
青藏高原高寒冻土区生物结皮对浅层土壤水热过程的影响
2020-11-12 07:29孔令阳赵允格杜玉霞
生态学报 2020年18期
明 姣,孔令阳,赵允格,杜玉霞
1 中国科学院西北生态环境资源研究院 冻土工程国家重点实验室, 兰州 730000 2 吉林省水利科学研究院, 长春 130022 3 中国科学院水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100
冻土是一种对温度和外界因素极为敏感的负温地质体[1],其稳定性和土壤的温度场有着直接的联系[1-2]。近年来,青藏高原浅层土壤温度呈升高趋势[3-4],从而导致冻土退化,严重影响多年冻土区地表热量平衡、植被生长、水文循环和工程建设等生态水文过程[5-7]。因此,冻土土体水热变化研究是冻土研究中一直被关注的问题[7-9]。而冻土区浅层土壤水热状态的变化与下垫面条件密切相关[10]。
20世纪70年代以来,随着气候的逐渐干旱化和人口、牲畜数量的迅速增加,青藏高原黄河源区生态环境急剧恶化,至20世纪末,植被退化面积占源区草地总面积1/3[11]。自2005年以来,随着三江源地区“生态保护和建设”工程的具体实施(围栏封育、退牧还草、沙漠化治理、湿地保护等)[11-12],生物结皮开始在该地区逐渐发育,其盖度可高达52%[13-14],成为青藏高原高寒草甸退化过程中地被层的重要组成部分。生物结皮作为地表特殊的结构层,其可通过改变土壤理化属性(颗粒组成、容重、孔隙度、养分含量等)、土壤反照率、土层水分状况,而使土壤热性质发生变化,继而改变土体温度[15-17]。迄今,关于生物结皮对土壤温度的影响,其研究结果在不同区域存在较大差异。早期Belnap(1995)在美国犹他州沙漠地区的研究表明,藻-地衣结皮在冬夏分别增加土表温度14、10 ℃[17]。在极地沙漠地区的研究也表明,在湿热的夏季,深色生物结皮降低土壤表层温度11.8 ℃,而在干冷的冬季,生物结皮则增加地表粗糙度,降低了地表反照率从而使土壤表层温度升高[18-19]。然而,近年来杨永胜等人在中国毛乌素沙地研究则表明,生物结皮的覆盖极显著降低了地表土壤温度[20-21]。同时,Xiao等研究也表明,在湿热的夏季,生物结皮降低土壤表层温度11.8 ℃,而在干冷的冬季,生物结皮则增加土壤表层温度1.2 ℃[22]。因此,生物结皮对土壤温度的影响可能由于生物结皮的组成、发育状况、土壤湿度条件,下伏土壤性质以及气候条件不同而不同[22-27]。
青藏高原地处中国西南边陲,海拔高,气候寒冷,植被类型独特,被誉为“地球第三极”[28]。高寒冻土区土壤温度是反映土壤水分相变(即冻融)过程中热量传递的重要指标,长期浅层(0—30 cm)地温升高会改变深层温度、水分空间分布特征及土壤水热耦合关系,引起冻土退化,进而影响冻土区生态平衡[29-33]。因此,该地区广泛发育的生物结皮很可能成为影响冻土区浅层土壤水热变化的关键影响因素之一。亟须探明生物结皮对青藏高原冻土区生物结皮对冻土浅层土壤水热变化的影响,这也是明确生物结皮在高寒冻土区生态功能的前提。
为此,本文以青藏高原黄河源区玛多县附近的退化草甸中的生物结皮为研究对象,布设野外定点监测装置,观测生物结皮覆盖下浅层冻土土体水温变化过程,以探知高寒冻土区草甸退化过程中,生物结皮对下覆土体水温变化的影响,为评估生物结皮在高寒地区生态功能提供理论基础。
1 材料与方法
1.1 研究区概况
观测场地位于玛多县野马滩,位于38°34′N,98°00′E,海拔4320 m。位于青海省南部(图1),巴颜喀拉山北麓。该地属于平原地区,平均海拔4200 m。植被类型以草地为主。区域气候类型属于青藏高原亚寒带半干旱半湿润型气候[34-35],昼夜温差大,年平均气温-1.6℃;年内降水多集中在5—9月,年内累积降水量407—582 mm;年累积蒸发量为484—584 mm[32]。该地区特有的地理位置和地形、地貌、水文、干燥寒冷的气候条件决定了该区为季节冻土区,并形成镶嵌着不连续多年冻土、零星高温多年冻土和季节性冻土[36-37]。

图1 高寒草甸冻土区生物结皮野外监测场地
1.2 场地布设及野外监测
1.2.1野外调查及采样
于两种(浅色藻结皮和深色藻结皮)不同类型生物结皮斑块中,分别选取人为干扰较少、地形地貌呈现较高一致性、生物结皮发育稳定的5—6封禁地作为研究样地,即每个样地重复5—6次。每个样地中随机设定4个5 m×5 m的调查样方(大样方),每个大样方中随机选择10个25 cm×25 cm的小样方,调查生物结皮物种组成、盖度及样地植被的详细情况。样点的具体情况如表1。
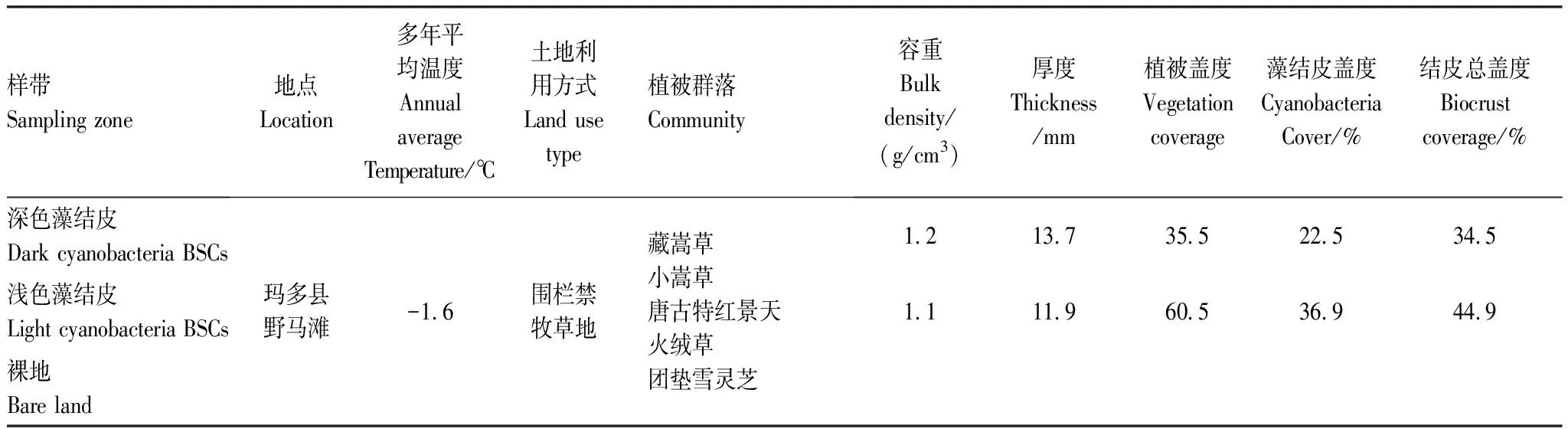
表1 土壤采样点基本情况
1.2.2场地布设及野外监测
2016年10月在研究区根据生物结皮的特征差异布置了3个水温监测场地,分别为深色藻结皮,浅色藻结皮及裸地,每个样点相距近100 m左右。地表状况如图2所示。在各场地5cm、15 cm、30 cm、50 cm深度处布设水温一体温度探头(5TM, Campbell Scientic, Inc., 精度:0.1℃/0.08% WC);采用HC2-S3 温度探头测量气温,其测量范围-40—100℃,精度为±0.1℃;采用RG3-M 自动式雨量筒雨量记录仪测定降水量,其量程0—127 cm/h, 精度±1.0%。所有的探头与CR300数据采集器连接,进行自动采集,所有的要素为每10 min记录一次数据。

图2 高寒草甸退化过程中地表特征
生物结皮层及裸地表面土壤导热系数测定:每种地表覆盖下选择三块试验样地,每个样地以梅花式选取5个点,使用导热系数测定仪 KD2 Pro(Decagon)进行土壤导热系数的测定,每种地表类型土壤导热系数取15次测量平均值。
1.2.3数据处理
本文采用5—50 cm土壤温度与土壤水分含量,其计算方法为:
式中,T10min为数采仪每10 min采集的温度值,θ10min数采仪每10 min采集的土壤未冻水含水量;Th为数采仪每小时平均温度,θh为每小时平均含水量;MST和MWC为日平均温度和土壤含水量;MST1和MWC1为2016年10月—2017年10月日均温及日均含水量,MST2和MWC2为2017年10月—2018年10月日均温及日均含水量MSTm和MWCm为2016年—2018年日均温及日均土壤含水量平均值。借助于Origin 9和SPSS 17.0完成数据处理和统计分析,针对各种类型的生物结皮及裸地土壤导热系数及其理化属性分别进行单因素方差分析,用LSD法进行多重比较,处理间差异均达5%显著水平。选取深色藻结皮为代表,以2016—2018年日均温度的平均值MSTm和日均含水量平均值MWCm,根据土壤温度变化情况,分为3个阶段,T<-4℃,-4℃
2 结果分析
2.1 生物结皮及其覆盖下浅层土体土壤导热系数
土壤的导热系数是反应土体热量传导的能力。通过野外实际测量可知生物结皮层土壤导热系数显著低于裸地(图3),且深色藻结皮的导热系数显著低于浅色藻结皮。同时,深色藻结皮覆盖下,0—30 cm 土层土壤导热系数均显著低于浅色藻结皮和裸地。然而浅色藻结皮0—10 cm土层土壤导热系数亦低于裸地,在20—30 cm 土层,二者导热系数没有显著差异。

图3 生物结皮层土壤导热系数
2.2 生物结皮对高寒草甸浅层土壤水分的影响
高寒地区土壤水分变化过程反映了土壤的干湿状况,是地气系统水循环中的重要组成部分。两种类型生物结皮与裸地日均含水量随时间变化情况如图4所示。由图4中可知,两种类型生物结皮对土壤未冻水含量具有显著的影响,但不同类型生物结皮对土壤不同土层土壤未冻水含量的影响不同。深色藻结皮对土壤含水量的影响主要集中在5—15 cm土层,对土壤30—50 cm土层未冻水含量影响较小:冻结期深色藻结皮覆盖下5—50 cm 土层土壤含水量与裸地相比仅增加了0.4%,而融化期5—15 cm土层土壤含水量较裸地增加1%—5.4%(平均为2.9%),其中八月中上旬(8.1—8.16),深色藻结皮覆盖下5 cm土层土壤含水量与裸地之间没有显著差异(差值<0.5%);浅色藻结皮则对表层土壤含水量影响很小,其对土壤未冻水含量的影响主要集中在15—30 cm土层:在融化期,浅色藻结皮覆盖下15 cm土层土壤含水量较裸地增加了2.3%,而浅色藻结皮在冻结期和融化期均增加了30 cm土层土壤含水量,含水量增量分别达3%—5%和5%—12%。

图4 土壤浅层日均未冻水含量变化(深色藻结皮/裸地;浅色藻结皮/裸地)
2.3 生物结皮对高寒草甸浅层土壤温度的影响
土壤的温度变化体现了土壤能量状况的变化过程,以土壤日平均温度开始持续小于0℃为开始冻结日期,日平均温度开始持续大于0℃为开始消融日期[38]。有无生物结皮覆盖浅层土壤温度年内变化过程可分为两个时期:冻结期(11月—次年3月)和融化期(4月—10月)[39]。两种类型生物结皮及裸地土壤日均温年内动态如图5所示。从图5可知,生物结皮对土壤温度的影响主要集中在融化期,冻结期两种类型生物结皮在不同土层于裸地土壤温度差异均不显著;从表2中可知,在冻结期,两种类型生物结皮对表层土壤温度没有显著的影响,其平均温差均小于0.5℃。而在融化期,两种类型生物结皮均显著降低了土壤温度达0.8—1.9℃。同时,生物结皮对表层土壤温度的影响显著低于其下15—50 cm土层,且不同类型生物结皮,对土壤温度的影响程度不同。表2显示深色藻结皮覆盖下土壤在0—50 cm土层深度处土壤温度较裸地分别平均降低了1.1—2.1℃;而浅色藻结皮则较裸地分别降低了0.8—1.3℃,其降温幅度低于深色藻结皮。

表2 两种类型生物结皮覆盖下不同土层土壤与裸地日均温差(ΔT)

图5 土壤浅层日均温变化(深色藻结皮/裸地;浅色藻结皮/裸地)
2.4 生物结皮覆盖下土壤水热耦合过程
土壤中水分与温度密切相关。本研究以深色藻结皮为例,选取深色藻结皮覆盖下5cm土层土壤含水量及土壤温度数据,进行相关性分析。图6为生物结皮影响下土壤未冻水含量与温度相关关系,由图6A可知,生物结皮覆盖下,冻土浅层土壤具有水热耦合效应。土壤中未冻水含量与温度变化的关系可以分为3个阶段:(1)当MSTdark<-4℃时,土壤中未冻水含量基本保持在4.3%左右(图6B);(2)当-4℃

图6 生物结皮覆盖下土壤温度与水分变化相关关系
3 讨论
3.1 生物结皮对浅层冻土水分含量的影响
冻土的温度、湿度等条件直接影响着冻土的分布、地气相互作用及生态环境的变化[39]。生物结皮作为地表广泛发育的地被层,其具有低容重、高孔隙度、高持水性等特点,因此生物结皮的存在能够增加土壤表层含水量这一研究结论在很多研究中已经被证实[26,40]。生物结皮对土壤水分的影响程度与结皮类型有关,不同类型生物结皮其理化属性(容重、孔隙度及厚度不同及有机质含量等)不同,对土壤水分的影响不同[13, 20-21]。肖波等人的研究也表明藓类生物结皮其结皮层土壤含水量显著高于藻类结皮[27, 41]。本研究结果表明,两种类型生物结皮对土壤未冻水含量及分布的影响在不同冻结融化期内存在显著差异。生物结皮对土壤含水量的影响与其持水性有关。如表3所示,由于深色藻结皮容重较低,孔隙度较大,其田间持水量显著高于浅色藻结皮和裸地,浅色藻结皮持水性高于裸地[13]。故深色藻结皮在冻结期内,对5—50 cm土层土壤未冻水含量均没有显著影响;而在融化期由于其高持水性,显著增加了5—15 cm土层土壤含水量,而对30—50 cm土层土壤含水量没有显著影响。浅色藻结皮则与之相反,在整个冻结融化期内,均显著增加了15—30 cm土层土壤含水量,而对表层土壤未冻水含量没有显著影响。生物结皮对土壤未冻水分布状况的影响则与生物结皮的渗透性有关。从表3可见,深色藻结皮的初始入渗速率为0.20 mm/min,显著低于浅色藻结皮,表明浅色藻结皮具有快速吸收降雨产生水分的能力。由于该区域降水主要发生在5—9月,加之深色藻结皮厚度较大,持水性强,故导致深色藻结皮在融化期显著增加了5—15 cm土壤含水量,而对下层土壤含水量没有显著影响。同时,在8月中上旬,深色藻结皮覆盖下5 cm土层土壤含水量与裸地没有显著差异。由于7月底8月初,太阳辐射最强,气温达到全年最高,土壤蒸发量较大,使得深色结皮层土壤含水量显著降低,验证了土壤温度驱动蒸发作用也是影响土壤含水量的重要因子之一[42-43]。但浅色藻结皮厚度较薄,渗透速率较深色藻结皮高(表3),因此其能够快速吸收降雨产生的径流,水分向下层迁移,使得30 cm土层土壤含水量显著增加,吸收的水分贮存在30 cm土层。因此,其土壤未冻水含量显著性高于裸地。

表3 生物结皮基本属性
3.2 生物结皮对浅层冻土温度的影响
生物结皮对土壤温度的影响主要有两方面:一方面,生物结皮的发育改变地表颜色,从而改变地表反照率,增加或减少进入土层、使土壤温度升高的热量;另外,生物结皮作为地表的特殊结构层,其具有自身的结构及特殊的理化属性,热性质也随之发生改变,故影响土壤温度变化[16,20-24]。早期有研究表明覆被在地表深色的生物结皮,可通过吸收更多太阳辐射而使土壤温度升高[17-18]。本研究区主要以深色及浅色藻结皮为主,其覆盖度可达35%—45%。而研究结果却表明,该区生物结皮的存在显著降低了冻土浅层土壤温度。由此可见,生物结皮由于改变地表颜色从而导致地表反照率的改变并不是冻土区导致土壤温度变化的主要原因。冻土区生物结皮对土壤温度的影响与土壤的冻结融化期相关。在冻结期,生物结皮对土壤浅层温度没有显著的影响;在融化期,生物结皮均显著降低了土壤温度。不同气候类型区及生态环境条件下,生物结皮的发育状况及其理化属性不同,因此其对土壤温度的影响也不同[18,21-22]。肖波等人在黄土高原的研究结果表明,在湿热的夏季,生物结皮降低土壤表层温度11.8℃[22]。杨永胜等在沙区的研究结果也表明生物结皮的覆盖极显著降低了地表的土壤温度[21],这些研究结果与我们的研究结果有相似之处。由于冻结期,生物结皮及其下层土壤均处于冻结状态,生物结皮层的含冰量与裸地的差异并不足以引起土壤热性质的变化[44]。加之,生物结皮由于其较低的导热系数(图3),地气之间热传递较慢,相当于土体表面保温层,在冻结期对冻土具有一定的保温作用,但其对土壤温度的影响并不显著。而在融化期,由于生物结皮层能够固持较多的水分,结皮层热容量随着土壤含水量的增加而增大,生物结皮层存储的热量增加,其向下传递的热量减少[44]。与此同时,在融化期,生物结皮的多孔结构中孔隙被水分填充较多,能够在融化期(暖季)阻止空气中热量向土壤中扩散,能够降低下层土壤温度。再者,生物结皮能够显著提高土壤中碳氮含量,形成一个富含有机质层。有机质作为不良导体,在融化期阻止土体中冷量的流出,也延缓了下覆土体的升温[21,45],亦对土壤具有降温的效应。
生物结皮对土壤温度的影响与生物结皮的类型有关,深色藻结皮覆盖下土壤在5—50 cm土层深度处土壤温度较裸地分别平均降低了1.1—2.1 ℃;浅色藻结皮则较裸地分别降低了0.8—1.8℃,其降温幅度较深色藻结皮低。由于浅色藻结皮的土壤有机质含量显著低于深色藻结皮,同时其田间持水量也显著低于深色藻结皮,故其热容量在融化期较深色藻结皮低,对土壤温度的影响也较小,亦表明生物结皮可通过增加土壤有机质而产生隔热效应及在融化期增加土壤含水量从增加自身储热量而对下层土体温度产生影响[22-25]。
3.3 水温相关关系
水热因子在土壤中相互影响、相互联系、相互制约。通过分析生物结皮层土壤水分和温度的相关关系发现,生物结皮覆盖下土壤含水量与土壤温度密切相关,表明土壤中水热运动的相互耦合作用[20,46-47]。本研究结果表明当MSTdark<-4℃时,生物结皮覆盖下,土壤中未冻水含量并未随着土壤温度的变化,保持在4.3%左右。表明生物结皮在冻结期对土壤温度影响不显著。验证了生物结皮对土壤温度的影响与土壤中水分含量密切相关[20-21]。当-4℃

图7 不同温度下土壤未冻水含量剖面变化
浅色藻结皮和裸地0—15 cm土层土壤含水量在15℃时略低于6℃及12℃时土壤含水量,但差异不显著。表明浅色藻结皮对土壤温度的影响较深色藻结皮低。然而,随着土壤温度的增加,浅色藻结皮30 cm土层处土壤含水量显著增加,图8为不同温度下土壤未冻水储量剖面变化。从图7及图8可知,随着温度升高,30 cm土层处土壤未冻水含量及储量均显著增加。因此浅色藻结皮在融化期降低了下层15—50 cm土层土壤的温度。长期浅层土体的温度降低对冻土的保护具有极为重要的作用,同时,由图8可知,由于生物结皮的存在,增加了下层0—50 cm土层土体未冻水的储量,对高寒冻土区植被恢复及生态环境具有重要意义[1,50]。

图8 不同温度下土壤未冻水储量剖面变化
4 结论
(1)生物结皮对冻土浅层土壤水热过程具有显著的影响,而其影响主要发生在土壤融化期内,在冻结期生物结皮对土水热过程没有显著的影响;同时,生物结皮对土壤水热过程的影响不仅与土壤的冻结融化期有关,还与生物结皮的类型密切相关,不同类型生物结皮对土壤的未冻水含量和温度的影响不同:
在融化期内,生物结皮增加了冻土浅层土壤水分,其中深色藻结皮增加了0—15 cm土层土壤含水量,而浅色藻结皮却显著增加了30 cm土层土壤含水量;同时,生物结皮降低冻土浅层(15—50 cm)土壤温度,其中深色藻结皮的降温效应显著高于浅色藻结皮。
(2)冻土地区生物结皮覆盖下浅层土壤具有水热耦合效应:根据浅层土壤水热运移特征可将深色藻结皮覆盖下冻土浅层水热过程分为两个阶段:
土壤未冻水含量<10% 时(土壤温度<4℃),土壤温度影响土壤含水量,其中T<-4℃时,土壤处于完全冻结状态,生物结皮层覆盖下,土壤未冻水含量保持在4.3%左右;-4℃
猜你喜欢
新疆大学学报(自然科学版)(中英文)(2022年3期)2022-06-04
中老年保健(2021年3期)2021-12-03
土壤(2021年5期)2021-11-17
阅读与作文(小学高年级版)(2020年6期)2020-07-04
小哥白尼(趣味科学)(2020年7期)2020-05-22
水土保持通报(2020年6期)2020-03-10
小哥白尼(趣味科学)(2018年9期)2018-12-18
新疆大学学报(自然科学版)(中英文)(2018年4期)2018-02-10
延河(2017年7期)2017-07-19
儿童故事画报·发现号趣味百科(2015年5期)2015-07-22
