
低磷胁迫对赤皮青冈幼苗生长及生物量的影响
2020-12-07 01:29陈雯彬欧阳泽怡欧阳硕龙李志辉何友军
湖南林业科技 2020年5期
陈雯彬, 欧阳泽怡, 欧阳硕龙, 李志辉, 何友军
(1.湖南省农林工业勘察设计研究总院, 湖南 长沙 410007; 2.中南林业科技大学, 湖南 长沙 410004;3.湖南省林业科学院, 湖南 长沙 410004; 4. 湖南省森林植物园, 湖南 长沙 410116)
赤皮青冈Cyclobalanopsisgilva又名红椆[1],在我国的贵州、湖南、广东等省份都能生长;适应环境的能力较强,在涵养水源、保持水土方面也有较好表现;是兼具园林观赏、生态绿化与工业用材等多功能的珍贵硬木乡土树种[2]。赤皮青冈成片天然林稀少,进行科学繁育扩大人工造林规模以满足市场需求显得十分必要[3]。
我国土壤普遍存在缺磷的状况,特别是南部省份的土壤类型绝大多数为典型缺磷红壤[4],赤皮青冈这一树种在推广栽培过程中必定面临缺磷的问题。针对这一问题,传统应对措施是加大林地养分用量和改善营林方法[5]。然而这些应对措施大多只是从改善林地土壤条件出发,没有深入研究树种个性化的生理特点,所以并没有收到理想效果。
众多研究表明,不同树种的磷素利用效率存在较大差异,并且能够在自身的生理生化过程中表现出来[6-10]。植物生长特征的变化是各植物适应逆境胁迫的最显著表现和重要方式,而生物量在各器官间的分配变化能全面体现植物对各种逆境胁迫的适应能力的强弱。低磷胁迫还造成植株生物量在不同部位的分配发生改变,从而造成植株根冠比发生变化,而植物的生产力与生物量在各部位的分配有密切联系[11-12]。所以,研究低磷胁迫下赤皮青冈生长及生物量响应有非常重要的现实意义。本研究选取1、2年生赤皮青冈幼苗,对其进行模拟低磷胁迫处理,并对各胁迫程度下植株的生长、生物量分配、根冠比等进行测量,探讨1、2年生赤皮青冈对不同浓度低磷胁迫的响应,可为赤皮青冈人工林施肥研究提供参考。
1 材料与方法
1.1 试验材料
以湖南省通道侗族自治县的1、2年生赤皮青冈家系幼苗为试材布置试验。1年生赤皮青冈苗高为(15.0±0.5)cm,地径为(1.9±0.1)cm,2年生苗高为(37.0±0.2)cm,地径为(3.5±0.2)cm。该种源地地理位置109°36′29″E,26°16′45″N。
1.2 试验地概况
试验地点设在湖南省天际岭国家森林公园内,该地位于长沙市区南部,与种源地同属亚热带季风气候。春秋季短,春季气温起伏大,夏季炎热而漫长,冬季结冰少但体感较冷。年平均降水量为1 361.6 mm,年平均温度为17.2 ℃,无霜期275 d[13]。
1.3 试验设计和处理
试验采用水培法,营养液按照霍格兰配方用去离子水配置[14]。试验前,设置5个磷素水平即0.100、0.500、1.000、2.000、5.000 mM(mM=mmol·L-1)的磷浓度试验,结果显示1 mM浓度时植株生长良好,无胁迫响应症状,于是确定该浓度为本研究的对照浓度。本试验设置8个磷素处理,处理1至处理8分别为1.000、0.500、0.100、0.050、0.010、0.005、0.001、0 mM(CK)。每个培养穴内分别放置1株1年生或2年生赤皮青冈幼苗,每个磷素水平3株,重复3次,共144株,其中1、2年生苗各72株。将KH2PO4配置成不同磷浓度水平的溶液,在改变磷浓度时用KCl来调节营养液中K+浓度的平衡,其他营养元素浓度保持不变。在胁迫试验过程中保证全天候不间断通气,及时使用0.1 mol·L-1的HCl溶液和0.1 mol·L-1的NaOH溶液调整营养液的pH值至5.8。为确保营养充足,每3 d要倒掉使用过的营养液,注入新配置的营养液,每30 d测定一次相关指标。
2017年5—10月,在湖南省天际岭国家森林公园的玻璃温室内进行水培试验。5月初,在试验苗圃内将赤皮青冈进行移植。选择大小分别基本一致的1、2年生赤皮青冈幼苗,先用自来水再用重蒸水将幼苗根系轻柔地冲洗干净,并对幼苗进行一定的修剪,然后移栽到黑色塑料培养槽里。培养槽内需提前注入正常培养液,培养1周后进行低磷胁迫处理。6月初(30 d后)开始测定相关指标,每30 d测量一次。9月底完成试验。
1.4 试验指标测定方法
1.4.1 株高和地径 用皮尺测量赤皮青冈幼苗株高,用游标卡尺测量地径。株高和地径均以cm为单位,保留两位小数。低磷胁迫第1天和第120天(试验结束后)分别测量1、2年生赤皮青冈幼苗的株高和地径。第1天的数据为初始值,第120天的数据是最终值,计算见式(1)、式(2)。
株高生长量=最终株高-株高初始值
(1)
地径生长量=最终地径-地径初始值
(2)
1.4.2 生物量和根冠比 低磷胁迫前,选取与试验幼苗大小基本一致的1、2年生赤皮青冈幼苗各9株,用纯化水或重蒸水把根部洗净。晾干后用枝剪将幼苗根、茎、叶分开,放进纱布袋置于烘箱中,保持105 ℃运转30 min后,再调节至75 ℃,持续24 h。然后每2 h称量干重一次,直到重量不再变轻。用千分位实验天平称量,保留三位小数,取其平均值作为根、茎、叶的生物量初始值。胁迫120 d后,将最后剩下的植株,照前述的同样方法晾干,将根、茎、叶分开,烘干水分,称量干重,计算见式(3)、式(4)、式(5)和式(6)。
地上部分生物量=最终叶生物量+最终枝生物量
(3)
生物量增量=最终生物量-生物量初始值
(4)
生物量积累=根部生物量增量+地上部分生物量增量
(5)
根冠比=地下部分干重÷地上部分干重
(6)
1.5 数据统计与分析
运用Microsoft Excel 2003软件进行数据统计和作图,采用SPSS 20.0进行数据分析,运用单因素方差分析方法(ANOVA)进行方差分析,运用邓肯法进行多重比较。
2 结果与分析
2.1 生长对低磷胁迫的响应
从表1可以看出,随着磷元素浓度降低,1、2年生赤皮青冈幼苗株高生长量也随之减少。除了处理2外,1、2年生赤皮青冈幼苗株高生长量和地径生长量与对照组都存在显著差异(P<0.05)。与对照相比,处理3、处理4、处理5、处理6和处理7的1年生苗株高生长量分别降低了15.32%、19.79%、33.86%、36.55和38.89%,2年生株高生长量分别降低了12.82%、16.67%、25.53%、28.50%和30.25%。在处理8的磷浓度下株高生长量降低幅度最大,植株生长缓慢,1年生赤皮青冈幼苗株高生长量减少了39.98%,2年生赤皮青冈幼苗株高生长量减少了31.32%,与对照组存在显著差异(P<0.05)。

表1 低磷胁迫下1、2年生赤皮青冈幼苗株高生长量Tab.1 Height increment of annual and biennial seedlings of Cyclobalanopsis gilva under low phosphorus stress处理磷浓度/ mM 株高生长量/cm1年生2年生11.00020.11±0.68 a25.19±0.68 a20.50019.42±0.72 a24.16±0.72 a30.10017.03±0.62 b21.96±0.81 b40.05016.13±0.51 b20.99±0.82 b50.01013.30±0.43 c18.76±0.71 c60.00512.76±0.36 cd18.01±0.70 cd70.00112.29±0.40 d17.57±0.69 cd8012.07±0.39 d17.30±0.71 d 注:表中数据为平均值±标准差;表中同列不同小写字母表示处理之间差异显著(P<0.05)。下同。
从表2可以看出,随着磷元素浓度降低,1、2年生赤皮青冈幼苗的地径生长量都随之减少。除了处理2外,其他浓度下1、2年生赤皮青冈幼苗的地径生长量与对照组都存在显著差异(P<0.05)。与对照相比,处理3、处理4、处理5、处理6和处理7的1年生苗地径生长量分别减少了13.25%、16.57%、28.61%、30.72%和32.23%,2年生苗地径生长量分别减少了10.51%、14.87%、23.85%、26.67%和28.46%。在处理8的磷浓度下地径生长量降低程度最大,与对照组相比较,1年生赤皮青冈幼苗地径生长量减少了34.04%,2年生赤皮青冈幼苗地径生长量减少了29.49%。

表2 低磷胁迫下1、2年生赤皮青冈幼苗地径生长量Tab.2 DBH increment of annual and biennial seedlings of Cy-clobalanopsis gilva under low phosphorus stress处理磷浓度/ mM 地径生长量/cm1年生2年生11.0000.33±0.02 a0.39±0.02 a20.5000.32±0.02 a0.38±0.02 a30.1000.29±0.02 b0.35±0.01 b40.0500.28±0.02 b0.33±0.01 b50.0100.24±0.01 c0.30±0.01 c60.0050.23±0.01 c0.29±0.01 c70.0010.23±0.01 c0.28±0.01 c800.22±0.01 c0.28±0.01 c
2.2 生物量分配对低磷胁迫的响应
从表3可以看出,在低磷胁迫试验中,1、2年生赤皮青冈幼苗的根部生物量增量和地上部分生物量增量随低磷胁迫程度增加而减少。在处理2、处理3和处理4的磷浓度下,与对照组相比,1、2年生赤皮青冈幼苗根部生物量增量差异不显著,根部的生物量增量的减少程度不大;在处理5、处理6、处理7和处理8的磷浓度下,与对照组相比,1、2年生赤皮青冈幼苗根部生物量增量存在显著差异(P<0.05),根部生物量增量的减少较明显,在处理8的磷浓度下,根部生物量增量减少幅度达到最大,与对照组相比较,1年生赤皮青冈幼苗根部生物量增量减少了21.09%,2年生赤皮青冈幼苗根部生物量增量减少了20.68%。在处理2的磷浓度下,1、2年生赤皮青冈幼苗地上部分生物量增量与对照组差异不显著,地上部分生物量增量减少幅度较小,地上部分生物量增量变化不大;在处理3、处理4、处理5、处理6、处理7和处理8的磷浓度下,1、2年生赤皮青冈幼苗地上部分生物量增量与对照组存在显著差异(P<0.05),地上部分生物量增量减少幅度增大;在0 mM的磷浓度下,地上部分生物量增量减少的程度达到顶峰,与对照组差异显著(P<0.05),1年生赤皮青冈幼苗地上部分生物量增量减少了80.91%,2年生赤皮青冈幼苗地上部分生物量增量减少了79.23%。
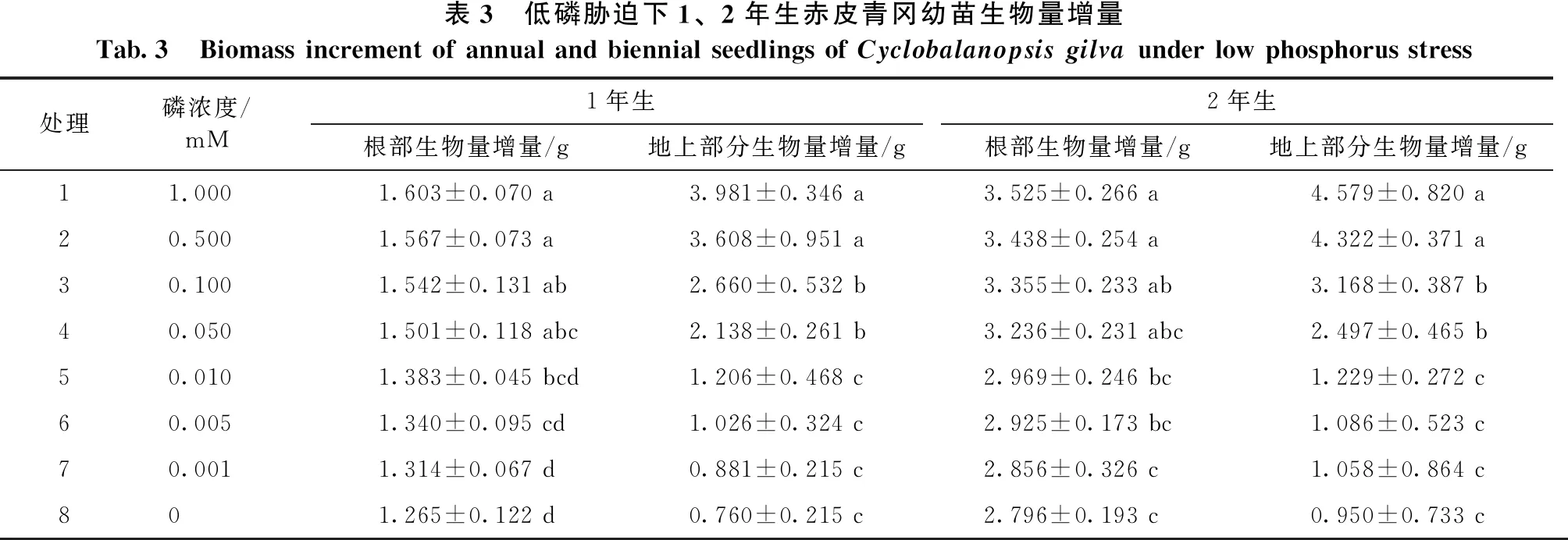
表3 低磷胁迫下1、2年生赤皮青冈幼苗生物量增量Tab.3 Biomass increment of annual and biennial seedlings of Cyclobalanopsis gilva under low phosphorus stress处理磷浓度/ mM 1年生2年生根部生物量增量/g地上部分生物量增量/g根部生物量增量/g地上部分生物量增量/g11.0001.603±0.070 a3.981±0.346 a3.525±0.266 a4.579±0.820 a20.5001.567±0.073 a3.608±0.951 a3.438±0.254 a4.322±0.371 a30.1001.542±0.131 ab2.660±0.532 b3.355±0.233 ab3.168±0.387 b40.0501.501±0.118 abc2.138±0.261 b3.236±0.231 abc2.497±0.465 b50.0101.383±0.045 bcd1.206±0.468 c2.969±0.246 bc1.229±0.272 c60.0051.340±0.095 cd1.026±0.324 c2.925±0.173 bc1.086±0.523 c70.0011.314±0.067 d0.881±0.215 c2.856±0.326 c1.058±0.864 c801.265±0.122 d0.760±0.215 c2.796±0.193 c0.950±0.733 c
2.3 总生物量累积对低磷胁迫的响应
从表4可以看出,在低磷胁迫试验中,1、2年生赤皮青冈幼苗的总生物量累积随低磷胁迫程度增加而减少。1、2年生赤皮青冈幼苗在处理2的磷浓度下,总生物量累积降幅比较小,与对照组差异不显著;在处理3、处理4、处理5、处理6和处理7的磷浓度下,总生物量累积明显降低,1年生赤皮青冈幼苗总生物量累积分别比对照组减少了24.74%、34.84%、53.63%、57.62%和60.68%。2年生赤皮青冈幼苗总生物量与对照组存在显著差异(P<0.05),分别比对照组减少了19.51%、29.26%、46.87%、50.51%和51.70%;在处理8的磷浓度下总生物量累积降低幅度最大,与对照组存在显著差异(P<0.05),1年生赤皮青冈幼苗总生物量累积比对照组减少了63.73%,2年生赤皮青冈幼苗总生物量累积减少了55.01%。
2.4 根冠比对低磷胁迫的响应
从表5可以看出,在低磷胁迫试验中,1、2年生赤皮青冈幼苗根冠比随低磷胁迫程度增加而增加。在处理2、处理3的磷浓度下,1年生赤皮青冈幼苗根冠比与对照组差异不显著;在处理2、处理3和处理4的磷浓度下,2年生赤皮青冈幼苗根冠比与对照组差异不显著;在处理4、处理5、处理6、处理7和处理8的磷浓度下,1年生赤皮青冈幼苗根冠比与对照组存在显著差异(P<0.05);在处理5、处理6、处理7和处理8的磷浓度下,2年生赤皮青冈幼苗根冠比与对照组存在显著差异(P<0.05);在0 mM的磷浓度下,1、2年生赤皮青冈幼苗根冠比最大,1年生赤皮青冈幼苗根冠比相比于对照组增加了54.55%,2年生赤皮青冈幼苗根冠比相比于对照组增加了57.14%。

表4 低磷胁迫下1、2年生赤皮青冈幼苗总生物量累积Tab.4 Total accumulation of biomass of annual and biennial seedlings of Cyclobalanopsis gilva under low phos-phorus stress处理磷浓度/ mM 总生物量累积/g1年生2年生11.0005.583±0.411 a8.104±1.583 a20.5005.176±1.002 ab7.760±0.778 ab30.1004.202±0.664 bc6.523±0.612 bc40.0503.638±0.372 c5.733±1.684 c50.0102.589±0.501 d4.306±1.511 d60.0052.366±0.415 d4.011±0.685 d70.0012.195±0.277 d3.914±1.183 d802.025±0.600 d3.646±0.922 d

表5 低磷胁迫下1、2年生赤皮青冈幼苗根冠比Tab.5 Root shoot ratio of annual and biennial seedlings of Cyclobalanopsis gilva under low phosphorus stress处理磷浓度/ mM 根冠比1年生2年生11.0000.44±0.06 c0.49±0.01 c20.5000.47±0.07 c0.52±0.09 c30.1000.51±0.08 c0.57±0.14 b40.0500.55±0.11 b0.61±0.06 b50.0100.64±0.17 b0.72±0.11 a60.0050.65±0.04 ab0.74±0.12 a70.0010.67±0.12 ab0.76±0.08 a800.68±0.07 a0.77±0.04 a
3 讨论
3.1 生长对低磷胁迫的适应
在自然界中,植物生长特征的变化是各植物适应逆境胁迫的最显著表现和重要方式,生长特征的变化带来生物量在各器官间的分配变化[15]。这是植物在逆境胁迫下自身适应能力的体现。所以,植物的株高、地径、生物量和根冠比是评价植物抗逆性的主要方面,在耐低磷品种选育方面存在非常重要的意义[16]。通过本研究发现,在低磷胁迫试验中,1mM的磷浓度下(处理1),1、2年生赤皮青冈幼苗的株高生长量和地径生长量最大,随着低磷胁迫的程度加大,1、2年生赤皮青冈幼苗的株高生长量和地径生长量都有不同幅度的减小,可以看出赤皮青冈通过调整生长速率来适应各种程度的低磷胁迫。而且不同苗龄赤皮青冈的生长变化存在一定的差异,2年生赤皮青冈株高生长量和地径生长量明显比1年生赤皮青冈幼苗降速要缓慢,在一定程度上可以说明在同等低磷处理下,2年生赤皮青冈幼苗生长比1年生赤皮青冈幼苗受低磷影响较小。
3.2 生物量累积、分配和根冠比对低磷胁迫的适应
低磷胁迫使植物的生长速度滞后迟缓,植株体型孱羸,导致被胁迫植物的生物量累积较正常植物降低[16]。低磷胁迫还造成植株生物量在不同部位的分配发生改变,从而造成植株根冠比发生变化,而植物的生产力与生物量在各部位的分配具有密切联系[17]。在本次试验中,随着磷胁迫的程度加深,所有幼苗不管是根部还是地上部分的生物量增量都表现出一定程度的降低,总生物量增量亦然。根冠比也随着磷元素浓度的降低而变大,根部生物量增量与地上部分生物量增量相比,根部生物量增量减少的程度较小,表明随着低磷胁迫程度增强,赤皮青冈幼苗缺磷程度加大,为满足幼苗对磷元素的需求,苗高的生长速度放缓,地径不再以之前的速度增大,而根部的生长加快,这样既能减少自身对磷的消耗,又能让根系获得更强大的吸收磷元素的能力,更加适应低磷环境。
植物的根部是最先受到胁迫影响的器官[18],根冠比能反映出植株根部与地上部分协调的状态[19]。梁秀兰等[20-23]研究证实,光合作用产物转移到地下部分的生物量增加,根部生物量增量占总生物量增量比例增大,这一表现是植株自觉适应环境的一种反应机制[24-25]。
在本次低磷胁迫试验中,1、2年生赤皮青冈幼苗根冠比变大,且随着磷浓度的降低持续变大,由此可知,低磷胁迫的程度与光合产物的分配相关。低磷胁迫对赤皮青冈地上部分生长的抑制相对大于其对根部生长的抑制,分配到根部的比例相对于地上部分较大。
4 结论
低磷胁迫下赤皮青冈幼苗可以通过调节自身生长和生物量的分配来适应低磷胁迫[26]。试验结果显示,随着低磷胁迫程度的加强,赤皮青冈幼苗的株高及地径受抑制加重,赤皮青冈幼苗的总生物量累积减少,根冠比增加,在0.01 mM、0.005 mM、0.001 mM、0 mM的磷浓度下,与对照组相比,1、2年生赤皮青冈幼苗生长量、根部生物量增量、总生物量累积和根冠比均存在显著差异(P<0.05);赤皮青冈幼苗通过降低自身生长和生物量增量来减少对磷元素的消耗,而且会将有限的资源优先分配给根系,从而充分吸收有限的磷元素来维持自身的地上部分的生长。
猜你喜欢
江西农业(2022年8期)2022-05-13
草业科学(2022年2期)2022-03-21
绿色科技(2021年21期)2021-11-26
耕作与栽培(2021年6期)2021-02-13
防护林科技(2020年12期)2020-03-08
中国科技纵横(2018年14期)2018-08-30
现代农业科技(2017年16期)2017-09-22
报刊荟萃(上)(2017年7期)2017-07-16
魅力中国(2016年29期)2017-05-27
江苏农业科学(2016年6期)2016-07-25
