
龙须菜对翘嘴红鲌生长、消化和血液生化指标的影响
2020-11-15 11:32宣雄智李文嘉朱文婷苏杰南
中国饲料 2020年19期
宣雄智, 李文嘉,朱文婷, 苏杰南
(1.苏州农业职业技术学院,江苏苏州 215008;2.河南省农业科学院畜牧兽医研究所,河南郑州 450002;3.广州市科沐生物科技有限公司,广东广州 510663)
随着人们生活水平的日益提高,对高质量鱼类蛋白的需求也越来越大,然而,当前水产养殖业的发展高度依赖以鱼粉和鱼油为代表的饲料原料(艾祖军等,2019)。因此,一方面,寻找可替代鱼粉的饲料蛋白源,另一方面,研发富含生物活性物质的绿色高效配合饲料以促进水产动物健康生长,提高水产品品质已成为现阶段水产养殖业发展的重要目标。
我国藻类资源丰富,尤其是海藻在饲料、食品和制药行业的应用受到广泛关注,其不仅可为水产动物提供蛋白质、脂肪与碳水化合物三大营养素,同时也是色素、水溶性和脂溶性维生素以及矿物质等生物活性化合物的天然来源 (吴晓娟等,2015)。研究表明,将不同种类、不同剂量的海藻粉添加到配合饲料中,可促进水产动物对营养物质的消化吸收、提高饲料利用率、抗应激性和免疫性能,还能一定程度改善鱼肉品质(陈明等,2016;李雅婷等,2016)。常见大型海藻主要有褐藻、红藻和绿藻三种类型,其中红藻的粗蛋白质含量高,活性物质种类丰富,比褐藻和绿藻更适合作为水产动物的饲料原料 (Fleurence等,2004)。龙须菜(Gracilaria lemaneiformis)是一种大型红藻,其粗蛋白质含量约占干物质的20%,还富含膳食纤维、微量元素等多种营养成分(陈晓凤,2012),广泛养殖于我国沿海地区,目前主要作为提取琼胶和卡拉胶的原料,还可以作为经济贝类鲍鱼的天然饵料及加工成即食海洋蔬菜。龙须菜作为饲料原料部分替代篮子鱼 (徐树德等,2014)、黑鲷(Xuan 等,2013)、真鲷(Xuan 等,2019)、异育银鲫(宣雄智等,2018)和草鱼(陈明等,2016)等水产动物饲料中的鱼粉或豆粕已被证明是可行的。然而龙须菜所含的碳水化合物多以纤维素、琼胶等非淀粉多糖形式存在,其不仅难以被鱼类有效利用,还可能对鱼类生长代谢产生负面影响。有研究表明,纤维素酶、木聚糖酶、β-葡聚糖酶等非淀粉多糖酶对藻类多糖有一定的体外酶解效果(李茜等,2013)。黄斑篮子鱼摄食含非淀粉多糖酶的石莼饲料后,生长性能和饲料利用率都有所提高(游翠红等,2014)。
翘嘴红鲌(Erythroculter ilishaeformis)属鲤形目,鲤科,鲌属,俗称太湖白鱼,是太湖著名河鲜“太湖三白”之一,分布于我国大部分水系。翘嘴红鲌生长快、肉质细嫩鲜美,颇受人们青睐。由于野生资源的减少,市场需求的不断增加,近年来翘嘴红鲌已成为长江中下游地区广泛养殖的名贵品种。
迄今为止,海藻在饲料中的应用多以海水杂食性或植食性鱼类为研究对象(宣雄智等,2018;陈明等,2016;游翠红等,2014;李茜等,2013),鲜有关于红藻替代饲料中的鱼粉对淡水肉食性鱼类的影响研究。因此,本试验通过在饲料中添加不同水平的龙须菜,研究其对翘嘴红鲌生长、体组成和代谢相关生化指标的影响,以判断龙须菜作为翘嘴红鲌饲料原料的可行性,同时,本试验探索了非淀粉多糖酶对摄食龙须菜饲料的翘嘴红鲌的影响。
1 材料与方法
1.1 试验饲料 本试验所用龙须菜购自福建宁德某公司。龙须菜经清洗后,置于户外阴干(约2周)。所有饲料原料粉碎后过60目筛,充分混匀后采用逐级扩大的方法添加维生素和矿物质等微量组分,然后加水混匀(饲料干物质和水分质量比约为7:3),随后用硬颗粒挤压机制成直径为1.5 mm的颗粒饲料,室温风干后密封于真空包装袋中,存放于-20℃冰箱中待用。以不含龙须菜藻粉的饲料为对照组,配制4组试验饲料,分别以2.5%、5%、7.5%、10%龙须菜等氮替代基础饲料中的鱼粉,编号为D1~D4,此外,以D4组饲料配方为基础,在其中添加0.2%非淀粉多糖复合酶,编号为D5。饲料组成及营养水平见表1。
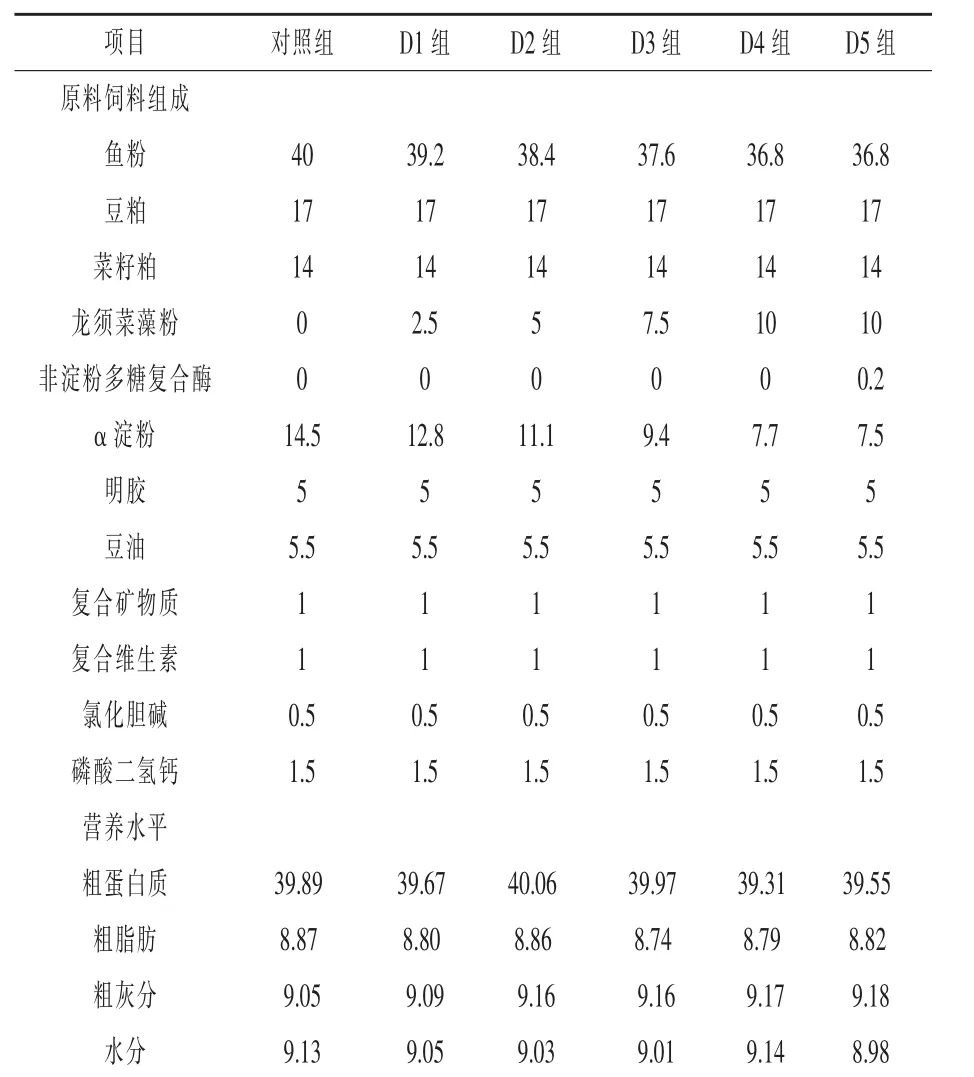
表1 饲料配方及营养水平(干物质)%
1.2 试验鱼和饲养试验 试验用翘嘴红鲌购自湖州某种苗场,试验开始前,先将试验鱼在室内水族箱 (规格70 cm×85 cm,300 L圆柱型)驯化2周,驯化期间投喂对照组饲料。养殖试验共6组,每组设3个平行。试验开始前,试验鱼禁食24 h,随机选择健康且规格基本一致的幼鱼用丁香酚(40 mg/L)麻醉,称体质量后放入水族箱中,每箱30 尾。 试验鱼初始体质量为(13.2±0.2)g,试验期间每日换水两次,换水量为总水量的1/3,水温为(25±2)℃,pH 为 7.5 左右;采用连续充气,水体溶氧≥5 mg/L;每天清晨用虹吸法吸除养殖缸底部的残饵与粪便。每天饱食投喂 2 次(8:30,16:30),具体以试验鱼不再抢食为准,记录投饲量,试验持续8周。养殖试验最后两周,给翘嘴红鲌投喂含1%Cr2O3的消化率试验饲料,饲喂 2 h后用虹吸法收集粪便,每次30 min。收集到的粪便用纱布过滤,60℃烘干,用于饲料表观消化率的测定。
1.3 样品收集 养殖试验结束后,鱼体禁食24 h,用40 mg/L丁香酚麻醉后逐尾称重。每缸随机挑取12尾,其中5尾麻醉致死,放入-20℃冰箱中用于全鱼成分分析。其余的鱼用不加抗凝剂的注射器从尾静脉采血,低温离心(1500 r/min,15 min,4℃)分离取上层淡黄色液体即为血清。离心后的血清经液氮速冻后置于超低温冰箱(-80℃)保存备用。将上述抽完血的试验鱼(每处理组共15尾)用于肝脏、前肠取样,所取样品装袋后经液氮速冻保存于超低温冰箱(-80℃)待用。
1.4 测定指标
1.4.1 常规生长及血清生化成分测定 饲料和全鱼样品的水分、粗蛋白质、粗脂肪、粗灰分含量,肝脏的水分和粗脂肪含量均按国标方法测定。全鱼和饲料蛋白成分通过凯氏定氮仪测定,粗脂肪用索氏抽提法测定;使用马弗炉在550℃下烘6 h测定灰分;使用烘箱105℃过夜测定样品中的水分;血清总蛋白、总胆固醇、甘油三酯、溶菌酶、谷草转氨酶和谷丙转氨酶的测定依照南京建成生物工程研究所试剂盒说明书进行。
试验鱼的增重率、饲料转化率、摄食率等指标计算公式如下:
增重率=(Wt-W0)/W0;
饲料转化率=(Wt-W0+Wd)/I;
摄食率/%=100×I/[t×(W0(g)+ Wt(g))/2];
成活率=末尾数/初尾数×100%;
蛋白质效率=(Wt-W0)/(I×Cp)×100%;
式中:Wt为试验结束时体质量,g;W0为试验初始体质量,g;t为试验时间,d;Wd为死亡鱼体质量,g;Cp为饲料中粗蛋白质含量;I为饲料干物质总投喂量,g。
肝体比、脏体比的计算公式为:
肝体比/%=肝脏鲜质量/鱼体鲜质量×100;
脏体比/%=内脏团鲜质量/鱼体鲜质量×100。
1.4.2 消化相关酶活性测定 酶液的制备及蛋白酶活性的测定参照宋林等(2013)所报道的方法,脂肪酶、淀粉酶和碱性磷酸酶活性均采用南京建成生物工程研究所生产的试剂盒测定,粗酶液中蛋白质浓度测定采用考马斯亮蓝 (南京建成生物工程研究所)染色法,具体测定方法参考试剂盒说明书。
1.4.3 表观消化率测定 表观消化率的测定参照Cho(1990)所报道的方法,本研究以 1%Cr2O3作为指示剂,对所配制的试验饲料组干物质和蛋白质的表观消化率进行了测定。具体计算如下:
表观消化率=[1-(A0/A×B/B0)]×100%;
式中:A、A0分别为饲料中和粪便中的营养物质含量;B、B0分别为饲料和粪便中的Cr2O3含量。
2 结果
2.1 龙须菜对翘嘴红鲌生长、体组成和饲料利用率的影响 养殖试验期间所有试验组翘嘴红鲌摄食率无显著差异,各组生长性能和饲料利用情况、表观消化率和全鱼及肝脏成分见表2~4。结果显示,本试验中,随着饲料中龙须菜替代比例的提高,翘嘴红鲌增重率呈先持平后下降的趋势而肝体比和脏体比则均呈下降趋势。与对照组相比,D3组和D4组增重率分别下降13.07%和14.07%(P<0.05),脏体比分别下降 11.19%和 11.72(P<0.05)。D4组全鱼粗脂肪含量比对照组下降10.4%(P<0.05),此外,该组干物质和粗脂肪表观消化率分别下降4.01%和3.79%(P<0.05)。
2.2 龙须菜对翘嘴红鲌消化相关酶活性和血液生化指标的影响 翘嘴红鲌消化相关酶活性和血液生化指标见表5~6。与对照组相比,D3组和D4组翘嘴红鲌前肠脂肪酶活性分别下降19.18%和19.88%(P<0.05),D4组前肠碱性磷酸酶活性也下降29.03%(P<0.05);血液指标方面,D1组翘嘴红鲌血液总蛋白含量上升15.18%(P<0.05),D3组和D4组血清胆固醇含量分别下降24.11%和33.69%(P < 0.05),D2组、D3组和 D4组血清甘油三酯分别下降20.79%、19.37%和27.06%(P<0.05)。随着龙须菜替代比例的提高,各试验组血清谷草转氨酶和谷丙转氨酶活性均呈上升趋势,但与对照组无显著差异。

表2 养殖8周后翘嘴红鲌的生长和饲料利用情况
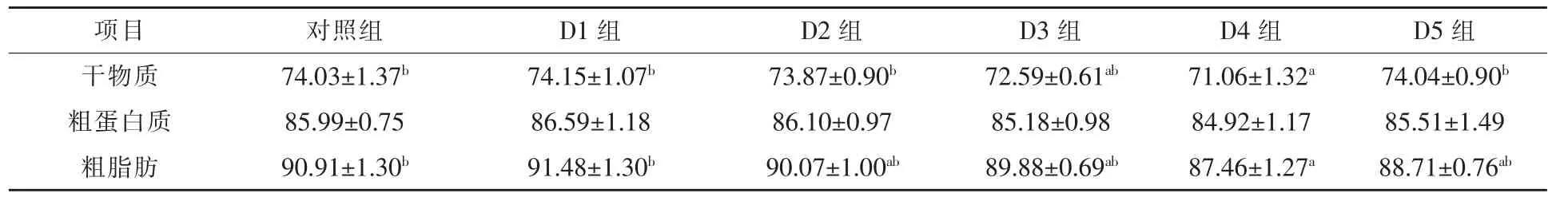
表3 养殖8周后翘嘴红鲌对龙须菜饲料的表观消化率%

表4 养殖8周后翘嘴红鲌全鱼及肝脏成分%
2.3 龙须菜饲料中添加非淀粉多糖酶对翘嘴红鲌的影响 在10%龙须菜饲料中添加0.2%非淀粉多糖酶后,D5组翘嘴红鲌摄食率和成活率比D4组分别提高9.85%和7.31%(P<0.05),与对照组均无显著差异(表2)。D5组饲料干物质、粗蛋白质和粗脂肪表观消化率均高于D4组,且干物质表观消化率比D4组提高了4.19%(P<0.05)(表3)。D5组前肠蛋白酶、脂肪酶和碱性磷酸酶活性分别比D4组提高 5.75%(P>0.05)、7.08%(P > 0.05)和 29.54%(P < 0.05)(表 5)。
3 讨论
3.1 龙须菜对翘嘴红鲌生长、体组成和饲料利用率的影响分析 龙须菜富含八种必需氨基酸、牛磺酸及多种必需微量元素,其藻胆蛋白具有抗氧化、抑制肿瘤等生理活性(余杰等,2006)。有研究表明用1%~5%的龙须菜饲料喂养眼斑拟石首鱼促进了该鱼的肠道健康,一定程度上提高了其增重率(李雅婷等,2016),本试验中,龙须菜藻粉替代比例不超过5%时,对翘嘴红鲌的生长性能无显著影响,与宣雄智等(2018)的研究结果相一致,但当龙须菜替代比例达到或超过7.5%时,翘嘴红鲌增重率显著下降。有研究指出,用龙须菜饲料饲喂黑鲷 (Xuan等,2013)、真鲷 (Xuan等,2019)和异育银鲫(宣雄智等,2018)时,其最大替代比例均可达15%,而对养殖鱼类的生长和摄食无负面影响。产生这种差异的原因可能是由于鲷科鱼类为海洋杂食偏肉食性鱼类,海藻是其天然饵料的组成部分,其对海藻有较好的消化能力(Nakagawa,1997),异育银鲫是淡水杂食偏植食性鱼类,其对植物蛋白源的接受度也较高 (王永玲等,2011),而翘嘴红鲌属于淡水肉食性鱼类,营养需求高,对植物蛋白源营养成分的消化率较低(蒋蓉等,2005),本试验中龙须菜替代比例达10%时,饲料干物质表观消化率的下降也说明了这一点。

表5 养殖8周后翘嘴红鲌肝脏和前肠消化酶活性U/mg protein
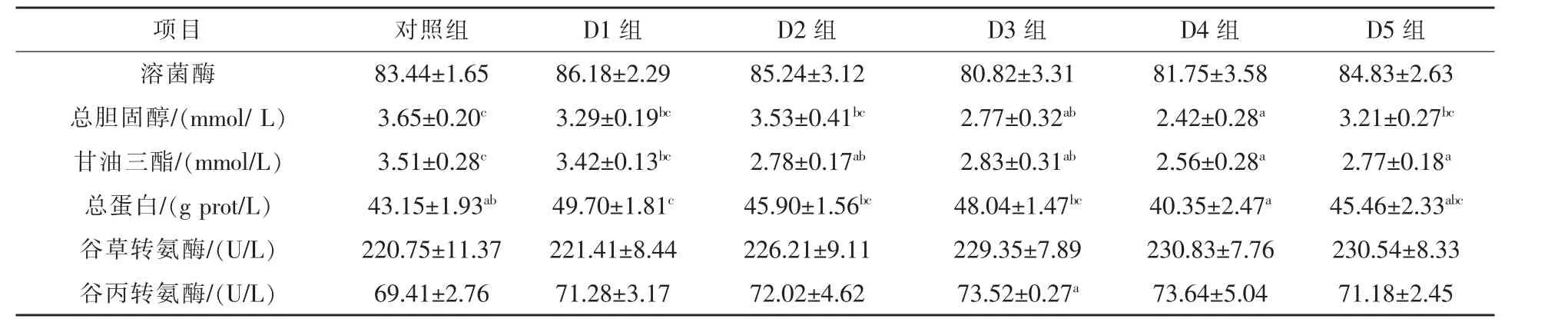
表6 养殖8周后翘嘴红鲌的血清指标
研究表明,在饲料中添加大型海藻或者微藻会对水产动物的体脂代谢产生影响,这种影响因藻类和养殖鱼类的种类差异而不同 (Xuan等,2019;宣雄智等,2018;Xuan 等,2013)。 本试验中D4组翘嘴红鲌全鱼脂肪含量显著下降,这可能是由以下原因所致,一方面,虽然本试验所用各组饲料是等氮等脂的,但饲料碳水化合物的种类不同,龙须菜粗多糖含有较多的胶体,不能被养殖鱼类有效利用(朱地琴等,2012),因此,随着龙须菜替代比例的提高,试验组饲料的可消化能逐渐降低,另一方面龙须菜富含的琼胶多糖有较高的黏性,能在一定程度上吸附油脂(宣雄智,2014),从而抑制了翘嘴红鲌对饲料脂肪的吸收,降低了粗脂肪表观消化率。本试验中,各试验组全鱼粗蛋白质含量、蛋白质效率和粗蛋白质表观消化率未受显著影响,这表明龙须菜蛋白质氨基酸比例平衡(Xuan 等,2019;徐树德等,2014),能被翘嘴红鲌较好地利用。
当龙须菜替代比例大于7.5%时,翘嘴红鲌脏体比显著降低,而肝体比却未受显著影响,这与龙须菜饲料饲喂黑鲷的试验结果不同 (宣雄智,2014),表明龙须菜饲料能影响翘嘴红鲌的脂肪代谢且腹腔脂肪含量比肝脏脂肪含量更易受影响。3.2 龙须菜对翘嘴红鲌消化相关酶活性和血液生化指标的影响分析 鱼类消化酶主要可分为蛋白酶、酯酶、碳水化合物分解酶三大类(张伟等,2007)。本试验中,各试验组翘嘴红鲌前肠蛋白酶、脂肪酶和淀粉酶的活性远高于肝胰脏相关酶的活性,可能是由于翘嘴红鲌为鲤科无胃鱼类,其肝胰脏分泌的酶原没有活性,进入肠道后才被激活(Das等,1991)。翘嘴红鲌肠道消化酶活性的大小将影响其对饲料营养物质的消化吸收,当龙须菜替代比例超7.5%时,翘嘴红鲌前肠脂肪酶活性显著下降,这一结果也与粗脂肪表观消化率和全鱼脂肪含量的下降相呼应。
碱性磷酸酶是存在于鱼类前肠上皮细胞浅部和纹状缘上的一种协助肠上皮细胞吸收的金属酶,与脂类、葡萄糖、钙和无机磷的吸收状况呈正相关,可表征肠细胞的分化程度和肠道消化功能的完善程度(张伟,2008)。本试验中,龙须菜替代比例达10%时,翘嘴红鲌碱性磷酸酶活性显著下降,推测龙须菜所含的多糖类膳食纤维可能抑制了翘嘴红鲌末端肠上皮细胞的消化代谢。
血清蛋白质主要维持胶体渗透压,还具有运输载体、免疫和代谢调控等作用。本试验中,龙须菜替代比例为2.5%时,翘嘴红鲌血清总蛋白含量显著上升,与宣雄智等(2018)用龙须菜饲喂异育银鲫的结果一致,这可能与龙须菜含有藻胆蛋白、多糖等具有益生素功能的物质有关 (陈美珍,2005),而血清蛋白质含量升高有利于提高翘嘴红鲌的代谢水平和免疫能力(赵红霞等,2008)。
一般认为,肝脏有炎症或肝细胞发生病变、坏死时,血清谷草转氨酶和谷丙转氨酶活性升高(宣雄智等,2018)。本试验中,随着龙须菜替代比例的提高,翘嘴红鲌血清谷草转氨酶和谷丙转氨酶活性虽呈上升趋势,但与对照组差异不显著,且肝胰脏各消化酶活性也未发生明显变化,因此翘嘴红鲌肝功能可能未受龙须菜饲料的影响。
鱼类血清中脂类主要包括:磷脂、胆固醇、甘油三酯、非酯化脂肪酸四大类。血脂水平可以间接反映全身脂质代谢状况(Nakagawa,1997)。本试验中,当龙须菜替代比例达到或超过5%时,翘嘴红鲌血清甘油三酯含量显著低于对照组,部分研究也发现当饲料中龙须菜替代达一定比例时,养殖鱼类血清甘油三酯含量会显著下降 (陈明等,2016;游翠红等,2014),这可能是由于龙须菜琼胶多糖在肠道中吸水膨胀,食糜黏性增加,胃排空减慢,间接导致饲料脂肪吸收减少 (倪德江等,2004)。血液总胆固醇含量可间接反映鱼体脂类吸收及肝脏脂肪代谢状况(Nakagawa,1997)。本试验中,当龙须菜替代比例达到或超过7.5%时,翘嘴红鲌血清胆固醇含量显著下降,不少研究表明鱼类摄食含植物蛋白源饲料后血液胆固醇含量下降(陈明等,2016;游翠红等,2014),可能是由于胃肠道对胆固醇的吸收受到抑制,也可能由于龙须菜多糖调节脂代谢基因表达,加快胆汁酸的排泄,从而降低体内胆固醇含量(黄诗铭,2019)。
3.3 龙须菜饲料中添加非淀粉多糖酶对翘嘴红鲌的影响分析 一般认为,水产动物不能分泌纤维素酶、β-葡聚糖酶和木聚糖酶等非淀粉多糖酶,大型海藻所含的碳水化合物主要是海藻胶、粗纤维类等非淀粉多糖,难以被水产动物消化(游翠红等,2014),特别是细胞壁中的纤维素和果胶不仅难以消化,还会影响肠道消化酶对饲料营养物质的分解,降低饲料利用效率 (宣雄智,2014)。非淀粉多糖酶则具有消除抗营养因子,提高饲料利用率,促进动物生长的作用(游翠红等,2014)。体外研究表明,龙须菜可被纤维素酶、木聚糖酶、β-葡聚糖酶酶解(李茜等,2013),游翠红等(2014)在石莼饲料中添加非淀粉多糖酶,有效提高了黄斑篮子鱼的生长效果和饲料利用率。本试验发现与摄食10%龙须菜饲料的试验组相比,添加非淀粉多糖酶可以显著提高翘嘴红鲌的干物质表观消化率、前肠碱性磷酸酶活性、增重率和存活率,这可能是由于非淀粉多糖酶的添加改善了肠道的生理状况,促进了翘嘴红鲌肠道对饲料营养物质的消化吸收,减轻了龙须菜多糖等抗营养因子带来的负面影响。本试验中,翘嘴红鲌的摄食率未受龙须菜饲料的影响,但在10%龙须菜饲料中添加非淀粉多糖酶后,翘嘴红鲌摄食率显著升高,这可能是由于龙须菜含有呈味氨基酸等诱食性物质(宣雄智,2014),另一方面在非淀粉多糖酶作用下,龙须菜所含膳食纤维被肠道菌群发酵,产生酸性物质,刺激了肠道黏膜,加速了肠道排空。
4 结论
用2.5%的龙须菜替代饲料中的鱼粉有助于提高翘嘴红鲌的非特异性免疫,当龙须菜替代比例达7.5%时,翘嘴红鲌生长性能显著下降;进一步提高龙须菜替代比例至10%时,翘嘴红鲌对饲料营养物质的消化吸收能力和生长性能均会受抑制。饲料中添加龙须菜能显著影响翘嘴红鲌的脂肪代谢,降低全鱼脂肪含量。非淀粉多糖酶的添加可以一定程度改善龙须菜饲料对翘嘴红鲌产生的负面影响,提高龙须菜在饲料中的添加比例,但具体的添加量仍需进一步研究。
猜你喜欢
农业技术与装备(2022年6期)2022-08-17
世界科学技术-中医药现代化(2021年8期)2021-12-21
河北果树(2021年4期)2021-12-02
中国畜牧杂志(2021年5期)2021-05-17
国际种业前沿动态(2020年16期)2020-10-14
三农资讯半月报(2020年16期)2020-09-14
农药科学与管理(2019年6期)2019-11-23
农药科学与管理(2019年6期)2019-11-23
湖南饲料(2019年5期)2019-10-15
福建基础教育研究(2019年10期)2019-05-28
