研究表明有色糙米比普通糙米γ-谷维素含量高27%。研究报道印度地区的糙米γ-谷维素含量为50~72 mg/100 g[20],可能在印度地区有色米种植量多,从而该地区的稻米γ-谷维素相比于其他地区最高,然而还需要研究者进一步对该区域稻米做广泛检测,才能得出更明确解释。但是有色糙米并不是γ-谷维素含量最高的品种,Aladedunye等[21]对比了北美州野生型稻米(Zizania palustris)和常规型稻米,结果得出北美野生型稻米的γ-谷维素含量(135.2 mg/100 g)是常规稻米的2 倍多。已有文献还报道了其他地区糙米中的γ-谷维素含量为24.6~33.0 mg/100 g(巴基斯坦)和46.9~48.2 mg/100 g(日本)[22-24],然而我国的相关数据却难以获得。与其他地区稻米对比,我国黑龙江地区糙米中γ-谷维素含量略低,为10.88~33.53 mg/100 g(表2),与巴基斯坦糙米极其相似,但在已有的文献研究中,发现稻米样本容量太小,进而很难从整体上反映该地区稻米γ-谷维素含量的变化,还需要研究者进一步提供数据。另外在实验中,检测设备和方法的不同也会造成一定差异[25],同时我国有色稻米的种植也受限,其他无色稻米中γ-谷维素含量也不高。在本实验中,发现γ-谷维素含量最大值是最小值的3 倍左右,这与南美稻米米糠中谷维素含量(251~684 mg/100 g)变化范围相似,此外也接近于自然条件下小麦(6.2~12.3 mg/100 g)和玉米(3.1~7.0 mg/100 g)中阿魏酸酯的变化量[26]。如表2所示,糙米中γ-谷维素含量远高于其他谷物的阿魏酸酯含量,如黑小麦(5 mg/100 g)、小麦(12 mg/100 g)、黑麦(6 mg/100 g)、大麦(0.4 mg/100 g)、玉米(7 mg/100 g)[27-29]。
对黑龙江省主要产区稻花香长粒米(五常10 份、方正2 份、响水5 份)与粳稻圆粒米(查哈阳10 份、方正8 份、响水3 份、建三江10 份)样本分析后发现,γ-谷维素含量分别为20.26 mg/100 g和19.25 mg/100 g,如表2所示,可以得出该区域内稻花香长粒米比粳稻圆粒米γ-谷维素含量高。以两区域间同一品种为考察因素,比较查哈阳(10 份)与建三江(9 份)龙粳稻,发现建三江地区糙米中γ-谷维素的平均值稍高于查哈阳地区。同时在比较五常地区(10 份样品)与响水地区(5 份样品)的稻花香系列时,发现五常地区均值(21.48 mg/100 g)明显高于响水地区(20.64 mg/100 g)。如表2所示,5 个地区间的γ-谷维素含量存在明显差异,五常均值为21.48 mg/100 g、建三江均值为21.40 mg/100 g、查哈阳均值为20.53 mg/100 g、方正均值为17.33 mg/100 g、响水均值为16.56 mg/100 g,发现这一结果与表1土壤有机质均值呈正相关。
同一区域相同品种间的差异也很大,发现五常稻花香糙米的γ-谷维素最大值(27.25 mg/100 g)是最小值(12.32 mg/100 g)的2 倍多;建三江龙粳系列糙米中,γ-谷维素最大值为26.06 mg/100 g比最小值16.77 mg/100 g高出近10 mg/100 g,但是由于在同一地区阳光等其他气候条件几乎没有太大差异,产生此现象是由人为环境差异所造成,即田间管理方式(如土壤施肥情况)等,Cho等[30]报道有机土壤栽培的稻米比传统栽培的稻米中γ-谷维素含量高5 mg/100 g;此外还有研究者以中国台湾粳稻Tainung 71为对象,设计4 组土壤有机质的变化对水稻生长情况的影响,当电导率值为8 dS/m时,稻米中谷维素含量最高(19.2 mg/100 g),其值比最低组高出5.8 mg/100 g,但是γ-生育酚含量却降到最低[31];在本研究中也得到相同的结果,土壤中有机质较高的地区,糙米中γ-谷维素含量也相对较高,如表1、2所示。由此可以通过土壤有机质的提高,从而改善稻米中营养物质的积累。
2.3 糙米中阿魏酸酯的组成分析
2.3.1 地区间阿魏酸酯的含量变化
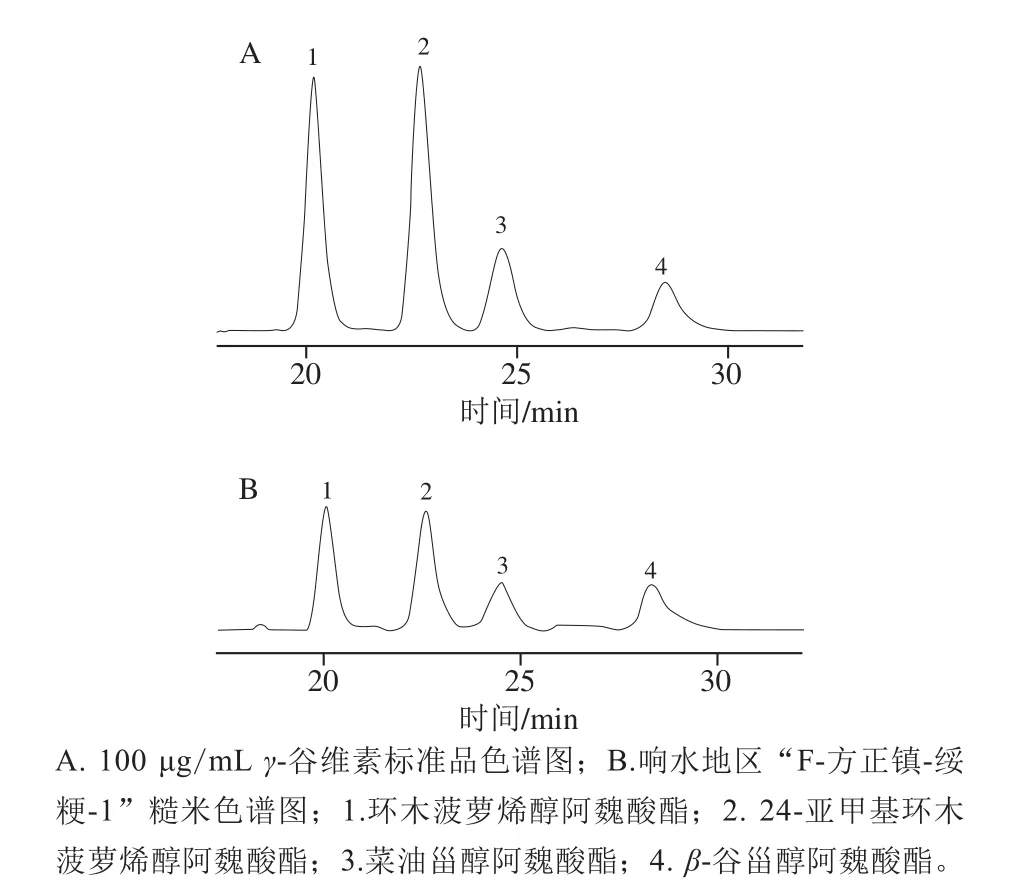
图2 样品液相色谱图Fig.2 Liquid chromatograms of samples
如表2、图2所示,环木菠萝烯醇阿魏酸酯和24-亚甲基环木菠萝烯醇阿魏酸酯含量最多,其次是β-谷甾醇阿魏酸酯和菜油甾醇阿魏酸酯,这些结论与其他研究者报道的结果一致[14-15],但糙米中阿魏酸酯单体均有显著变化,这与小麦和玉米中阿魏酸酯的组成及变化有所不同,玉米和小麦中4,4’-二甲基植物甾醇阿魏酸酯含量极少,甚至无法合成;此外多个小麦和玉米样品间的菜油甾醇阿魏酸酯和β-谷甾醇阿魏酸酯相比变化不明显,组成比例趋于稳定[28]。
同一地区内同一品种的阿魏酸酯组成比有所不同,对建三江地区的9 份龙粳样品分析,得出β-谷甾醇阿魏酸酯占比是菜油甾醇阿魏酸酯占比的1~3 倍,样品间的变化差异较大;同时对查哈阳地区的10 份龙粳样品分析,也得出相同的结果,如表2所示,但是它们的环木菠萝烯醇阿魏酸酯和24-亚甲基环木菠萝烯醇阿魏酸酯的比例很稳定,均约为1∶2,并且4,4’-二甲基类甾醇阿魏酸酯(环木菠萝烯醇阿魏酸酯、24-亚甲基环木菠萝烯醇阿魏酸酯)与4,4’-去甲基类甾醇阿魏酸酯(菜油甾醇阿魏酸酯、β-谷甾醇阿魏酸酯)比为2.6∶1。稻花香长粒米与绥粳圆粒米间阿魏酸酯的组成与前面的分析结果一致,也是环木菠萝烯醇阿魏酸酯和24-亚甲基环木菠萝烯醇阿魏酸酯占比均趋于1∶1.7的比例,并且4,4’-二甲基植物甾醇阿魏酸酯与4,4’-去甲基植物甾醇阿魏酸酯比均约为2.8∶1。
2.3.2 阿魏酸酯含量的相关性

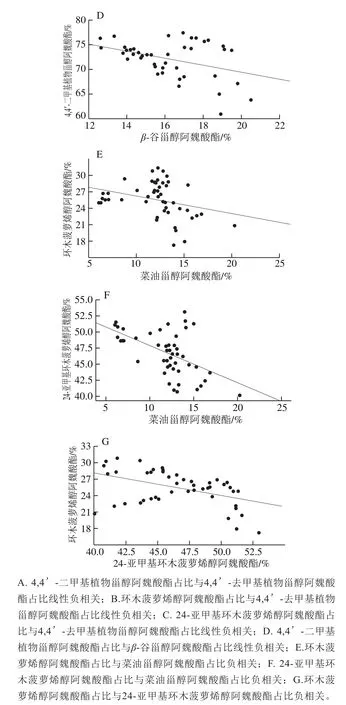
图3 黑龙江水稻糙米中阿魏酸酯的组成(占总γ-谷维素比例)Fig.3 Ferulic acid ester composition of Heilongjiang-grown brown rice(proportions relative to total γ-oryzanol)
在本实验样本中4,4’-二甲基植物甾醇阿魏酸酯的占比和4,4’-去甲基植物甾醇阿魏酸酯的占比相对恒定,分别为76.80%~60.85%和39.15%~23.20%,两类阿魏酸酯间呈负相关,如图3A所示。如图3B、C所示,环木菠萝烯醇阿魏酸酯、24-亚甲基环木菠萝烯醇阿魏酸酯分别和4,4’-去甲基植物甾醇阿魏酸酯呈负相关:r2=-0.455、r2=-0.590。可能是在自然条件下代谢中4,4’-二甲基植物甾醇阿魏酸酯和4,4’-去甲基植物甾醇阿魏酸酯存在相互转化,然而目前还未能探明阿魏酸酯的代谢机理,需进一步的研究。
Miller等[15]研究发现糙米中4,4’-二甲基植物甾醇阿魏酸酯和4,4’-去甲基植物甾醇阿魏酸酯各自的变化不大,认为4,4’-二甲基植物甾醇阿魏酸酯和4,4’-去甲基植物甾醇阿魏酸酯的含量相对恒定。从研究的共同结果看,可以得出4,4’-二甲基植物甾醇阿魏酸酯约是4,4’-去甲基植物甾醇阿魏酸酯的2~3 倍,如表2所示。如图3D所示,β-谷甾醇阿魏酸酯变化12.62%~20.49%相对恒定,是由于β-谷甾醇阿魏酸酯与4,4’-二甲基植物甾醇阿魏酸酯呈负相关(r2=-0.422)。如图3E、F所示,菜籽甾醇阿魏酸酯分别与环木菠萝烯醇阿魏酸酯为r2=-0.316,与24-亚甲基环木菠萝烯醇阿魏酸酯为r2=-0.528的负相关性。
如图3G所示,环木菠萝烯醇阿魏酸酯和24-亚甲基环木菠萝烯醇阿魏酸酯负相关(r2=-0.451),但4,4’-二甲基植物甾醇阿魏酸酯占比恒定,可以得出土壤环境对环木菠萝烯醇阿魏酸酯和24-亚甲基环木菠萝烯醇阿魏酸酯有重要的影响,如表2所示。已经有文献研究得出在蒲公英中发现环木菠萝烯醇脂肪酸酯与24-亚甲基环木菠萝烯醇脂肪酸酯呈负相关;此外在尚未成熟的椰子油中发现,β-谷甾醇含量与Δ5-燕麦甾醇也存在明显的负相关[32],但对糙米样本的分析并没有得出β-谷甾醇阿魏酸酯与菜油甾醇阿魏酸酯存在负相关。
2.4 不同成熟期稻米中γ-谷维素和阿魏酸酯单体的含量变化
2.4.1 不同成熟期稻米中γ-谷维素含量变化
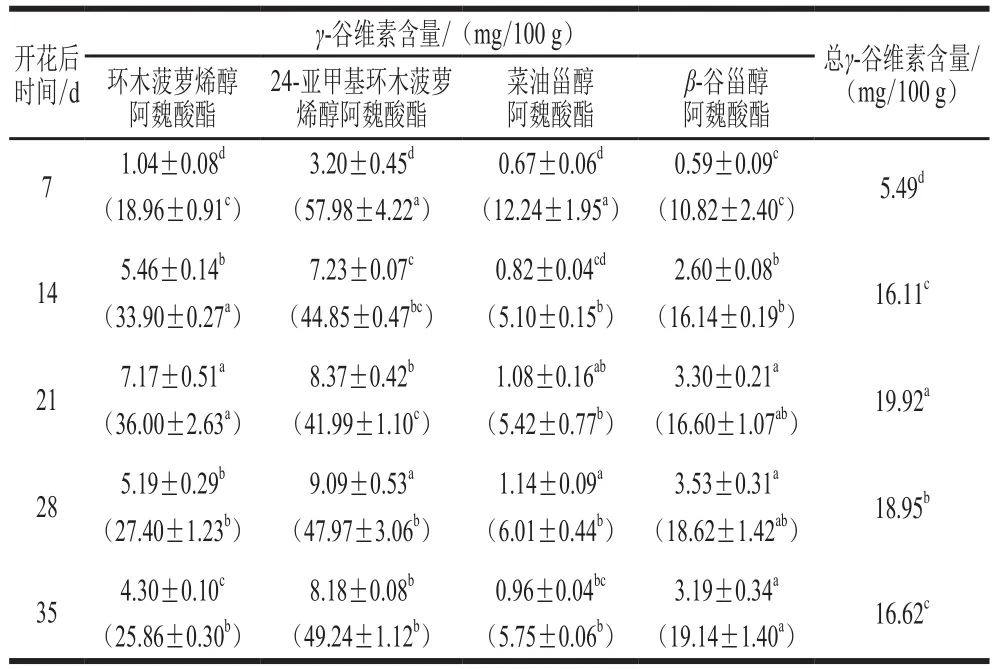
表3 不同成熟期水稻籽粒干物质中γ-谷维素代谢合成量变化Table 3 Changes in γ-oryzanol content at different maturity stages in dry matters
表3为龙粳-31水稻在成熟过程中γ-谷维素含量和阿魏酸酯单体的比例,由开花后7 d的5.49 mg/100 g增加到开花后21 d的19.92 mg/100 g,后又降低到开花后35 d的16.62 mg/100 g。水稻成熟度也对γ-谷维素含量有一定的影响,在早期含量最少,在成熟的过程中逐渐合成,后期γ-谷维素合成量基本停止,含量趋于稳定。水稻籽粒在成熟过程中主要涉及淀粉的积累,占籽粒最终干质量的90%左右,有33 种重要酶参与该过程,其中主要酶为蔗糖合成酶、转化酶、二磷酸腺苷葡萄糖焦磷酸酶、淀粉合成酶、粒结合型淀粉合成酶以及淀粉分支酶[33],水稻籽粒灌浆速率在开花后30 d左右基本降低为零,后期基本维持不变,是由于淀粉合成酶、粒结合型淀粉合成酶以及淀粉分支酶的活性从开花后12 d到开花后36 d开始逐渐降低[34],水稻籽粒在成熟后期主要以脱水为主。由表3可知,开花后21 d籽粒中γ-谷维素含量达到最大值,随后又开始下降,从实验结果和分析可以得出γ-谷维素在开花后21 d前期合成速率大于淀粉合成速率,开花后21 d时淀粉的合成处在旺盛期,γ-谷维素合成的速率低于淀粉合成率,开花后21 d后以淀粉累积为主导,从而在开花后21 d后γ-谷维素含量呈逐渐降低趋势。
2.4.2 不同成熟期稻米中阿魏酸酯单体的含量变化
如表3所示,在水稻籽粒成熟过程中环木菠萝烯醇阿魏酸酯从开花后7 d的1.04 mg/100 g增加到开花后21 d的7.17 mg/100 g,后又减少到开花后35 d的4.30 mg/100 g;24-亚甲基环木菠萝烯醇阿魏酸酯含量最高,开花后7~14 d含量急剧增加,后缓慢增加到开花后28 d的9.09 mg/100 g;菜油甾醇阿魏酸酯含量前后变化不大,由开花后7 d的0.67 mg/100 g缓慢增加到开花后28 d的1.14 mg/100 g;β-谷甾醇阿魏酸酯由开花后7 d的0.59 mg/100 g增加到开花后21 d的3.30 mg/100 g,后趋于稳定。在整个水稻籽粒成熟过程中4,4’-二甲基植物甾醇阿魏酸酯占比由开花后7 d的77%降低到开花后35 d的75%,基本保持不变,其中24-亚甲基环木菠萝烯醇阿魏酸酯占比呈先急剧下降后缓慢增加,环木菠萝烯醇阿魏酸酯占比则相反,呈先急剧增加后缓慢下降的趋势,在开花后21 d是比例变化突变点。4,4’-去甲基植物甾醇阿魏酸酯在整个籽粒成熟过程中的百分比变化不大,其中菜油甾醇阿魏酸酯占比由开花后7 d的12.24%显著降低到开花后14 d的5.10%,后期基本保持不变;β-谷甾醇阿魏酸酯占比从开花后7 d的10.82%显著增加到开花后21 d的16.60%,在开花后14 d是比例变化突变点。图4为植物体甾醇的合成路径,环木菠萝烯甾醇作为前体经过24-亚甲基环木菠萝烯甾醇中间体后分别通过不同的路径合成菜油甾醇和β-谷甾醇。γ-谷维素是由阿魏酸酯与甾醇酯化合成的脂质体化合物[35],在籽粒35 d生长过程中,甾醇代谢前体环木菠萝烯醇甾醇及中间代谢体24-亚甲基环木菠萝烯醇甾醇和阿魏酸结合的酯化物(环木菠萝烯醇阿魏酸酯、24-亚甲基环木菠萝烯醇阿魏酸酯)占比量最高,有文献报道来源于黏稠色杆菌(Chromobacterium viscosum)和念球菌(Candidacylindracea)的脂肪酶对甾醇A环羟基有专门酯化作用[36],除了酶的专一性可能还与甾醇体结构性的差异有关,环木菠萝烯醇甾醇及24-亚甲基环木菠萝烯醇甾醇的甾醇体末端羟基邻位4号碳上存在2 个甲基,导致醇羟基上的氧电负性增强,更容易亲核攻击阿魏酸上的羧基碳,从而使水稻在生长代谢过程中环木菠萝烯醇甾醇及中间代谢体24-亚甲基环木菠萝烯醇甾醇更容易酯化,但是水稻籽粒中甾醇与阿魏酸酯的酯化合成路径还需要经一部的研究。

图4 植物中甾醇的代谢合成路径Fig.4 Biosynthesis pathways of sterols and fatty acids in plants
3 讨 论
基于高效液相色谱检测技术分析了自然条件下黑龙江五大稻米产区糙米样品中γ-谷维素的含量差异,并且得出阿魏酸酯单体含量之间存在一定的相关性。区域γ-谷维素均值的最大极差值为4.92 mg/100 g(五常均值最高,响水均值最低),所有样品中4,4’-二甲基植物甾醇阿魏酸酯(环木菠萝烯醇阿魏酸酯、24-亚甲基环木菠萝烯醇)和4,4’-去甲基植物甾醇阿魏酸酯(菜油甾醇阿魏酸酯、β-谷甾醇阿魏酸酯)占比为2.6∶1,可能是由于环木菠萝烯醇阿魏酸酯和24-亚甲基环木菠萝烯醇阿魏酸酯和甾醇代谢通路也存在如图4相同的相互转化关系,维持比例的恒定。菜籽甾醇阿魏酸酯与24-亚甲基环木菠萝烯醇阿魏酸酯相关性最强。水稻籽粒在开花后28 d后淀粉合成基本停止,后期以脱水为主,理论上谷维素含量应该呈平稳趋势或者呈略微者上升趋势,但是在本实验结果中开花后28 d后水稻籽粒干物质中谷维素含量仍呈下降趋势。实验结果得出土壤有机质含量影响糙米谷维素的含量(表1、2),这一结果与Cho[30]和Tung[31]等的研究结果一致,而在本实验中土壤pH值的变化未发现影响糙米谷维素的含量,同时发现稻米谷维素含量也受不同成熟度的影响。
本实验分析了水稻籽粒不同成熟阶段的阿魏酸酯单体变化,为进一步探究水稻籽粒中阿魏酸酯单体代谢提供参考。得出的数据能为黑龙江省稻米业的精深加工以及掌握该区域内稻米中主要营养物质γ-谷维素含量提供依据。
猜你喜欢
食品安全导刊(2021年21期)2021-08-30
生物化工(2021年3期)2021-07-10
猪业科学(2018年4期)2018-05-19
天然产物研究与开发(2018年2期)2018-04-04
中国粮油学报(2017年5期)2017-07-19
中国粮油学报(2016年5期)2016-01-23
中国粮油学报(2015年5期)2015-02-06
中国药理学通报(2014年2期)2014-05-09
中国粮油学报(2014年8期)2014-02-06
中国粮油学报(2014年7期)2014-02-06
