
有机微量元素和酵母培养物替代氧化锌对断奶仔猪生长性能和肠道健康的影响
2020-11-15 11:32晏家友张锦秀黄崇波魏小兰周梦佳
中国饲料 2020年19期
刁 慧 , 晏家友 , 张锦秀 ,3, 张 纯 , 黄崇波 ,4, 魏小兰 , 周梦佳 ,4
(1.四川省畜牧科学研究院,四川成都 610066;2.动物遗传育种四川省重点实验室,四川成都 610066;3.四川省畜科饲料有限公司,四川成都 610066;4.畜禽生物制品四川省重点实验室,四川双流616200)
锌对大部分生物包括猪是有毒的重金属,锌离子浓度过高会产生极大的细胞毒性(Nies,2007)。研究发现,连续12周以上每天摄入超过50 mg Zn可导致铜缺乏 (Yu等,2013)。添加2500 mg/kg氧化锌会对仔猪肝脏产生毒性效应,肝细胞质内出现空泡,脂肪沉积增多(Jensenwaern等,1998)。体外研究发现,高剂量的氧化锌可能会对CACO-2细胞产生毒性,使通透性增加(Roselli等,2003)。此外,日粮中添加2000~3000 mg/kg氧化锌可显著增加粪样中Zn的排放量(Meyer等,2002)。在农场施用富Zn粪肥后,会在土壤中逐渐蓄积,这不仅对农作物有潜在的危害,对人体健康也有潜在的威胁(Jondreville等,2003)。2018年1月农业部2625公告对锌的用量做了进一步限制,由此可见,高剂量氧化锌的禁用将是未来发展的趋势,因而寻求高剂量氧化锌的替代方案,是养猪业亟待解决的重要问题之一。
酵母培养物作为一种微生态制品,是由酵母菌在特定工艺条件下发酵而来,含有丰富的营养物质和代谢产物,包括矿物元素、维生素、寡糖、小肽、有机酸和未知的生长因子等,具有提高饲料利用率、改善动物胃肠道健康和提高免疫力等功能(Upadhaya等,2019;Lee 等,2018)。同时,酵母培养物可以通过调节断奶仔猪肠道热休克蛋白和细胞因子的表达来缓解热应激引发的肠道绒毛损伤进而维持生长,减轻应激对动物的不利影响(Kumar等,2017)。有机微量元素适口性好,具有稳定的化学性能、较高的吸收率和生物学效价,对于改善仔猪的肠道健康、提高吸收利用率、免疫力和抗应激能力以及减少金属排放至关重要(刘丹,2007)。因此,有机微量元素作为安全高效环保型饲料添加剂越来越受到大家的关注。目前,关于有机微量元素和酵母培养物联合使用能否可以替代高剂量氧化锌在断奶仔猪上的研究鲜见报道。本试验采用有机微量元素和酵母培养物替代日粮中氧化锌,探讨其对断奶仔猪生产性能、锌排放和肠道健康的影响,阐明其作用效果及可行性,为深入认识有机微量元素和酵母培养物的营养生理作用提供参考,为提供高剂量氧化锌的替代方案及减少锌排放对环境的压力提供理论依据,为缓解断奶仔猪应激的营养措施提供新的思路。
1 材料与方法
1.1 试验动物与设计 选择48头28日龄、体重为(9.11±0.44)kg 的健康杜×长×大断奶仔猪,按照体重一致、公母均匀的原则和完全随机区组的设计分为2个处理(表1),即基础饲粮(微量元素均为无机微量元素,不加锌)+1600 mg/kg氧化锌(以锌计);基础饲粮(微量元素为复合有机微量元素)+3000 mg/kg酵母培养物。每个处理6个重复,每个重复4头猪。试验期共14 d。

表1 试验设计
1.2 试验材料 氧化锌市场购置,氧化锌含量≥95.0%,锌含量≥76.3%;酵母培养物(益康XP)由美国达农威公司提供;复合有机微量元素由四川省畜科饲料有限公司提供。
1.3 试验饲粮及饲养管理 试验饲粮为玉米-豆粕型饲粮,参考 NRC(2012)7~11 kg阶段猪营养需要配制。基础饲粮组成及营养水平见表2。每天饲喂4 次(08:00,12:00,16:00,20:00),少喂勤添,喂量以料槽内略有剩余为准,各组饲养管理条件一致。保持圈舍通风、清洁、干燥、卫生。每日注意观察其行为表现、采食和饮水情况,发现异常及时记录并处理。期间不使用任何抗生素以及微生态制剂。
1.4 样品采集
1.4.1 饲料样 通过四分法采集饲料样品,每个处理取150 g左右,装入样品袋并记号,放入-20℃冰箱保存待测养分的含量。

表2 基础饲粮组成及营养水平(风干基础)
1.4.2 粪样 于试验第11~14天,采用内源指示剂收粪法,进行消化试验,共4 d,进行直肠粪样收集。每次收集后加10%硫酸和几滴甲苯进行固氮和防腐。每头猪收集的粪便经充分混合后,取适量在样品盘中,于60~65℃烘至恒重。样品干燥后粉碎,过40目筛,储存于-20℃,用以测定粪中常规养分的含量。
1.4.3 血样和组织样 试验第14天,每个重复选择一只接近平均体重的猪只空腹后前腔静脉采血5 mL,室温下倾斜放置30 min,3000 r/min离心10 min,分离血清,置于-20℃冷冻保存。采血后立即屠宰,打开腹腔,迅速分离肠段,取完好无损的十二指肠、空肠和回肠保存于4%多聚甲醛溶液中,用于肠道形态分析和杯状细胞数量的测定;取空肠黏膜测定肠道发育和屏障相关基因表达。黏膜样品放入液氮冻存,并及时转入-80℃冰箱保存待测。
1.5 测定指标及方法
1.5.1 生长性能 于试验开始前和第14天,分别对每头仔猪进行空腹称重。试验期间准确记录每天每圈的投料量和剩料量(余料量+浪费量),实际采食量=投料量-剩料量。根据平均日增重(ADG)和平均日采食量(ADFI)计算料重比(F/G)。
1.5.2 腹泻评分 每天早晚对所有猪只进行腹泻评分,腹泻评分标准如下:0分,坚硬的条形或粒状;1分,软便、能成形;2分,浆糊状、不成形;3分,液状、粪水分离。将得分为2分和3分者确定为腹泻。统计仔猪腹泻头次数,并计算腹泻率,其中腹泻率/%=[腹泻头次数/(仔猪头数×试验天数)]×100。
1.5.3 粪锌含量 采用原子吸收光谱法测定样品中锌含量,样品前处理方法为浓硝酸、高氯酸消化法。具体测定方法为:准确称取1 g左右(精确至0.0001 g)的样品放入250 mL锥形瓶内,加入20 mL左右的浓硝酸,摇匀并静置过夜,缓慢加热进行消化。当液体快蒸干时取下锥形瓶冷却后加入5 mL高氯酸继续消化,待锥形瓶内液体呈现无色、无烟冒出时消化结束。冷却后用双蒸水在100 mL容量瓶中定容,原子吸收光谱仪待测。
1.5.4 养分消化率 内源指示剂盐酸不溶灰分的测定方法参见标准“饲料中盐酸不溶灰分的测定”(GB/T23742)。饲料及粪便中的干物质、粗蛋白质、粗脂肪和粗灰分的检测参照张丽英(2003)的方法。饲粮养分消化率的计算公式为:
某养分消化率/%=100-A1F2/A2F1×100;
式中:A1为饲料中盐酸不溶灰分的含量,%;A2为粪中盐酸不溶灰分的含量,%;F1为饲料中某养分的含量,%;F2为粪中某养分的含量,%。
1.5.5 小肠组织形态学 取保存于4%多聚甲醛溶液中固定8~24 h的十二指肠、空肠和回肠,然后经脱水、透明和包埋等制作石蜡切片,切片厚度为10 μm。随后对切片进行苏木精和伊红染色,封片后在显微镜下观察,并测定绒毛高度和隐窝深度。采用阿新兰-过碘酸雪夫氏(AB-PAS)组织化学染色,通过Olympus光学显微镜对空肠杯状细胞计数。1.5.6 肠道发育、肠道消化吸收和肠道屏障相关基因mRNA相对表达量 实时荧光定量PCR测定:(1)肠道发育相关基因:表皮生长因子(EGF)和胰岛素样生长因子 1(IGF-1);(2)肠道增殖与凋亡相关基因:促凋亡基因(Bax)和抗凋亡基因(Bcl-2);(3)肠道消化吸收相关基因:钠葡萄糖共转运载体 1(SGLT-1)、葡萄糖转运蛋白-2(GLUT-2)、碱性氨基酸转运载体(SLC7A1)、锌离子转运载体(ZNT1)、锌离子转运载体(ZNT2)和锌离子转运载体(ZIP4);(4)肠道屏障相关基因:闭锁蛋白(Occludin)、黏蛋白 1(MUC1)、黏蛋白 2(MUC2)、白介素 1β(IL-1β)和白介素 10(IL-10)的mRNA相对表达量。
用Trizol(宝生物工程有限公司)提取组织总RNA,具体步骤如下:将空肠样品迅速放入预冷好的研钵中充分研磨成粉末,取50 mg左右的粉末加入到含有1 mL Trizol的1.5 mL离心管中,随后剧烈振荡打散直至溶解;静置5 min,离心10 min(4 ℃,12000 g),取上清液,加入 200 μL 氯仿,剧烈振荡5 min,静置5 min,离心15 min(4℃,12000 g);转移上清液至另一1.5 mL离心管中,加入1:1的异丙醇,室温放置 15 min,离心 10 min(4 ℃,12000 g),弃上清;加入 1 mL 75%乙醇,用手指轻弹使沉淀悬浮,离心 15 min(4℃,7500 g),弃上清,室温放置1~2 min晾干沉淀;待RNA略干,加入40 μL RNase-free水;待RNA充分溶解后,用1%琼脂糖凝胶电泳 (100 V,15 min)检查RNA相对分子量大小和完整性,在紫外分光光度计(Beckman DU-800,布雷亚,美国)上检测 A260nm和A280nm,根据OD260/OD280判断RNA纯度。
参照反转录试剂盒(宝生物工程(大连)有限公司)说明书配制反转录混合液1和2。加入反转录混合液1及cDNA后在PCR仪上除去基因组DNA(42 ℃,2 min),随后加入反转录混合液 2,经程序37℃15 min,85℃5 s完成反应。本试验采用10 μL荧光定量PCR反应体系,包括1 μL cDNA,上下游引物各 0.5 μL,5 μL SYBR Premix Ex TaqTM和3 μL RNase Free H2O。反应循环条件为:95 ℃ 30 s,95 ℃ 10 s,60 ℃ 25 s, 共 40 个循环,引物序列见表3,以β-actin作为内参基因进行计算组织中各基因的相对表达量。
1.6 数据处理 所有数据均用Excel 2010进行初步整理,采用SAS 8.0软件中的t检验对数据进行统计分析,P<0.05为差异显著,0.05≤P<0.10视为有趋势。
2 结果与分析
2.1 替代氧化锌对断奶仔猪生长性能和腹泻率的影响 由表4可知,与氧化锌组相比,替代组断奶仔猪1~14 d的平均日采食量和平均日增重显著降低(P<0.05),同时1~14 d的料肉比显著降低(P<0.05),饲料转化率得到改善。与氧化锌组相比,替代组仔猪腹泻率显著提高(P<0.05)。

表3 实时荧光定量PCR引物序列

表4 氧化锌替代对断奶仔猪生长性能和腹泻率的影响
2.2 替代氧化锌对断奶仔猪养分消化率的影响由表5可知,与氧化锌组相比,替代组断奶仔猪干物质、粗灰分和Zn的表观消化率显著提高 (P<0.05)。有机微量元素和酵母培养物替代氧化锌有提高断奶仔猪粗蛋白质养分消化率的趋势 (P=0.063)。与氧化锌组相比,替代组断奶仔猪粪中Zn含量显著降低(P<0.05)。

表5 氧化锌替代对断奶仔猪养分消化率和锌排放的影响
2.3 替代氧化锌对断奶仔猪肠道形态和杯状细胞数量的影响 由表6可知,与氧化锌组相比,替代组断奶仔猪十二指肠隐窝深度显著降低,绒隐比显著提高 (P<0.05),空肠绒毛高度显著增加 (P<0.05)。有机微量元素和酵母培养物替代氧化锌可显著增加断奶仔猪空肠杯状细胞的数量(P<0.05)。
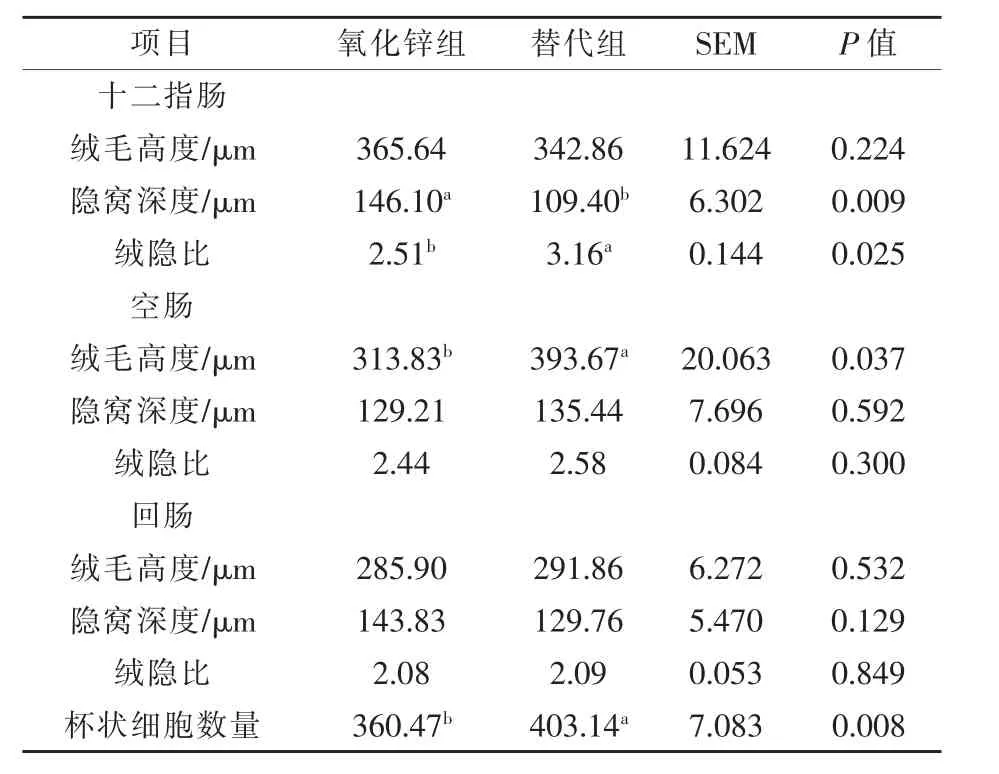
表6 氧化锌替代对断奶仔猪小肠形态结构和杯状细胞数量的影响
2.4 替代氧化锌对断奶仔猪空肠转运载体基因表达的影响 由图1可知,与氧化锌组相比,替代组空肠ZNT1和ZNT2的mRNA相对表达量显著降低(P<0.05),ZIP4的mRNA相对表达量显著升高(P<0.05)。有机微量元素和酵母培养物替代断奶仔猪饲粮中氧化锌对空肠GLUT-2、SGLT-1和SLC7A1 mRNA相对表达量无显著影响(P>0.05)。
2.5 替代氧化锌对断奶仔猪空肠肠道发育相关基因表达的影响 如图2所示,与氧化锌组相比,替代组空肠IGF-1 mRNA相对表达量显著升高(P<0.05),Bax mRNA 相对表达量显著降低(P<0.05)。与氧化锌组相比,有机微量元素和酵母培养物替代氧化锌有提高空肠EGF(P=0.066)mRNA相对表达量的趋势。
2.6 替代氧化锌对断奶仔猪空肠肠道屏障相关基因表达的影响 由图3可知,与氧化锌组相比,替代组空肠IL-1β mRNA相对表达量显著降低(P<0.05),MUC2 mRNA相对表达量显著升高(P<0.05)。有机微量元素和酵母培养物替代断奶仔猪饲粮中氧化锌对空肠IL-10、MUC1和Occludin mRNA相对表达量无显著影响(P>0.05)。
3 讨论
3.1 生长性能 研究表明,饲粮添加有机微量元素能显著提高日增重和饲料报酬,相对低剂量有机微量元素能使动物生产性能达到或优于高剂量无机微量元素。Woolen等(1990)给仔猪人工感染痢疾,结果发现饲粮中添加40 mg/kg蛋氨酸锌可降低仔猪死亡率。韩友文等(2000)以赖氨酸微量元素鳌合物饲喂幼龄仔猪,试验结果显示相比无机微量元素,日增重提高了6.4%,饲料效率得到改善。育肥猪上的研究得到了与之一致的结果,饲粮中添加复合氨基酸鳌合盐可以提高日增重,改善饲料效率 (刘桂琴和陈顺,1994)。Creech等(2004)报道,与无机微量元素相比,低剂量有机微量元素日粮有提高后备母猪平均日增重和平均日采食量的趋势。肉鸡上的研究同样发现,饲粮中添加复合有机微量元素可显著提高肉仔鸡的生长性能,特别是在生长期(Baloch等,2017)。无机微量元素在动物机体吸收过程中必须借助辅酶与氨基酸或其他物质形成螯合物或络合物,而有机微量元素被吸附到氨基酸、肽或其他化合物上后更容易被吸收。因此,有机微量元素的吸收率是无机微量元素的1.8~4.0倍,更利于生长性能的提高(呙于明,2004)。
酵母培养物是一种含有多种代谢产物的发酵产品。Shen等(2009)在断奶仔猪上开展酵母培养物梯度试验 (0、2.5、5、10、20 g/kg), 研究结果表明,饲粮中添加5 g/kg酵母培养物可显著提高仔猪平均日增重。吴徐俊(2016)报道指出,饲粮中添加1%啤酒酵母培养物可显著提高断奶仔猪的平均日增重和平均日采食量。Liu等(2018)研究发现,饲粮中添加5 g/kg酵母培养物可显著增加仔猪断奶后1~14 d平均日增重和平均日采食量,改善试验全期饲料转化率,说明酵母培养物对仔猪断奶后前两周效果非常显著。Upadhaya等(2019)研究表明,断奶仔猪饲粮中添加4 g/kg马克斯克鲁维酵母和酿酒酵母混合培养物可提高断奶仔猪试验第一周和全期的平均日增重。母猪上的研究表明,用2 kg/d酵母培养物(益康XPC)饲喂妊娠期和泌乳期母猪,可以提高窝增重,改善其繁殖性能和后代生长性能(Kim等,2008)。家禽上的研究同样发现酵母培养物能改善饲料转化效率,提高生产性能(Firman 等,2013)。
本试验研究结果表明,与氧化锌组相比,有机微量元素和酵母培养物替代高剂量氧化锌可以显著降低断奶仔猪1~14 d的平均日采食量和平均日增重,增加腹泻率,但1~14 d的料肉比显著降低,饲料转化率得到改善。这与本试验的设计有很大关系。本试验的研究对象为处于断奶应激的仔猪,选择的对照组不是没有添加氧化锌的空白组,而是以添加了高剂量的氧化锌作为对比。因此,有机微量元素和酵母培养物替代组取消了高剂量的氧化锌,势必会对采食量和腹泻造成一定的影响。但值得注意的是本研究发现有机微量元素和酵母培养物联合使用替代氧化锌后仔猪的饲料转化率得到改善,这可能和肠道的功能得到改善有关。
3.2 养分消化吸收 机体健康受到肠道功能和发育的影响,而肠道的生长、消化吸收功能和屏障完整性共同筑成肠道的发育。在本试验中,有机微量元素和酵母培养物替代氧化锌可提高断奶仔猪干物质、粗灰分、Zn和粗蛋白质的表观消化率,降低粪中锌的排放量,这与前人研究一致。Shen等(2009)在断奶仔猪上的研究发现,饲粮添加5 g/kg酵母培养物显著提高了仔猪干物质、粗蛋白质和总能的表观消化率。Upadhaya等(2015)研究表明,饲粮中添加2、3 g/kg酵母培养物显著提高了仔猪断奶后14 d的干物质和能量表观消化率。母猪上有关酵母培养物的研究也得到了类似的结果(汤飞飞,2012)。大量研究表明,饲粮添加有机微量元素同样可以提高养分消化率。与无机微量元素组相比,复合寡糖微量元素络合物组育肥猪的粗脂肪、粗灰分、钙和磷的表观消化率显著提高,铜、锌的粪排泄显著降低(周颖等,2015)。Seyfori等(2019)在鸵鸟上的研究结果发现,饮水中添加有机微量元素可提高能量、粗纤维和粗脂肪的表观消化率。Ma等(2019)在母猪上的研究发现,饲粮添加复合有机微量元素可显著降低微量元素的排放量。由此说明有机微量元素和酵母培养物联合使用替代氧化锌可以提高断奶仔猪养分消化率,这与饲料转化率的改善相呼应。同时,氧化锌替代后可以降低粪中微量元素特别是锌的排放量,从而缓解重金属排放对环境造成的压力。此外,本研究发现与氧化锌组相比,替代组空肠ZNT1和ZNT2的mRNA相对表达量显著降低,ZIP4的mRNA相对表达量显著升高。ZnT转运蛋白可以协助锌离子从细胞质内流出细胞外或转运到细胞器内降低细胞质锌浓度,ZIP家族转运蛋白通过转运锌离子内流进入细胞或者通过从细胞内囊泡或细胞器转运出锌离子,增加胞内锌浓度(Kambe等,2004)。体外研究试验表明,与空白组相比,添加50、100、200 mg/L乳酸锌可以上调ZnT2 mRNA表达,下调ZIP4 mRNA表达(韩国全等,2012),这与本研究结果一致。
3.3 肠道形态 仔猪断奶由吮吸母乳过渡到采食固体饲料,容易导致小肠绒毛萎缩和隐窝深度增加,进而使成熟的小肠绒毛上皮细胞数量减少和生长速度减慢,影响仔猪消化能力(Pluske等,1996)。酵母培养物作为一种发酵型物质,可以改善肠道形态、维护肠黏膜完整性,对维持动物肠道健康有积极意义。Shen等(2009)研究表明,饲粮中添加5 g/kg酵母培养物可显著提高断奶仔猪空肠绒毛高度和绒隐比。汤飞飞(2012)研究表明,仔猪饲粮中添加 1、2、4 g/kg酵母培养物可显著提高35日龄仔猪空肠绒毛高度。空肠绒毛高度提高说明仔猪肠道形态得到改善,这使得肠道的消化和吸收能力更强,生产性能更佳(Price等,2010)。酵母培养物的补充导致更浅的隐窝深度,表明细胞更新和周转代谢的必要性更小,从而降低宿主维持肠上皮细胞周转代谢的能量需要 (Gao等,2008)。有机微量元素同样可以影响仔猪的肠道形态结构完整性。研究表明,有机锌可以改善生长猪回肠肠道形态结构,维持上皮形态完整性(Pearce等,2015)。 Echeverry 等(2016)研究发现,饲粮添加复合有机微量元素可显著提高肉鸡回肠绒毛高度,降低隐窝深度。El-Katcha等(2017)研究表明,饲粮中添加45 mg/kg或60 mg/kg多糖锌可提高肉鸡肠道绒毛高度,降低隐窝深度,改善肠道形态。饮水中添加有机微量元素得到了相同的结果(Seyfori等,2019)。本试验发现,有机微量元素和酵母培养物替代氧化锌可显著提高断奶仔猪十二指肠绒隐比和空肠绒毛高度,降低隐窝深度,同时提高空肠IGF-1和EGF mRNA相对表达量,降低Bax mRNA相对表达量,这与前人的研究结果相吻合。研究发现,Bax基因表达的升高和细胞凋亡呈正相关(Hass等,1997),一些参与肠道上皮生长调控的分子肽类生长因子,包括IGF-1和EGF等,都能参与调节肠道的增殖、分化和生长(Steeb等,1994)。说明有机微量元素和酵母培养物联合使用替代氧化锌可以改善断奶仔猪肠道形态结构,促进肠道生长发育,进而提高养分消化率,改善饲料报酬。
3.4 肠道屏障 肠道屏障功能的完整性对机体养分消化吸收功能及肠道发育具有重要的作用。研究表明,饲粮中添加5 g/kg酵母培养物显著提高了仔猪空肠IFN-γ的含量,降低血清中CD4的含量,对调节肠道免疫应答有积极的作用,说明酵母培养物通过调节机体免疫功能改善保育猪生长性能(Shen等,2009)。 Weedman 等(2011)考察酿酒酵母培养物对仔猪断奶应激和运输应激的影响,研究结果发现,1 g/kg酿酒酵母培养物通过影响TLR信号通路调控断奶仔猪免疫功能,同时调节肠道微生物菌群的多样性和组成,共同缓解应激对仔猪造成的不利影响。仔猪饲粮中添加1、2、4 g/kg酵母培养物均可显著降低70日龄仔猪空肠IL-4浓度,说明酵母培养物能增强Th-1型免疫应答,对仔猪免疫机能有提高作用(汤飞飞,2012)。在饲粮中添加酵母培养物,可能有助于清除来自受感染动物的病原体。一项关于接种沙门氏菌的研究表明,仔猪饲粮中添加酵母培养物可以使病原体迅速从肠道脱落(Price等,2010)。在肉鸡饲料中添加酵母培养物也可以增强适应性免疫系统的T淋巴细胞,进而更好地清除病原体(Gao等,2009)。酵母培养物成分复杂,低聚糖是其中的一种。研究发现,低聚糖可以激活吞噬细胞、自然杀伤细胞、B淋巴细胞和T淋巴细胞,增加细胞因子的产生和巨噬细胞的吞噬作用 (Hoffman等,1993)。这些分子的存在会发挥免疫调节作用抑制病原体,阻止其附着肠黏膜上,这不仅保护了宿主不受病原体的侵害,而且也允许宿主的能量储备被用于自身的生长而不是肠上皮修复和再生(Gao等,2008)。研究发现,有机微量元素同样参与调控机体肠道免疫屏障。与氧化锌组相比,饲粮中补充蛋氨酸锌可显著提高绵羊的布氏杆菌试管凝集反应抗体效价,增加血清γ球蛋白含量、淋巴细胞染色阳性率和外周淋巴细胞转化率,说明有机锌提高绵羊机体细胞免疫和体液免疫能力优于无机锌(邵凯,2000)。Pearce 等(2015)研究表明,与 120 mg/kg硫酸锌组相比,60 mg/kg有机锌 (氨基酸锌)可以降低生长猪内毒素的产生,缓解热应激带来的不利影响。Echeverry等(2016)研究发现,在肉鸡饲粮中添加复合有机微量元素可以显著上调回肠TLR2、TLR4和IL-10的基因表达,说明复合有机微量元素可以调节toll样受体和抗炎细胞因子,从而对肠道免疫屏障系统产生有益的影响,并可同时改善健康状态。本试验研究发现,与氧化锌组相比,替代组空肠IL-1β mRNA相对表达量显著降低,MUC2 mRNA相对表达量显著升高,同时杯状细胞数量增加。肠道黏液层位于肠黏膜和肠腔食糜之间,主要由杯状细胞分泌的黏蛋白构成,参与肠道化学屏障的组成,且MUC2是众多黏蛋白中最为主要的一种黏蛋白(Lamont,1992)。 因此,可以推测,有机微量元素和酵母培养物通过上调肠道上皮细胞抗菌蛋白基因的表达,下调促炎性因子的基因表达进而维护肠道屏障功能。
4 结论
有机微量元素和酵母培养物替代氧化锌可降低断奶仔猪平均日采食量和平均日增重,增加腹泻率,但同时可提高饲料转化率和养分消化率,改善肠道形态结构,维持肠道屏障功能。
猜你喜欢
新乡医学院学报(2022年6期)2022-11-23
动物营养学报(2022年9期)2022-10-12
中国饲料(2022年19期)2022-09-29
动物营养学报(2022年8期)2022-09-01
现代消化及介入诊疗(2022年4期)2022-07-12
世界科学技术-中医药现代化(2021年9期)2021-12-31
中国畜牧杂志(2021年3期)2021-12-05
中国饲料(2021年17期)2021-11-02
医学前沿(2021年18期)2021-04-14
兽医导刊(2020年11期)2020-12-28
