
疏勒河源高寒草甸土壤微生物生物量碳氮变化特征
2020-11-12 07:29吴明辉魏培洁贾映兰陈生云
生态学报 2020年18期
刘 放,吴明辉,魏培洁,贾映兰,陈生云
1 中国科学院西北生态环境资源研究院 冰冻圈科学国家重点实验室 疏勒河源冰冻圈与生态环境综合监测研究站, 兰州 730000 2 青海师范大学 高原科学与可持续发展研究院 地理科学学院, 西宁 810008 3 中国科学院大学, 北京 100049
土壤微生物生物量是指除植物根系和体积大于5×103μm3土壤动物以外的土壤中所有活有机体的量[1]。常用土壤微生物生物量和活性表征土壤微生物数量及其功能[2],土壤微生物生物量主要反映了土壤微生物的重量或质量。虽然土壤微生物生物量碳氮仅占有机质中的很小比例,但却是最为活跃的部分[3],影响着土壤有机质的转化且能敏感的指示土壤理化性状的变化,可准确及时地反映土壤质量[4]。此外,土壤微生物生物量与土壤有机质的比率可作为指示土壤碳动态的数量指标[5]。
气候、土壤和植被因子会对土壤微生物生物量产生显著影响,不同区域生态系统中气候、土壤和植被不同,土壤微生物生物量也有较大差异。由于植物的季节性生长及有机质季节性的回归土壤(植物和动物的残留物是形成土壤有机质的主要生物输入物,秋冬季节大量残留复合物进入土壤,被微生物分解形成土壤有机质),使土壤微生物生物量碳氮也具有季节动态[6],其季节变化与环境因子以及植物生理特征密切相关[7]。另外,水热条件的季节变化会影响土壤微生物生物量碳氮的周转、植物对养分的利用以及养分的有效性[8]。早期土壤微生物生物量碳氮的研究主要侧重在森林、草地、农田等土壤肥力方面,随后逐渐深入到其对自然因素和人类活动响应及其与生态系统功能关系等方面。如:土壤微生物生物量碳氮和有机质动态的关系[9]、土地利用[10]对土壤微生物生物量碳氮的影响、不同生态系统中土壤微生物生物量碳氮功能[11]等。国内对土壤微生物生物量碳氮的研究主要分布在红壤侵蚀区[12]、内蒙古草原[13]、黄土丘陵沟壑区[14]、长白山[15]等地,但对青藏高原多年冻土区高寒草地土壤碳氮变化特征方面的研究还相对较少。为准确认识不同气候区不同生态系统中碳、氮平衡和循环机制,还需对不同生态系统中土壤微生物生物量碳氮进行深入研究。
青藏高原位于高海拔地区,约占中国国土面积的25%左右[16],被称为“世界第三极”,是气候变化的敏感区和脆弱区。疏勒河发源于青藏高原东北缘祁连山西段,是我国河西走廊三大内陆河之一。该流域年均气温低,降水量少,蒸发量大,生态环境脆弱。高寒沼泽草甸、高寒草甸、高寒草原是该流域最为典型的生态系统类型,土壤微生物在这些生态系统土壤碳氮循环过程中发挥重要作用。从土壤微生物生物量角度分析疏勒河源区高寒草甸土壤肥力及土壤生态健康状况对该区域生态功能保护和建设具有重要意义。近年来,有关青藏高原高寒草地土壤微生物生物量的研究虽逐渐增多[17-19],但对多年冻土区高寒草甸的相关报道依然缺乏,研究该区域土壤微生物生物量碳氮的季节动态及其影响因素对理解和预测全球气候变化对多年冻土区高寒草甸生态系统的影响具有重要意义。因此,本研究以青藏高原东北缘疏勒河源多年冻土区高寒草甸为研究对象,分析土壤微生物生物量碳氮的季节与土层变化特征及其控制因子,以期为了解疏勒河源区高寒草甸土壤微生物特征提供依据,为系统认识区域土壤碳氮循环规律提供参考。
1 材料与方法
1.1 研究区概况
试验样地位于青藏高原东北缘疏勒河源多年冻土区高寒草甸综合观测场(98°16′14″E,38°21′17″N,海拔约4014 m),2010年8月布设该样地,面积为10000 m2。气象资料显示,年均气温为-4.8℃,降水量为417 mm(图1),空气相对湿度为48.1%,总辐射为204.5 W/m2,干冷多风,属于大陆性干旱荒漠气候[20]。多年冻土类型按其稳定性划分属于亚稳定型多年冻土,土壤类型为简育干润均腐土。植被类型为高寒草甸,覆盖度约40%,代表性植物有波伐早熟禾(Poapoophagorum)、高山嵩草(Kobresiapygmaea)、矮生嵩草(K.humilis)、柔软紫菀(Asterflaccidus)、紫花针茅(Stipapurpurea)、沙生风毛菊(Saussureaarenaria)、西伯利亚蓼(Polygonumsibiricum)和青藏苔草(Carexmoorcroftii)等。
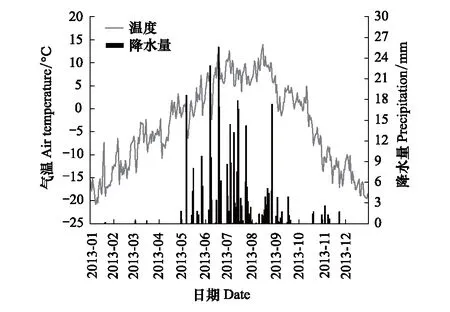
图1 气温和降水量日均变化(2013年样地实测)
1.2 环境因子监测和土壤样品采集
采用HMP155A型温湿度传感器(Vaisala,芬兰)测定气温和相对湿度,Hydra-Probe II型土壤传感器(Stevens,美国)测定土壤温度、含水量和盐分,上述传感器连接至CR1000型数据采集器(Campbell,美国),每10 min记录一次数据。采集土壤样品的时间为2013年4月底和5月初、6月底和8月初、9月底和10月初和12月底,分别表征为春季、夏季、秋季和冬季。在样地内随机选取3个样方(50 cm×50 cm),将各样方内的植物用剪刀齐地剪除,然后用内半径为2.75 cm的土钻由地表到深层分为5层分别取样(0—10、10—20、20—30、30—40、40—50 cm)[20],每层取2份土样。剔除土样中植物根系和石块后,分别用自封袋装好,一份放入4℃冰箱中冷藏,用于测定土壤微生物生物量碳(Soil microbial biomass carbon,SMBC)、土壤微生物生物量氮(Soil microbial biomass nitrogen,SMBN)、pH值和氧化还原电位;另一份自然风干,充分研磨后过筛,用于测定土壤全氮和有机碳含量。
1.3 分析方法
土壤有机碳和全氮含量分别用重铬酸钾氧化-外加热法(GB7857-87)和半微量开氏法(GB7173-87)测定。植物地下生物量测定是将地下生物量样品装入土筛,用水洗掉土壤部分,并去除石块和其他杂物,然后同地上生物量样品一起放入烘箱烘干至恒重,最后称重(精度为0.01 g)测得生物量数据。SMBC和SMBN采用氯仿熏蒸浸提法测定[21]。
1.4 数据处理
数据处理和统计采用Excel 2010和SPSS 24.0软件分析,Origin 8.5作图。采用单因素方差分析(One-way ANOVA)及最小显著性差异法(Least-Significant Difference,LSD)检验SMBC、SMBN季节和土层变化的差异,用双因素方差分析(Two-way ANOVA)检验季节和土层变化及其交互作用对SMBC、SMBN的影响。用Pearson相关和逐步回归分析土壤环境因子与SMBC、SMBN的相互关系及其影响因素。
2 结果分析
2.1 SMBC和SMBN的变化特征
如图2所示,不同土层SMBC均具有明显的季节变化规律,即春季开始逐渐升高,夏季达到最大值,秋季逐渐降低,冬季值最小。0—10、20—30 cm和40—50 cm层不同季节间均无显著差异,而10—20 cm层SMBC夏季与冬季有显著差异,30—40 cm层SMBC夏季与秋、冬季均有显著差异。季节变幅方面,0—10 cm为165.11—257.52 mg/kg、10—20 cm为83.48—183.93 mg/kg、20—30 cm为33.5—96.06 mg/kg、30—40 cm为32.14—67.04 mg/kg、40—50 cm为25.08—42.25 mg/kg。可见,0—10 cm层SMBC季节变幅大于其他各层。
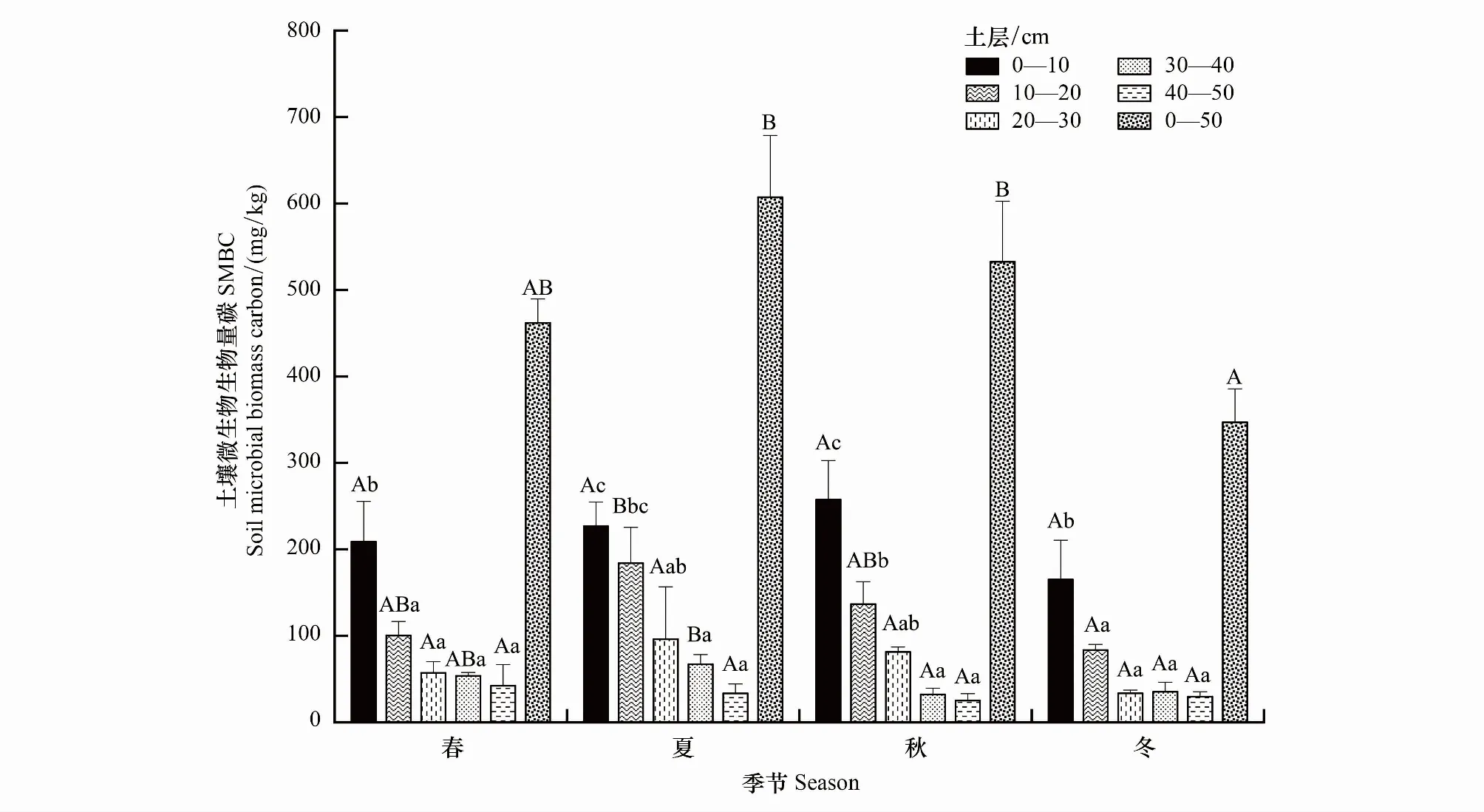
图2 不同土层土壤微生物生物量碳的季节差异
不同土层SMBN也具有明显的季节变化规律(图3)。0—10、20—30、40—50 cm层SMBN均表现春季开始逐渐降低、夏季值最小、秋季逐渐升高、冬季达到最大值,但其不同季节间均无显著差异,10—20、30—40 cm层SMBN最低值分别出现在春季和秋季,而最大值均在冬季,10—20 cm层春季和冬季SMBN有显著差异而30—40 cm层SMBN不同季节间无显著差异。季节变幅方面,0—10 cm为8.72—16.94 mg/kg、10—20 cm为5.49—7.82 mg/kg、20—30 cm为5—6.25 mg/kg、30—40 cm为2.51—5.1 mg/kg、40—50 cm为1.96—3.83 mg/kg,即0—10 cm层SMBN季节变幅大于其他各层。
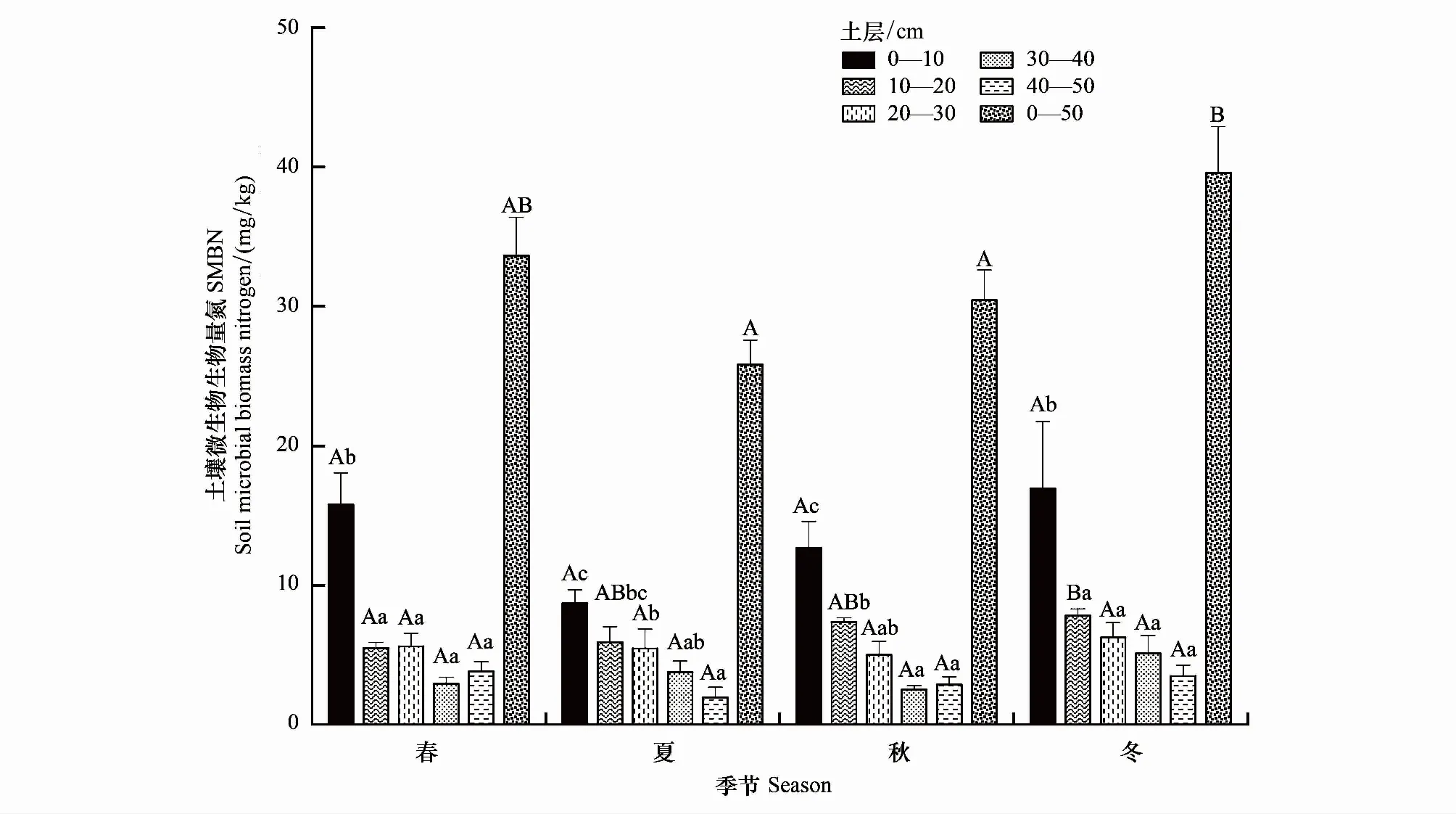
图3 不同土层土壤微生物生物量氮的季节差异
同一季节内,SMBC、SMBN在0—50 cm各土层上均呈现降低趋势,其中0—10 cm层显著高于20—30、30—40、40—50 cm三层。
2.2 SMBC/SMBN的变化特征
如图4所示,不同土层土壤微生物生物量碳氮比(SMBC/SMBN)具有明显的季节变化规律。0—10、10—20、20—30 cm层SMBC/SMBN均春季开始逐渐升高、夏季达到最大值、秋季逐渐降低、冬季值最小,0—10 cm层夏、秋季和冬、春季SMBC/SMBN有显著差异而10—20、20—30 cm层SMBC/SMBN不同季节间均无显著差异。30—40 cm层春、夏季SMBC/SMBN显著高于冬季,40—50 cm层SMBC/SMBN夏季显著高于春、秋和冬季。季节变幅方面,0—10 cm为9.99—26.47、10—20 cm为10.66—36.71、20—30 cm为5.82—17.54、30—40 cm为6.77—19.36、40—50 cm为8.33—17.62,即0—10 cm层SMBC/SMBN季节变幅大于其他各层,且0—50 cm层SMBC/SMBN季节变幅为8.77—23.59。
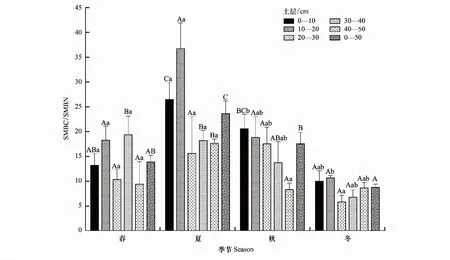
图4 不同土层土壤微生物生物量碳氮比的季节差异
SMBC/SMBN在0—50 cm各土层的季节变化特征各异。其土层变化春季呈“双峰”型变化模式,夏季呈“N”型变化模式,且春和夏季各层间无显著差异。秋季随着土层加深,SMBC/SMBN呈下降趋势,冬季呈“N”型变化模式,且秋、冬两季各层有显著差异。
2.3 SMBC/SOC、SMBN/TN的变化特征
土壤微生物生物量碳占有机碳比例(SMBC/SOC)的各土层季节变化趋势与SMBC变化趋势相似(图5)。0—10 cm的季节变幅为0.97%—1.36%、10—20 cm为0.71%—1.34%、20—30 cm为0.29%—0.99%、30—40 cm为0.26%—1.27%、40—50 cm为0.38%—0.89%。0—50 cm层SMBC/SOC夏、秋季显著高于冬、春季,且其季节变幅为0.58%—1.18%。同一季节内随土层加深,SMBC/SOC大致呈下降趋势。除夏季土层变化无显著差异外,SMBC/SOC均表现0—10 cm层显著高于40—50 cm层。

图5 不同土层土壤微生物生物量碳占有机碳和生物量氮占全氮的比例季节差异
土壤微生物生物量氮占全氮比例(SMBN/TN)的各土层季节变化趋势与SMBN变化趋势相似。季节变幅方面,0—10 cm为0.46%—0.97%、10—20 cm为0.37%—0.71%、20—30 cm为0.4%—0.66%、30—40 cm为0.29%—0.64%、40—50 cm为0.27%—0.74%。0—50 cm层SMBN/TN秋、冬季显著高于夏季,且其季节变幅为0.39%—0.72%,而SMBC/SOC季节变幅大于SMBN/TN。同一季节内随土层加深,SMBN/TN各土层变化特征与SMBC/SOC的变化特征一致。
2.4 影响因素
由表1可知,不同季节的SMBC、SMBN有显著差异(P<0.05),SMBC/SMBN有极显著差异(P<0.001);不同土层的SMBC、SMBN有极显著差异(P<0.001),SMBC/SMBN有显著差异(P<0.05)。季节和土层变化不存在显著的交互效应,对SMBC、SMBN和SMBC/SMBN的差异没有贡献。
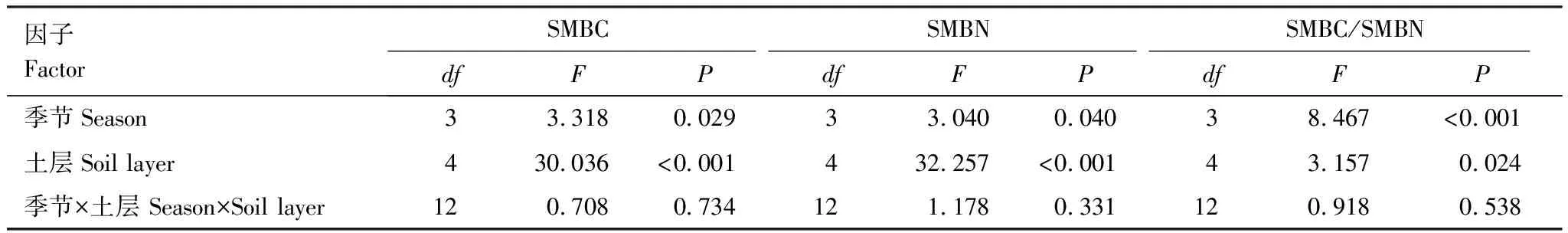
表1 土壤微生物生物量碳、氮及其比值的方差分析结果
季节变化而言,SMBC与土壤含水率(SWC)、土壤盐分(SS)、pH值显著正相关(P<0.05),与土壤温度(ST)有极显著正相关关系(P<0.01),但与地下生物量(BGB)、有机碳(SOC)、全氮(TN)、氧化还原电位(Eh)无显著相关关系。SMBN与ST、SS、pH值显著负相关(P<0.05),与其他土壤环境因子无显著相关关系。SMBC/SMBN与ST、SWC、pH值显著正相关(P<0.05),而与其他土壤环境因子无显著相关关系。逐步回归分析表明,SMBC、SMBN、SMBC/SMBN季节变化的驱动因子分别为BGB、BGB和ST、ST(表2)。

表2 季节变化上的土壤微生物生物量碳、氮及其比值与土壤环境因子相关性及回归分析
随土层变化,SMBC与SOC、TN、ST有极显著正相关关系(P<0.01),与BGB显著正相关(P<0.05),与pH值显著负相关(P<0.05),而与SWC、SS、Eh无显著相关关系。SMBN与BGB、SOC、TN、ST有极显著正相关关系(P<0.01),而与其他土壤环境因子无显著相关关系。SMBC/SMBN与土壤环境因子之间无显著相关关系。逐步回归分析表明,SMBC、SMBN、SMBC/SMBN土层变化的驱动因子分别为ST、BGB、ST(表3)。
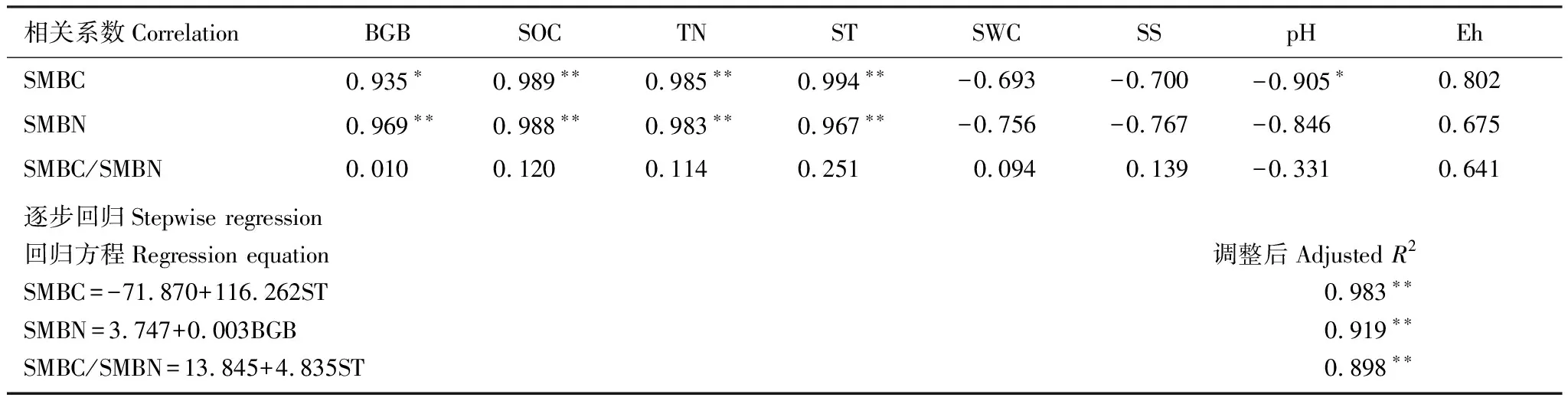
表3 随土层变化的土壤微生物生物量碳、氮及其比值与土壤环境因子相关性及回归分析
3 讨论
3.1 SMBC、SMBN的变化特征
植被生态系统土壤微生物生物量受复杂的生物地球化学过程影响,导致不同季节、不同土层SMBC、SMBN存在一定差异[22]。众多研究结果也表明,SMBC、SMBN受季节变化、土层变化影响显著[23-25],然而其结果并没有统一的规律。疏勒河源多年冻土区高寒草甸SMBC与SMBN的季节变化有所差异,总体上SMBC春季开始逐渐升高,夏季达到最大值,秋季逐渐降低,冬季值最小,这与以往研究结果一致[26-27]。夏季适宜的土壤温度、湿度为土壤微生物的繁殖提供了良好的代谢环境,从而增加了SMBC,冬季较低的土壤温度、湿度抑制了土壤微生物的生长与繁殖,削弱了其代谢功能,从而降低了SMBC[4]。而SMBN总体上春季开始逐渐降低、夏季值最小、秋季逐渐升高、冬季达到最大值,这与Saratchandra等[28]结果一致,峰值出现在冬季可能与秋季大量凋落物的输入以及碳水化合物由地表不断向地下转移有关,为冬季土壤微生物提供了较丰富的代谢底物,尽管该区冬季气温很低,但由于积雪覆盖对土壤的保温作用,使土壤微生物仍具活性继而生长和繁殖;Wardle[3]发现,在生长旺季植物和土壤微生物对养分的利用存在激烈的竞争关系,可导致生长旺季微生物量发生变化,夏季SMBN最低,可能是植物进入生长旺季,需从土壤中摄取大量营养物质,从而限制了土壤微生物对养分的可利用性。但研究区SMBN季节变化特征与Devi和Yadava[4]、Singh等[8]的研究结果不同,可能是由于不同研究区域气候、土壤、植被条件的不同,导致研究结果有差异。
表层SMBC、SMBN季节变幅较深层土壤更大,可能是表层土壤物理环境变化较剧烈、底物输入波动较大、植物根系与土壤微生物对养分的竞争较激烈等原因[29],从而使表层土壤微生物生物量季节变幅更大。SMBC、SMBN均随土层深度的增加而显著降低,与Wen等[30]研究结果一致,且0—10 cm层SMBC、SMBN显著高于20—30、30—40、40—50 cm,这反映了土壤微生物的垂直分布格局。形成这种垂直分布格局的原因可能是表层土壤水热、通气状况良好、有机物质丰富且有较多的碳源输入(凋落物分解、根系分泌物形成),为微生物提供充足的能源,而中层和深层土壤微生物生物量主要受营养元素在土壤中长期积累及该土层根系周转的影响[31],因此,SMBC、SMBN均随土层深度的增加而降低。
3.2 SMBC/SMBN的变化特征
有研究表明,细菌的碳氮比为3—5之间,而真菌的碳氮比为4—15之间[32],且相较于细菌,真菌往往能适应较差的土壤环境。因而,SMBC/SMBN在一定程度上不仅可以反映土壤中真菌和细菌的比例,同时也可以说明土壤环境状况[33]。同样,季节变化和土层变化对SMBC/SMBN均有显著影响。研究区SMBC/SMBN季节变化介于5.82—36.71,与杨成德等[34]研究结果(3.19—20.55)有所差异,说明疏勒河源多年冻土区高寒草甸土壤微生物群落中,真菌占优势。总体上SMBC/SMBN春季开始逐渐升高、夏季达到最大值、秋季逐渐降低、冬季值最小,可能由于夏季土壤含水率较高,同时有地表凋落物的输入,促进了土壤中真菌的大量繁殖,此时高寒草甸固碳能力达到最高[35];而冬季SMBC/SMBN偏低,这可能是由于冬季植物根系分泌了能促进土壤中细菌繁殖的物质,导致SMBC/SMBN偏低[36]。0—10 cm层SMBC/SMBN的季节变幅较40—50 cm层更大,即40—50 cm层的固碳能力较0—10 cm层更稳定,这与Bailey等[35]研究结果一致。不同季节SMBC/SMBN土层变化特征各异,春季和夏季,10—20 cm层SMBC/SMBN均高于0—10 cm层,可能由于春季积雪融水和夏季降雨增加,淋溶作用使营养物质渗入地表下层,下层土壤植物根系发达,从而使大量菌根真菌繁殖,导致SMBC/SMBN升高[37]。而SMBC/SMBN在秋季随土层深度的增加而降低,在冬季则呈“N”型变化趋势,这与魏卫东和刘育红[38]研究结果不同,可能是由于不同研究区域气候、土壤、植被条件的不同,导致研究结果有差异。SMBC/SMBN的季节和土层差异说明了青藏高原环境的特殊性,其土壤中可能存在独特的微生物,即该区域有很多需进一步研究的特殊微生物群落。
3.3 SMBC/SOC、SMBN/TN的变化特征
土壤微生物生物量与土壤养分的比值可以反映土壤养分向土壤微生物生物量的周转效率、土壤养分损失,且其在表征土壤变化过程时比单独使用土壤微生物生物量或土壤养分更有效[39]。研究区SMBC对SOC贡献率在夏季最高,冬季最低;而SMBN对TN贡献率在秋季最高,夏季最低,这与杨成德等[34]研究结果相同。SMBC对SOC贡献率介于0.58%—1.18%,而SMBN对TN贡献率介于0.39%—0.72%,与杨成德等[34]在东祁连山高寒草地的SMBC/SOC(0.16%—0.84%)相比略高,与SMBN/TN(0.48%—1.93%)相比略低;低于Liao和Xie[40]研究植被覆盖下的SMBC/SOC(2%—7%),且低于Devi和Yadava[4]研究混合森林系统的SMBN/TN(2%—7.8%),造成这种差异的原因可能是研究区气候恶劣,海拔较高,微生物活性较低。同一季节随土层加深,SMBC/SOC、SMBN/TN均大致呈下降趋势,即0—10 cm层SMBC对SOC贡献率、SMBN对TN贡献率均高于40—50 cm层,表明40—50 cm层对SMBC、SMBN的累积能力弱于0—10 cm层,这与吴建国和艾丽[2]研究结果一致。
3.4 影响因素分析
影响SMBC、SMBN的因素复杂多样。季节变化而言,SMBC与ST呈极显著正相关关系,即研究区ST是影响SMBC季节变化的主要因素之一,且SMBC与SWC、SS、pH呈显著正相关关系,说明SWC、SS、pH可能也是影响SMBC季节变化的重要因素,这与前人的研究结果一致[41-42]。但SMBN与ST、SS、pH均呈显著负相关关系,这与Arnold等[43]研究结果有差异,表明土壤微生物生物量受ST、SS、pH的影响具有一定复杂性,不同研究区得到的研究结果可能不同。SMBC/SMBN与ST、SWC、pH呈显著正相关关系,其季节变化趋势受ST、SWC、pH季节变化的综合影响。
随土层变化,SMBC、SMBN均与SOC、TN、ST呈极显著正相关关系,这与前人研究结果一致[44-45],由此可见,除ST外,不同土层土壤养分状况的差异可能也是影响SMBC、SMBN变化的重要因素,有机质含量越高可为微生物新陈代谢提供的物质和能量来源越充足,SOC、TN随土壤深度增加而降低,这与SMBC、SMBN的土层变化趋势一致。SMBC与pH呈显著负相关关系,这与柴雪思等[46]研究结果一致,但与Ma等[47]在三角洲湿地的研究结果不完全相同,表明弱酸性环境可能更有利于微生物的繁殖,但pH值对SMBC的影响因研究区不同可能有差异。BGB与SMBC呈显著正相关关系,与SMBN呈极显著正相关关系,说明BGB也是影响SMBC、SMBN在土层变化的重要因素,随土层深度增加,BGB降低,SMBC、SMBN也呈下降的趋势。SMBC/SMBN与土壤环境因子之间未发现显著相关关系,这可能与研究区特殊的土壤环境条件有关。
4 结论
疏勒河源多年冻土区高寒草甸SMBC、SMBN季节变化范围分别为346.9—607.25 mg/kg、25.84—39.59 mg/kg;SMBC对SOC贡献率介于0.58%—1.18%,SMBN对TN贡献率范围为0.39%—0.72%,均处较低水平;SMBC/SMBN变化范围为8.77—23.59,处于较高水平。SMBC、SMBN均随土层深度的增加而显著降低且0—10 cm层SMBC、SMBN、SMBC/SMBN季节变幅大于其他各层,即40—50 cm层的固碳能力较0—10 cm层更稳定;随土层加深,SMBC/SOC、SMBN/TN均大致呈下降趋势,表明40—50 cm层对SMBC、SMBN的累积能力弱于0—10 cm。SMBC、SMBN的季节和土层变化受ST、SOC、TN、BGB、pH、SWC、SS的影响显著,其中ST和BGB是SMBC、SMBN变化的主要影响因素。
猜你喜欢
农业装备与车辆工程(2021年10期)2021-11-03
今日农业(2020年19期)2020-12-14
水电与新能源(2019年10期)2019-11-06
北京航空航天大学学报(2019年8期)2019-09-10
资源节约与环保(2018年1期)2018-02-08
资源节约与环保(2018年1期)2018-02-08
中国资源综合利用(2017年3期)2018-01-22
中国科技信息(2015年2期)2015-11-16
中国质量与标准导报(2015年2期)2015-02-28
振动与冲击(2014年9期)2014-09-05
