
高分子纤维及其集合体的拓扑结构对细胞行为的调控
2020-12-02 01:18武小童刘来俊李超婧王富军
东华大学学报(自然科学版) 2020年5期
武小童,何 儿,王 璐,刘来俊,李超婧,王富军
(东华大学 a. 纺织学院;b. 纺织面料技术教育部重点实验室,上海 201620)
由于高分子纤维基材料具有较高的强度、柔性、孔隙率以及比表面积等特点,与人体组织的结构和力学性能相似,因此,在植入修复型医疗器械中具有广泛的应用和巨大的发展潜力。目前,高分子纤维基材料已经被应用于手术缝合线[1-3]、伤口敷料[4-6]、疝气修复补片[7-9]与人工心脏瓣膜[10-12]等生物医用材料中。特别是随着再生医学概念的提出与推广,诱导组织再生成为生物医用材料的重要发展方向。高分子纤维基材料能从物理结构上高度模拟细胞外基质[13],从而为细胞的生长繁殖提供有利环境,因此受到广泛关注。目前,利用高分子纤维基材料诱导诸如皮肤[14-16]、血管[17-19]、韧带[20-22]、肌腱[23-25]、膀胱[26-28]、神经[29-31]及软骨[32-34]等组织再生的研究层出不穷。研究发现,通过构建纤维材料拓扑结构可达到提高材料和生物体整合的效果,且该效果可持续作用于材料的整个服役周期[35-40]。因此,研究拓扑结构对细胞行为的调控对于提升材料诱导组织再生的能力具有重要意义。
高分子纤维基材料的拓扑结构包括两大层次:单纤维结构与纤维集合体结构。单纤维结构包括沟槽结构、多孔结构、串晶结构等;纤维集合体的结构参数包括纤维直径、纤维取向、孔径/孔隙率、孔形等,如图1所示。高分子纤维及其集合体的拓扑结构对细胞行为有显著影响。文献[41-42]综述了纤维直径、取向与孔尺寸等结构参数对细胞行为的影响,但未对细胞的各项行为分类进行讨论。Beachley等[43]综述了纤维基支架结构对细胞形态、黏附、增殖和分化等行为的影响,但未对支架的多项结构参数进行分类讨论。目前,鲜有针对单纤维形貌对细胞行为的调控作用的综述文献。
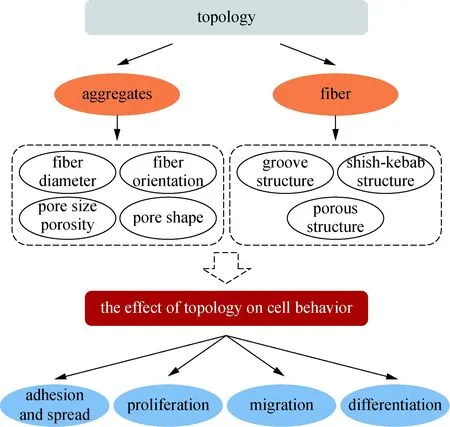
图1 全文架构Fig.1 Article structure
本文从高分子纤维基材料的单纤维及其集合体这两个结构层次出发,较全面地总结评述了拓扑结构对细胞的黏附铺展、增殖、迁移与分化等行为的调控作用,并对纤维材料结构调控细胞行为的知识体系进行梳理,以期为诱导再生材料的设计提供参考。
1 单纤维结构
1.1 沟槽结构
1.1.1 沟槽结构对细胞黏附和铺展的调控
具有适当沟槽结构的纤维或基于此类纤维的支架能够在一定程度上改善细胞的黏附。例如,Sinclair等[44]研究发现,人皮肤成纤维细胞在具有沟槽结构的毛细管通道纤维支架上的黏附优于圆形截面纤维对照组。这是由于表面凹槽形成的二级结构提高了纤维与集合体的比表面积,使得细胞与纤维间的力学相互作用更加紧密。
1.1.2 沟槽结构对细胞增殖的调控
沟槽结构对于细胞增殖具有一定的促进作用,但并非起决定性作用。Park等[45]发现,成骨肉瘤细胞在十字形截面聚己内酯(polycaprolactone, PCL)纤维支架上的增殖量显著高于圆形、三角形截面纤维支架;Williamson等[46]通过湿法纺丝联合冷拉伸工艺制备了表面具有沟槽状纹理的PCL纤维,发现该纤维能够提升成纤维细胞和成肌细胞的增殖率;Lu等[47]发现,表面带有微米级凹槽的纤维因其毛细作用而能够瞬间移动流体,具有向三维支架内部进行物质输送的潜力。对于贴壁细胞,黏附是增殖的先决条件[48],因此沟槽结构可通过促进细胞黏附间接促进增殖;同时,沟槽结构所增大的比表面积为细胞增殖提供了更多的可用表面;此外,特定尺寸的沟槽具有毛细作用,能够显著提升支架的流体输送性能,提高三维支架内部营养物质与氧气的递送效率,从而促进三维支架内部细胞的增殖。
1.1.3 沟槽结构对细胞迁移的调控
沟槽结构能够引导细胞的迁移方向,如图2所示。Huang等[49]发现,沟槽结构能够促进神经细胞的线性生长和细胞骨架的伸长;Lu等[47]将鼠皮肤成纤维细胞和鼠主动脉平滑肌细胞接种至毛细通道纤维支架上,4周后发现细胞在沿凹槽的方向高度对齐并沉积细胞外基质,证实细胞按照凹槽的方向进行迁移;Kang等[50]制备了具有纵向沟槽微结构的扁平带状海藻酸钠纤维,证实沟槽纤维的表面拓扑特征可被成肌细胞和神经细胞感知,并能够指导其迁移。沟槽结构纤维调控细胞迁移的表观因素是高度有序的表面凹槽结构所给予细胞的各向异性力学刺激,内在机制是细胞的机械转导[51],即通过黏着部位感知各向异性力学刺激,并从基因层面调整自身形态特征,沿沟槽方向延伸以适应基质形貌。
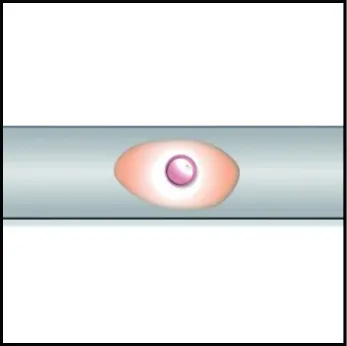
(a) 单根圆形纤维
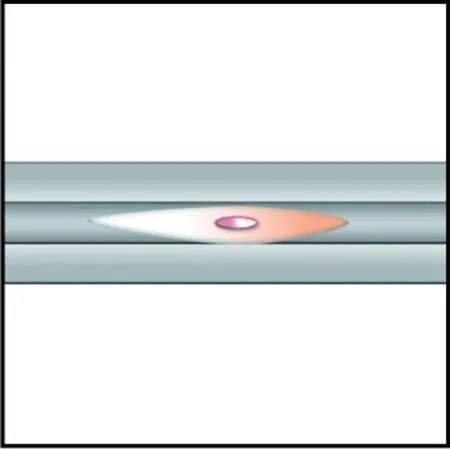
(b) 单根沟槽纤维
沟槽的深度与宽度对细胞迁移有着重要影响,引导不同细胞发生迁移的最佳沟槽宽度并不统一,但基本都集中于微米及亚微米尺度[47, 49, 50, 52]。文献[53]证实,细胞对于沟槽宽度的感知具有一定的阈值,超过阈值范围将失去诱导效果。
此外,Andersson等[54]发现,具有连续规则边缘的凹槽表面诱导细胞取向迁移排列的作用更加明显。
1.1.4 沟槽结构对细胞分化的调控
文献[55-56]证实表面沟槽机制对细胞分化存在调控作用,但这些研究都基于平面基质上的沟槽处,而尚未见到针对纤维表面沟槽结构影响细胞分化的研究报道。通过类比可知:在纤维表面设置类似尺度的沟槽同样能够对细胞分化产生特定作用;而纤维表面的沟槽结构与纤维的直径、排列方式可能具有协同作用。因此,通过基于纤维的多尺度结构设计来实现对细胞分化的调控十分令人期待。
1.2 多孔结构
1.2.1 多孔结构对细胞黏附和铺展的调控
与圆形纤维集合体相比,具有多孔表面结构的纤维集合体能够显著增大细胞的铺展程度,促进细胞的黏附,如图3(a)和3(b)所示。Jiang等[57]发现,成纤维细胞和内皮细胞在多孔PCL纤维支架上的铺展程度显著优于同尺寸圆形纤维支架;Yang等[58]提出细胞的伪足可以锚定在多孔纤维表面的孔隙中,从而改善细胞的黏附作用。多孔纤维有利于细胞黏附,这是因为多孔二级结构所带来的高比表面积与表面粗糙度有利于黏附蛋白的吸附[59],可为细胞提供更多的结合位点,增大细胞的铺展程度,进而促进细胞的黏附。
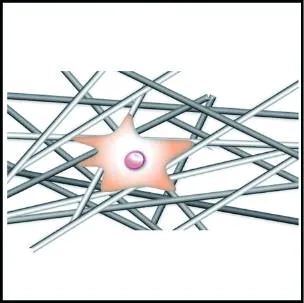
(a) 圆形纤维集合体
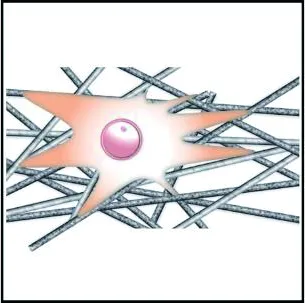
(b) 多孔纤维集合体
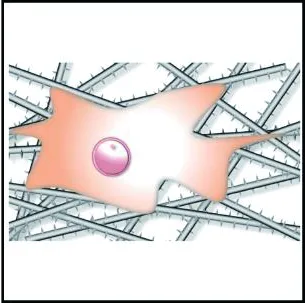
(c) 串晶纤维集合体
1.2.2 多孔结构对细胞增殖的调控
纤维表面的孔结构有利于细胞增殖。Pant等[60]制备了高度多孔PCL纤维,并使用成骨细胞评估基于该纤维制备的支架,发现相比同尺寸的光滑表面纤维基支架,其细胞增殖情况有显著改善;Jiang等[57]研究发现,细胞在多孔纤维基支架上的增殖率明显高于同尺寸的圆形纤维支架。这是由于多孔表面显著提升了纤维及其集合体的表面粗糙度和比表面积,使得细胞黏附作用显著增强,增殖空间扩大,进而促进细胞增殖。
1.2.3 多孔结构对细胞迁移的调控
纤维表面特定尺度的孔结构能够提高细胞与纤维的结合紧密程度,从而加强纤维对细胞定向迁移的引导作用。周祺惠[61]制备了由多孔左旋聚乳酸(poly-L-lactide, PLLA)纤维组成的取向纤维支架,研究发现,随着纤维表面孔径的增大和孔隙率的提高,细胞沿纤维轴向的伸长率与取向程度均显著增大。此外,纤维表面的多孔结构还有利于细胞向支架内部的快速迁移[60]。
1.3 串晶结构
1.3.1 串晶结构对细胞黏附和铺展的调控
部分纤维表面具有的串晶结构有利于细胞黏附和铺展,如图3(c)所示。Jing等[62]将成纤维细胞接种至PCL串晶纳米纤维支架上,发现细胞沿纤维铺展程度高于圆形纤维支架,且可以明显地看到细胞的丝状伪足;Wang等[63]指出串晶纤维的高比表面积使其能够吸附更多的玻连蛋白和纤连蛋白分子,从而进行细胞黏附。
1.3.2 串晶结构对细胞增殖的调控
在纤维表面构建串晶结构,能够显著提升细胞的增殖率。Li等[64]发现串晶PCL纤维支架上的猪髂动脉内皮细胞增殖显著优于圆形PCL纤维支架,并且不同串晶尺寸对于细胞增殖的影响不同,片晶尺寸为100 nm的串晶PCL支架具有最佳的细胞增殖率。
1.3.3 串晶结构对细胞迁移的调控
串晶结构能够促进细胞向支架内部的迁移。Jing等[65]制备了串晶PCL纤维三维支架并对成纤维细胞的迁移情况进行评估,发现与圆形纤维三维支架相比,串晶纤维三维支架上的细胞向支架内部迁移的深度明显增大。
目前鲜有关于串晶对纤维细胞定向迁移的调控作用的文献报道,但通过与多孔纤维类比可知,串晶结构可能会加强细胞与纤维结合的紧密程度,促进纤维对于细胞定向迁移的引导作用。
2 纤维集合体结构
2.1 纤维直径
纤维直径直接影响纤维集合体的比表面积及表面粗糙度,因此其对细胞行为有着重要的调控作用。表1总结了多种不同纤维直径的纤维集合体对不同细胞的作用。
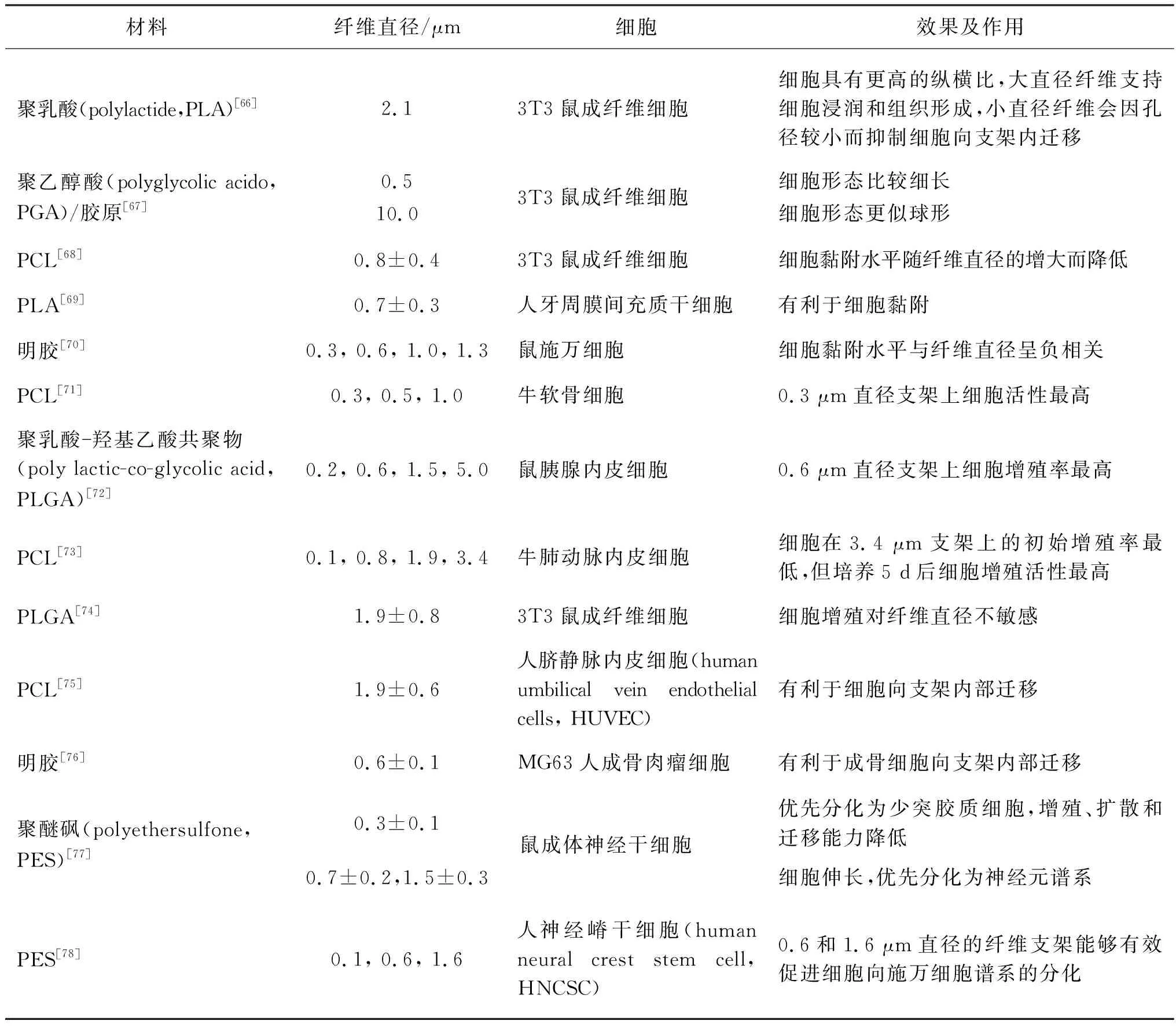
表1 不同纤维直径的纤维集合体对不同细胞的作用
2.1.1 纤维直径对细胞黏附和铺展的调控
许多研究显示直径在亚微米尺度的纤维有利于细胞的黏附。Chen等[68]通过静电纺丝制备了具有不同直径的PCL纤维支架,研究发现,当直径处于纳米尺度(428~1 051 nm)时,细胞黏附速率随纤维直径的增大而降低,而纤维直径处于微米尺度(1 051~1 647 nm)时,细胞黏附速率对直径变化不敏感;Luis等[69]将间充质干细胞接种至直径为0.704~0.880 μm的静电纺丝支架上,发现纤维平均直径为0.704 μm的支架最适宜细胞的黏附;Gnavi等[70]制备了直径分别为300 nm、600 nm、1.0 μm和1.3 μm的明胶纤维支架,发现施万细胞黏附水平与纤维直径呈负相关,纤维直径为300 nm的支架上的黏附最优。纤维直径从微米尺度减小至亚微米尺度时,支架的比表面积明显增加,并且比表面积的增加与纤维排列无关[79],而具有高比表面积的纤维更易吸附黏附蛋白,可为细胞附着提供许多黏着位点。
细胞的铺展状态与所黏附的纤维直径有重要的关联。文献[62, 64]指出:细胞在小于或接近自身尺度的纤维上倾向于包裹缠绕纤维并沿着纤维伸长;细胞在显著大于自身尺度的纤维上呈鹅卵形,类似于在平面基质上的形态,如图4所示。Badami等[66]发现2.1 μm直径的纤维支架上的成纤维细胞比光滑表面上的具有更高的纵横比;Tian等[67]发现,成纤维细胞黏附到较小直径(500 nm)的纤维上时更加细长,黏附到较大直径(10 μm)的纤维上时形态更似球形。细胞包裹小直径纤维的原因是,当细胞与纤维接触时,细胞为保持膜表面的扩散性受体与纤维表面的配体的结合,自发产生细胞膜局部缠绕以减少相互作用的自由能。
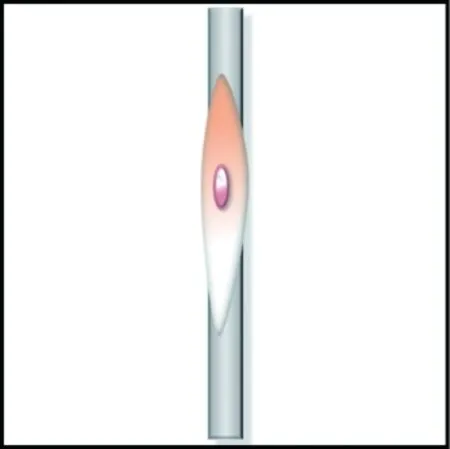
(a) 直径小于细胞尺度的纤维

(b) 直径大于细胞尺度的纤维
2.1.2 纤维直径对细胞增殖的调控
适当的纤维直径能够提高附着其上的细胞的增殖活性与增殖率。多数研究认为,亚微米尺度的纤维有利于细胞增殖。Ko等[72]制备了平均直径分别为200 nm、600 nm、1.5 μm和5.0 μm的PLGA纤维支架,发现内皮细胞在纤维直径为600 nm的支架上增殖率远高于其他组;Noriega等[80]制备了平均直径为300 nm、500 nm和1 μm的壳聚糖纳米纤维支架,发现软骨细胞在纤维直径为300 nm支架上的增殖活性最高。
然而对于亚微米尺度纤维有利于细胞增殖这一观点,有研究人员提出了不同看法。Kim等[73]发现内皮细胞在微米尺度纤维支架上的增殖活性最高;而Bashur等[74]发现,鼠成纤维细胞对PLGA纤维支架的直径0.14~3.60 μm变化不敏感。
2.1.3 纤维直径对细胞迁移的调控
纤维直径能够影响纤维对细胞迁移方向的引导能力。例如,Li等[75]指出,微米级纤维比亚微米级纤维更利于引导细胞的迁移方向,并提出较大尺度的拓扑结构可能具有更强的接触引导效应。
不同细胞的迁移方向对纤维直径的响应有所不同。Fioretta等[71]研究了不同纤维直径的静电纺PCL随机取向支架对内皮细胞和内皮集落形成迁移方向的影响,在直径为5~11 μm的纤维支架上培养时,内皮细胞沿纤维周向排列细胞骨架并沉积成胶原层,而内皮集落形成细胞沿着纤维轴向排列细胞骨架并沉积成胶原层。
Badami等[66]推测,大直径纤维会使细胞浸润并形成组织,而小直径纤维会因孔径较小而抑制细胞向支架内部的迁移。纤维集合体的孔径很大程度上由纤维直径决定[81],直径大的纤维会产生较大的孔,纤维直径较小则支架的孔径往往也较小,使得细胞难以向集合体内部迁移。
2.1.4 纤维直径对细胞分化的调控
纤维直径对干细胞分化表型及分化水平有一定影响。Christopherson等[77]在具有不同纤维直径的电纺PES随机取向纤维支架上培养神经干细胞,发现细胞在纤维直径为283 nm支架上优先分化为少突胶质细胞,在纤维直径为749和1 452 nm支架优先分化为神经元谱系;Ren等[78]制备了直径为160 nm、600 nm与1.6 μm的对齐取向纤维支架,发现直径为600 nm和1.6 μm的纤维支架能够有效促进神经嵴干细胞向施万细胞谱系分化。分化依赖于细胞形态和细胞周期[82],细胞在纤维或支架上的形态与其分化趋势与分化程度密切相关,推测细胞在不同直径纤维上的分化行为很大程度上取决于细胞与纤维尺度的比例。例如,在不同直径的随机取向纤维基质上培养的大鼠成体神经干细胞,在直径远小于自身尺度的纤维基质上不受单根纤维引导,细胞的形态更圆,倾向于胶质细胞表型,而在直径较大的纤维上沿单根纤维延伸,细胞倾向于分化为神经元谱系[77]。
2.2 纤维取向
2.2.1 纤维取向对细胞黏附和铺展的调控
随机取向支架比对齐取向支架更有利于细胞的黏附。Zhong等[83]分别在随机取向与对齐取向的胶原蛋白电纺支架上培养成纤维细胞,发现对齐取向支架上的细胞黏附较差。导致该现象的原因是,对齐支架的表面与内部结构相对规整,而随机支架的不规则表面结构增大了支架的比表面积,并且随机支架内部孔与孔之间高度互连,细胞的丝状伪足能够在随机纤维网络内找到更多的攀附点,使得细胞对随机支架的黏附作用显著优于对齐支架。
细胞在对齐取向的纤维支架上往往沿着纤维的取向伸长,而在随机取向的纤维支架上沿着周围的纤维多极化铺展,如图5所示。Bashur等[74]通过静电纺丝制备了取向度不同的PLGA纤维支架,发现成纤维细胞的长宽比和长轴长度随纤维取向的增加而增加;Whited等[84]将PCL和I型胶原混合静电纺丝形成支架,发现对齐取向支架上的人脐静脉内皮细胞呈细长形,而随机支架上的细胞呈多边形。这是因为对齐取向纤维支架的高度有序的结构给予细胞各向异性的力学刺激,使细胞产生形态变化的响应;随机取向的纤维支架能够给予细胞比较均匀的物理支持,并且表面粗糙度更高,有利于细胞伪足的多向延伸攀附,因此细胞表现为多极铺展形态。
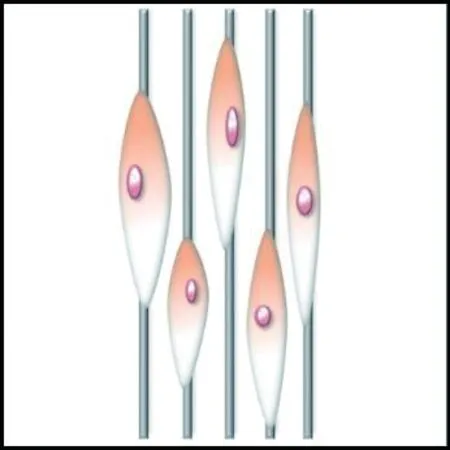
(a) 对齐取向纤维集合体
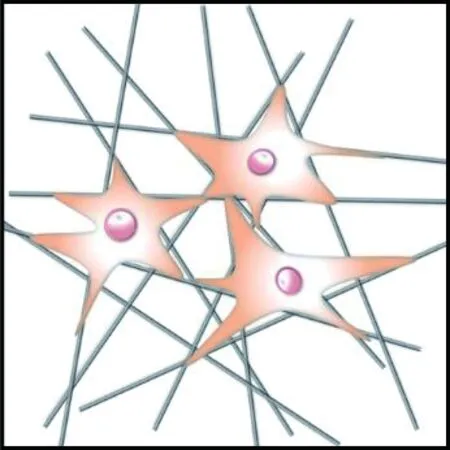
(b) 随机取向纤维集合体
然而上述规律并非适用于所有情况,纤维取向与纤维直径往往共同调控细胞行为,细胞对取向与直径的响应具有更复杂的规律,此外,纤维排列的紧密程度影响细胞行为[79]。
2.2.2 纤维取向对细胞增殖的调控
目前已有许多研究指明了对齐取向或随机取向纤维集合体对细胞增殖的作用,但不同细胞对纤维取向程度的偏好存在很大差异。Subramanian等[85]发现,相比随机纳米纤维支架,施万细胞在对齐取向PLGA-PCL纳米纤维支架上的增殖更好;Shafiee等[86]制备了对齐和随机取向的PLLA/PCL静电纺丝支架,接种细胞后进行培养,发现随机支架上的人鼻中隔源干细胞增殖率较高;Mirzaei等[87]发现,在对齐的碳纳米纤维支架上培养的人子宫内膜干细胞的增殖量显著小于随机纤维支架;也有研究人员提出,成纤维细胞的增殖对纤维取向并不敏感[74, 88]。上述各项研究表明,不同组织细胞对于纤维取向的响应并不统一。促进细胞增殖的关键在于纤维支架所提供的环境与组织细胞所在体内环境的相似程度,当从纤维取向上实现对天然细胞外基质的高度模拟时,可能最有利于细胞的增殖。
2.2.3 纤维取向对细胞迁移的调控
相比随机取向纤维,对齐取向的纤维能够引导细胞的迁移方向,提高细胞的迁移速度。Xu等[19]制备了对齐取向的电纺聚(L-丙交酯-己内酯)(poly(l-lactid-co-ε-caprolactone), P(LLA-CL))的纳米纤维支架,发现平滑肌细胞沿着对齐的纳米纤维附着迁移,细胞骨架蛋白的排布也与纳米纤维的方向平行,证明细胞倾向于沿着纤维取向排列;Bouta等[89]将内皮细胞接种到具有对齐和随机两种取向的纤维支架上,发现细胞沿着对齐的纤维延伸与迁移,而随机纤维对细胞定向迁移有明显的抑制作用。
对于对齐取向纤维,其引导细胞定向迁移的原理与纤维表面沟槽结构对细胞迁移的引导机理类似:对齐取向的支架给予细胞各向异性力学刺激,影响了细胞肌动蛋白网络与黏着斑的分布与接触面积[66],细胞通过机械转导感受力学刺激并从基因层面调整自身形态[51],沿着纤维方向伸长并迁移。此外,细胞所感知到的附着物刚度对细胞迁移的方向有重要影响[90],而对齐纤维的取向方向的有效纤维刚度比正交方向高,这将促使细胞沿纤维取向迁移。对于随机取向纤维,接种在随机纤维支架上的细胞:单个细胞可以在单根纤维上排列和迁移,但当细胞遇到交叉纤维时,其迁移会被阻碍;另一种情况是细胞可以附着在沿不同方向排列的多根纤维上,但不同取向的纤维给予细胞相互冲突的力学刺激可能会抑制细胞的排列和迁移[89]。
纤维取向与纤维直径、排列紧密程度往往共同作用于细胞的迁移。例如,Liu等[91]制备了直径为0.16~8.64 μm的电纺聚甲基丙烯酸甲酯(polymethy1 methacrylate, PMMA)纤维,发现成纤维细胞沿纤维轴取向时,具有直径下限( 0.97 μm),在该值以下未观察到细胞沿纤维轴向排列。Bouta等[89]认为,当纤维尺度较小且排列紧密时,由于细胞所接触的纤维数量增多,细胞更牢固地附着于支架上,这可能使得细胞更难以迁移。
细胞向随机取向纤维集合体内部的迁移优于对齐取向纤维集合体[92]。这可能是由于相比随机取向支架,对齐取向支架内部的孔形状比较规则,并且孔与孔之间相互隔离,使得细胞在支架内部不易攀附与移动。
2.2.4 纤维取向对细胞分化的调控
纤维的取向对于细胞的分化行为有着非常重要的调控作用。Lim等[93]发现,相比随机纤维,对齐纤维上有更高比例的神经干细胞显示出神经元分化的标志物;Kijenska-Gawronska等[94]制备了随机和对齐取向的静电纺丝P(LLA-CL)/胶原蛋白I /胶原蛋白III纳米纤维支架,发现对齐的纳米纤维支架促进神经干细胞的分化;Yin等[95]发现,肌腱干细胞在对齐取向PLA纤维支架上的肌腱特异性基因的表达明显高于随机取向纤维支架,并且随机取向纤维支架能够诱导成骨,而对齐取向纤维支架阻碍了成骨过程。
纤维取向对干细胞分化的影响源于底物选择性[93],例如,对齐的纤维基质不利于少突胶质细胞的附着和持续存活,却有利于神经元谱系细胞的附着和存活,因此细胞表现为向神经元谱系分化。同时,对齐取向纤维的各向异性力学刺激也对细胞的分化起到一定作用,细胞感受力学刺激并调整自身形态,而细胞形态与细胞的分化行为直接相关,故可通过对齐纤维的拓扑结构间接调控细胞的分化。
2.3 孔径及孔隙率
孔径与孔隙率是纤维集合体的重要特征,大量研究指出孔径与孔隙率对细胞行为有着重要的调控作用。表2总结了多种不同孔径与孔隙率的纤维集合体对不同细胞的作用。
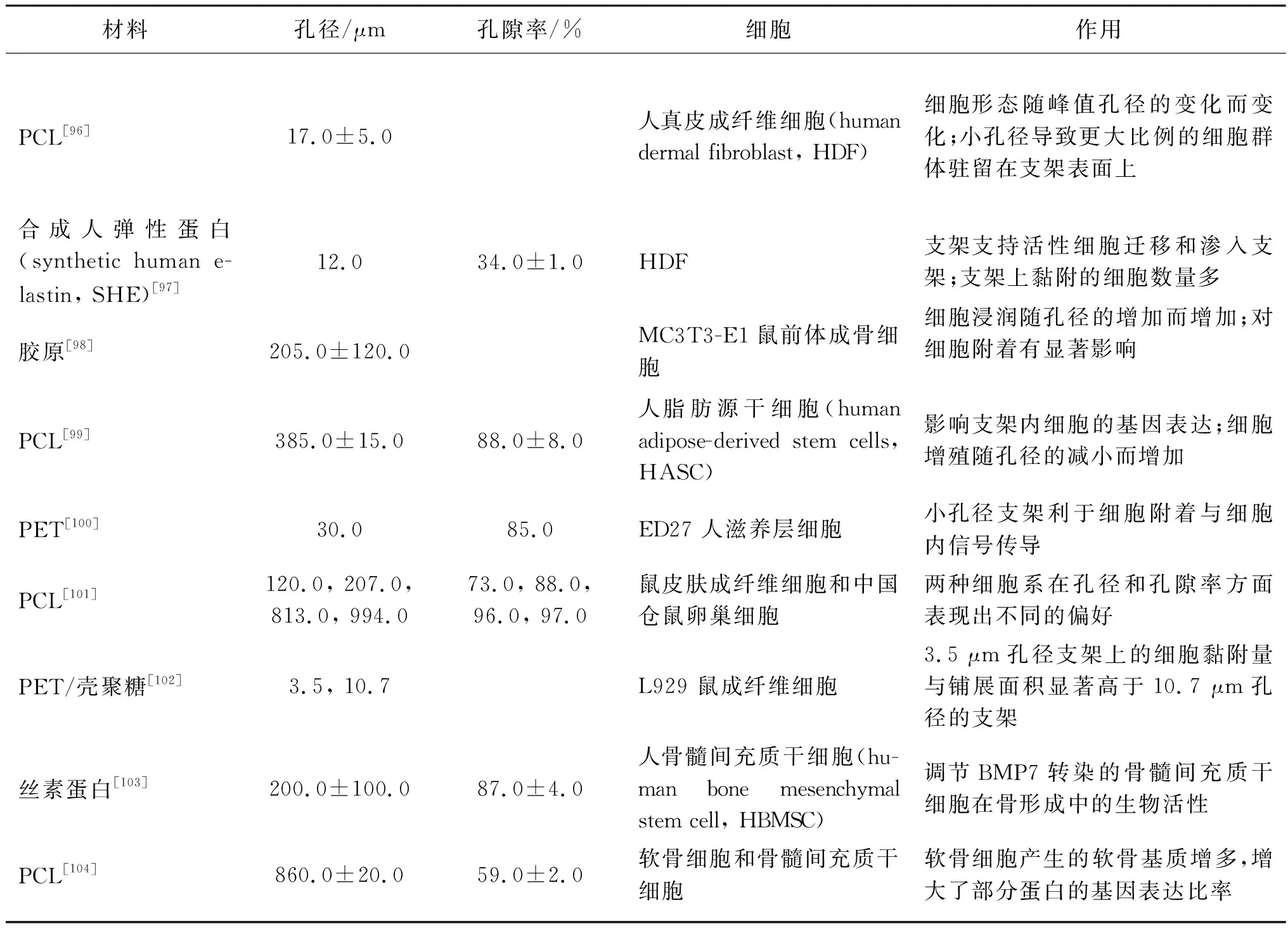
表2 不同孔径与孔隙率的纤维集合体对不同细胞的作用
2.3.1 孔径及孔隙率对细胞黏附和铺展的调控
细胞在孔径小于或接近自身尺寸的纤维集合体上往往呈多极铺展,在孔径显著大于自身尺寸且孔隙率较高的纤维集合体上表现为双极伸长形态。Lowery等[96]制备了孔径为12~23 μm的静电纺丝PCL支架,研究发现,较小孔径支架上的细胞呈多极铺展状态,较大孔径支架上的细胞包裹缠绕在单根纤维上。这是由于在纤维集合体中,孔径与孔隙率的大小与纤维间距密切相关,而细胞能够桥接的纤维间距有限,超过桥接阈值将使得细胞无法同时黏附在多根纤维上,则细胞可用的黏附位点大大减少。因此,过大的孔径与过高的孔隙率不利于细胞的黏附。例如,Veleirinho[102]评估成纤维细胞在平均孔面积分别为9.4和89.3 μm2(孔径分别为3.5和10.7 μm)的静电纺丝聚对苯二甲酸乙二醇酯(polyethylene terephthalate, PET)/壳聚糖支架上的黏附情况,发现细胞在3.5 μm平均孔径的支架上的黏附量与铺展面积均更高。
2.3.2 孔径及孔隙率对细胞增殖的调控
孔径与孔隙率能够调控细胞增殖。Rnjak-Kovacina等[97]将成纤维细胞接种至孔径约为8.0和11.7 μm的支架上并培养8 d,发现孔径较小的支架上的细胞增殖率高于孔径较大的支架;Ma等[100]发现,相比大孔径(35 μm)、高孔隙率(89.6%),小孔径(30 μm)、低孔隙率(84.9%)的PET纤维支架中培养的ED27人滋养层细胞具有较高的初始细胞增殖率和代谢活性。这可能是由于小孔径支架更利于细胞的附着与细胞间信号传导。
研究发现,不同的细胞或组织对孔径的偏好不同。Tang等[101]在研究纤维基质结构与鼠皮肤成纤维细胞和中国仓鼠卵巢细胞增殖关系的过程中发现,虽然两种细胞系在基质中都能正常生长,但它们在孔径和孔隙率方面表现出不同的偏好。因此,在设计组织工程支架时应当考虑种子细胞对孔径及孔隙率的偏好,从而最大程度地诱导组织再生。
2.3.3 孔径及孔隙率对细胞迁移的调控
细胞在小孔径的支架上通常会桥接多根纤维而均匀扩散,而在大孔径的支架上大多沿着单根纤维迁移。Lowery等[96]证实,较大的孔径促使细胞沿着单根纤维排列。Ma等[100]发现,孔径和孔隙率较低的支架上培养的细胞能够更容易地在相邻纤维上扩散。Kennedy等[42]指出,细胞可以跨越较小的孔桥接纤维,而在大于自身尺寸的孔内其会黏附在单根纤维上并沿着单根纤维生长。
细胞向大孔径、高孔隙率纤维集合体内部的迁移程度更高,而小孔径、低孔隙率集合体上的细胞往往驻留在表面。Rnjak-Kovacina等[97]发现,高孔隙率支架有利于细胞向支架内部迁移。Murphy等[98]发现,孔径越大细胞向支架内部迁移程度越高。较大的孔径使细胞向集合体内部迁移的阻碍减少,同时能更高效地实现氧气、营养物质的输送与代谢废物的清除(如图6所示),使得细胞能够快速向内迁移;而小孔径会将细胞屏蔽在支架表面,并且物质交换效率更低,使得细胞难以渗透。因此,Lowery等[96]提出,在组织工程用电纺支架的设计开发过程中,孔隙必须足够大,以促进细胞快速渗透入支架内部,但不能大到阻止细胞快速构建细胞外基质时对纤维的桥接。
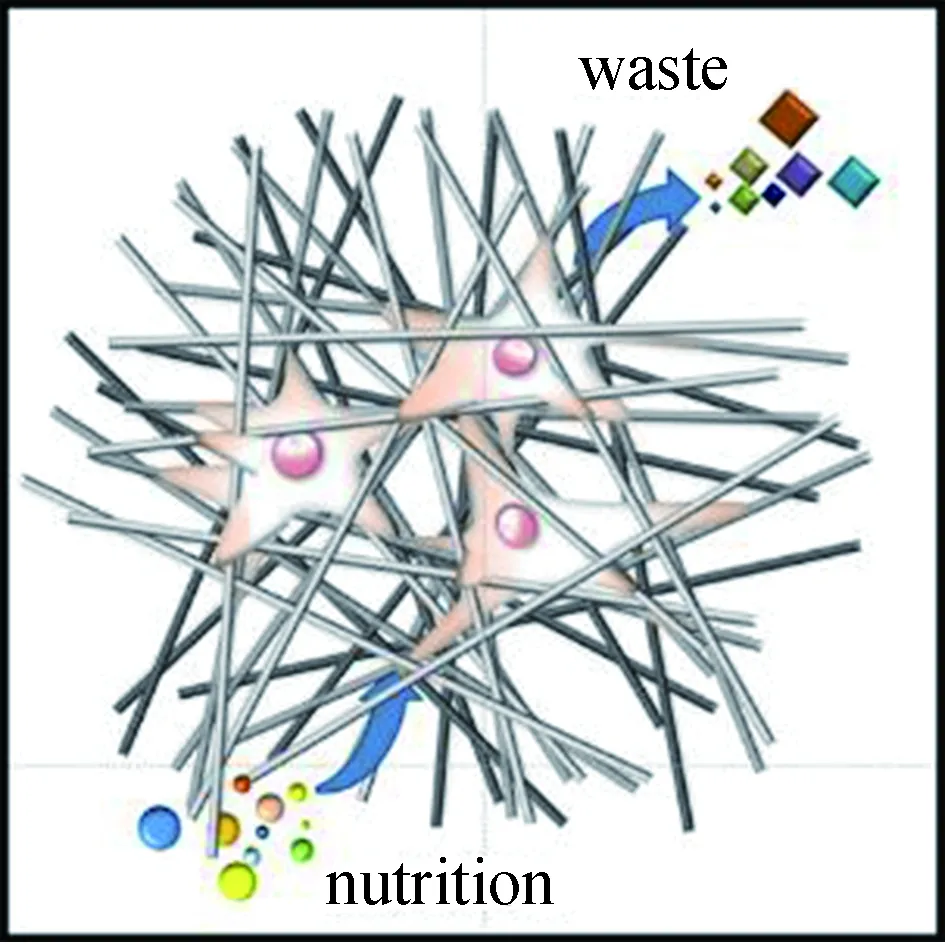
图6 大孔径、高孔隙率的纤维集合体有利于细胞渗透和物质交换Fig.6 Fiber aggregate with large pore size and high porosityfacilitates cell infiltration and material exchange
从另一个角度来看,小孔径材料难以被组织细胞渗透的特点也可加以利用。例如,Sill等[105]提出,具有小孔径的静电纺纳米纤维网的高比表面积能够支持内皮细胞的良好黏附,而其纳米尺度的孔径能够阻止微米尺度的平滑肌细胞向血管腔内的迁移,因此非常适合应用于血管组织工程,如图7所示。

图7 静电纺丝网状纳米纤维集合体用于血管组织工程Fig.7 Reticular electrospun nanofiber aggregate usedfor vascular tissue engineering
2.3.4 孔径及孔隙率对细胞分化的调控
纤维集合体的孔径及孔隙率能够调控细胞的分化方向与分化活力。Kennedy等[42]提出,孔径能够改变细胞的铺展面积,从而影响细胞分化;Kemppainen等[104]发现,通过改变支架孔径与孔隙率而改变支架通透性,能够影响软骨细胞的再分化进程;Zhang等[103]通过冷冻干燥技术制备了孔径为50~300 μm的4种丝素蛋白纤维支架,研究发现,转染了骨形态发生蛋白BMP7的骨髓间充质干细胞在100~300 μm孔径的支架上的成骨分化标志物表达良好,分化活力最高;Oh等[99]在孔径和孔隙率梯度分布(90~400 μm,80%~97%)的PCL纤维支架上接种脂肪干细胞,发现孔径为380~405 μm的支架上细胞的软骨分化标志物表达最高。
2.4 孔形
2.4.1 孔形对细胞黏附和铺展的调控
目前已有研究表明,纤维集合体孔形能够调控细胞黏附铺展的形态。Di Luca等[106]将间充质干细胞接种在孔隙率相同而孔形不同的PCL纤维支架内研究细胞情况,结果显示,不同孔形支架上的细胞黏附形态不同,在孔形较规则的支架上的细胞具有更均匀的形态。在纤维集合体中,孔形与纤维间距密切相关。规则的孔形意味着纤维间距具有更高的各向同性,因此细胞在支架上向四周铺展更加均匀;而不规则的孔形反映纤维间距具有各向异性,细胞能在某一方向上向周围的纤维上铺展,而在另一方向上无法桥接多根纤维,只能在单根纤维上包裹缠绕,且形态不规则。
2.4.2 孔形对细胞增殖的调控
细胞增殖总量对孔形的变化并不敏感。文献[106]指出,支架内的孔形对细胞增殖总数量基本无影响;Rumpler等[107]研究了几何形状(三角形、正方形、六边形和圆形)对三维组织生长的影响,结果发现,MC3T3-E1鼠前体成骨细胞在人造三维基板中形成的新组织的总量与孔形无关,而是取决于孔周长。对于该研究使用的棱柱形孔,其孔周长与孔表面积成正比,由此推测,细胞增殖总量的决定性因素可能是材料所能够提供的表面积。该研究同时发现,细胞在圆形孔道内表面的增殖速率处处相同;在其他孔形中,细胞首先在高曲率的角落区域快速增殖,直至相邻角落的细胞集群相接触而形成统一的界面,再以处处相同的速度继续增殖,如图8所示。这一现象产生的主要原因,可能是孔道内的曲率对细胞骨架产生了复杂的机械作用,进而影响细胞行为。Bidan等[108]提出,细胞在孔道中增殖的曲率驱动行为是为了使新生组织处于更加稳定的状态,最终细胞会趋向于形成一个均匀圆形界面。虽然上述研究并非都基于高分子纤维及其集合体,但通过类比可知,细胞在纤维集合体内的局部增殖速率会受到不同孔形产生的复杂机械力的影响。

(a) 三角形孔

(b) 方形孔

(c) 圆形孔
2.4.3 孔形对细胞分化的调控
目前已有研究证实孔形对细胞分化存在一定的调控作用。Di Luca等[106]分析了间充质干细胞在孔形梯度分布的支架上的分化情况,发现细胞在菱形孔和方形孔区域内的分化趋势存在明显差异。然而,目前纤维集合体孔形对细胞分化的具体影响机理尚不明确,原因可能是纤维集合体的不同孔形给予细胞不同的力学刺激,而细胞通过力学传导通路感受刺激并调节基因表达和干细胞分化[109-110]。
3 结论与展望
本文从单纤维形貌与纤维集合体结构2个层次上阐述了高分子纤维材料拓扑结构对细胞行为的调控作用。基于单纤维的沟槽结构、多孔结构与串晶结构能够显著增大纤维及其集合体的比表面积与表面粗糙度,有利于细胞的黏附与增殖;单纤维沟槽结构与纤维集合体的取向通过给予细胞各向异性力学刺激而改变细胞的铺展形态,可调节细胞分化并引导细胞迁移方向;孔径、孔隙率是细胞向纤维集合体内部迁移的决定性因素,同时也对集合体的物质交换能力有着重要的调控作用;而孔形能够通过局部曲率的变化来调控细胞局部增殖速率。
不同组织细胞对于支架有着不同的结构偏好,当支架各项结构参数与天然细胞外基质相类似时,最适于细胞的生长繁殖。这将给予研究者们一定的启示,即通过支架拓扑结构调控细胞行为的关键在于从孔径、孔隙率、纤维形貌、纤维直径与取向多个角度对细胞外基质进行高度模拟,给予细胞与天然组织类似的微观环境,可促进细胞生长繁殖与组织迅速再生。
在开发用于诱导组织再生的新型高分子纤维基材料时,应当从所要调控的细胞行为出发,并从纤维与集合体两个层次综合设计材料拓扑结构。例如,为促进细胞黏附,选择串晶纤维与较小的孔径;为加强对细胞迁移方向的引导,选择沟槽纤维与对齐的取向。未来应致力于,在不同细胞种类、细胞行为与多种材料及其拓扑结构之间建立一套完整细致的关系系统,这对于组织诱导性新型高分子纤维及其集合体材料的精细化设计与研究有着重要的指导意义。
猜你喜欢
农业工程学报(2022年10期)2022-08-22
煤炭工程(2022年7期)2022-07-21
西北工业大学学报(2022年2期)2022-05-11
数字海洋与水下攻防(2022年2期)2022-04-26
清华金融评论(2022年4期)2022-04-13
建材发展导向(2022年1期)2022-03-08
军事文摘(2021年24期)2022-01-11
航空发动机(2021年2期)2021-05-18
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
