
硬骨鱼类血红蛋白转换表达的新模式
——以黄河裸裂尻鱼胚胎型血红蛋白研究为例
2020-12-10 03:55陈祺昌郑志琴王发艳祁得林
水生生物学报 2020年6期
陈祺昌 郑志琴 刘 丹 王发艳 晁 燕 章 昭 祁得林
(1. 青海大学省部共建三江源生态与高原农牧业国家重点实验室, 西宁 810016; 2. 青海大学生态环境工程学院, 西宁 810016;3. 青海大学农牧学院动物科学系, 西宁 810016)
血红蛋白(Hemoglobin, Hb)占脊椎动物红细胞胞浆总蛋白的98%以上, 是机体内运输氧的特殊蛋白[1]。在大多数脊椎动物中, 血红蛋白是由2条α-珠蛋白链和2条β-珠蛋白链组成的异源四聚体(α2β2),每条多肽链均与在氧气传输中起直接作用的血红素基团配位。研究表明, 脊椎动物拥有多个珠蛋白编码基因, 在个体发育的不同阶段所表达的珠蛋白编码基因有所不同[1—3]。
在人类(Homo sapiens)基因组中, 胚胎型(Embryonic)、胎儿型(Fetal)和成年型(Adult)珠蛋白多肽链由两大类基因簇(Gene cluster)指导合成[1,3,4]:一类是位于16号染色体的α珠蛋白基因簇, 编码胚胎型HBZ、成年型HBA1和HBA2珠蛋白多肽链; 另一类是位于11号染色体的β珠蛋白基因簇, 编码胚胎型HBE1、HBG2和HBG1, 成年型HBD和HBB珠蛋白多肽链。在斑马鱼(Danio rerio)基因组中, 胚胎型(Embryonic)、仔鱼型(Larval)和成年型(Adult)珠蛋白编码基因分属主要(Major globin cluster)和次要珠蛋白(Minor globin cluster)两大类基因簇, 主要珠蛋白基因簇位于3号染色体, 而次要珠蛋白基因簇位于12号染色体[2,5]。3号染色体的主要基因簇编码7个珠蛋白多肽链, 其中成年型成对珠蛋白基因hbba1/hbaa1和hbba2/hbaa2以头对头(3′-5′—5′-3′)的转录方式串联在一起共享相同的启动子和增强子[5—7], 或以尾对尾(5′-3′—3′-5′)的转录方式串联在一起分别拥有独立的启动子[2], 同时在主要基因簇中还包括一个位点控制域LCR; 胚胎/仔鱼型成对珠蛋白基因hbbe1/hbae1以头对头的转录方式串联在一起共享相同的启动子和增强子[5]。12号染色体的次要基因簇编码4个珠蛋白多肽链, 均为胚胎/仔鱼型, 其中成对珠蛋白基因hbbe2/hbae5以头对头的转录方式串联在一起, 而hbbe3/hbae4以尾对尾的转录方式串联在一起[3]。
随着个体发育血红蛋白的珠蛋白多肽链组成会发生截然不同的变化, 这种特定阶段表达特殊血红蛋白的现象称为血红蛋白的时序转换(Hemoglobin switching)[1—4,8]。血红蛋白的时序转换表达是脊椎动物的一个相对保守的过程, 脊椎动物血红蛋白多肽链组分的改变是对机体发育过程中需氧量变化的响应[3,5,8,9]。在人类个体发生的初始阶段, 胚胎型血红蛋白(HbE)由2条ε-珠蛋白链和2条ζ-珠蛋白链组成(ε2ζ2), 主要在卵黄囊中的红系祖细胞(Red blood cell progenitors)和幼红细胞(Erythroblast)中合成[1,3,9,10], 随后珠蛋白基因簇上的基因表达发生两次重要的转换。第一次由胚胎型到胎儿型的血红蛋白转换发生在受孕后12周, 其显著特征是胚胎型血红蛋白HbE的合成急剧下调, 直至由胎儿型血红蛋白HbF(α2γ2)所取代[3,11]; 第二次由胎儿型到成年型的血红蛋白转换发生在胎儿出生后6周左右, 其特征是γ-珠蛋白基因表达下调并由成年型血红蛋白HbA(α2β2)所取代[12,13]。斑马鱼在其胚胎发育过程中, 血红蛋白也经历了两次重要的时序转换表达。第一次由胚胎型到仔鱼型血红蛋白转换发生在受精后24—36h, 其显著特征是hbbe3珠蛋白表达急剧下降而hbbe2珠蛋白开始表达, 仔鱼期hbbe2珠蛋白表达显著上升并伴随hbbe5珠蛋白的表达; 第二次由胚胎/仔鱼型到成年型的血红蛋白转换发生在受精后22d, 其显著特征是胚胎型珠蛋白hbbe1/hbbe3和仔鱼型珠蛋白hbbe2表达持续下降, 而成年型主要珠蛋白hbaa2和hbba1开始表达, 直到受精后32d完全建立成年型血红蛋白体系[3]。
鱼类作为连接着脊椎动物和无脊椎动物的典型代表, 在进化史中占有重要地位。但是, 以往有关于鱼类胚胎型血红蛋白的研究主要围绕模式动物斑马鱼展开。截至目前, 还未见有关裂腹鱼亚科鱼类胚胎型血红蛋白的研究报道。因此, 本文以黄河裸裂尻鱼(Schizopygopsis pylzovi)为研究对象, 开展胚胎型血红蛋白组成、基因结构及其发育早期的表达研究, 为深入探讨鱼类血红蛋白的时序转换表达和环境适应机制奠定基础。
1 材料与方法
1.1 实验用鱼
实验中黄河裸裂尻鱼及其不同发育时段胚胎皆采自于青海省循化县苏只黄河鱼类增殖站, 实验用黄河裸裂尻鱼为人工繁殖个体, 平均体重300 g,体长20—25 cm。
1.2 实验试剂与仪器
基因组DNA提取试剂盒购自天根生化科技(北京)有限公司。总RNA提取试剂(RNAiso Plus), 逆转录试剂盒Prime ScriptTMRT Master Mix(Perfect Real Time)、SYBR®Premix ExTaqⅡTM等购自TaKaRa生物(大连)有限公司; 整胚原位杂交用主要试剂DIG-Nick-Translation-Mix、NTP-DIG-RNA labeling Polymerase SP6 and T7均购自Roch; 荧光定量PCR仪(Roch LightCycler480Ⅱ)、微量核酸蛋白测定仪(Implen NP80 Mobile)、体式显微镜(尼康SMZ18)。
1.3 黄河裸裂尻鱼胚胎样品及组织样品
在黄河裸裂尻鱼人工繁殖过程中, 首先对亲鱼进行称重, 根据其体重提前1周注射促黄体素释放激素(LHRH-A2); 然后雌雄鱼分开养殖1周左右以促进其性成熟, 按照雌雄比为3﹕1进行人工受精。受精后脱粘、清除死卵, 0.5h后将受精卵转移至孵化器进行孵化, 受精卵孵化温度为(13.5±0.5)℃。
采集受精后2 hpf (hours post fertilization, hpf)、4 hpf、6 hpf、8 hpf、10 hpf、12 hpf、18 hpf、24 hpf、48 hpf、72 hpf、96 hpf、120 hpf、144 hpf、168 hpf、192 hpf、216 hpf的受精卵, 以及破膜后4h、8h、12h、18h、1d、2d、3d、4d、5d、6d、7d、8d、9d(即220 hpf、224 hpf、228 hpf、234 hpf、240 hpf、264 hpf、288 hpf、312 hpf、336 hpf、360 hpf、384 hpf、408 hpf、432 hpf)的胚胎各两份。用4%的多聚甲醛处理固定其中一份, 备用于原位杂交; 另一份液氮保存备用。另外, 成体黄河裸裂尻鱼麻醉后, 采集肾脏组织液氮保存备用。
1.4 胚胎型血红蛋白基因CDS序列获取
以NCBI中已公布的斑马鱼胚胎型α珠蛋白基因(hbae1、hbae3、hbae4和hbae5)和β珠蛋白基因(hbbe1、hbbe2和hbbe3)CDS序列为搜索条件(Queries), 通过NCBI-Blast-2.2.28 +软件(http://www.ncbi.nlm.nih.gov/)中的Blastn命令搜索实验室前期构建的黄河裸裂尻鱼转录组本地数据库, 获取黄河裸裂尻鱼相应的胚胎型血红蛋白基因CDS序列。为了提高搜索的严格性和准确性, 在Blastn搜索中将E值设置为≤0.001。
1.5 内含子确定及扩增
根据已获得的胚胎型α和β珠蛋白基因序列与斑马鱼基因组序列比对, 初步确定黄河裸裂尻鱼血红蛋白基因内含子和外显子组成。然后在第1外显子和第3外显子区域分别设计各基因的内含子扩增引物(表 1), 以黄河裸裂尻鱼肾组织基因组DNA为模板克隆内含子序列。PCR扩增程序: 95℃预变性3min; 95℃变性30s, 55℃退火30s, 72℃延伸1min,循环35 次; 72℃续延伸5min。PCR扩增产物经1.0%的琼脂糖凝胶电泳后, 进行回收纯化, 然后连接转化以进行测序。
1.6 胚胎型血红蛋白基因串联方式及间隔序列确定
参照斑马鱼基因组序列中hbbe1/hbae1和hbbe3/hbae4“头对头”或“尾对尾”的串联特点, 在黄河裸裂尻鱼成对基因近端外显子序列设计组合引物, 以基因组DNA 为模板, PCR扩增和测序间隔序列, PCR扩增程序和测序方法同上。获得序列经Blast比对验证, 最终确定黄河裸裂尻鱼胚胎型α和β珠蛋白基因的串联方式(表 1)。
1.7 胚胎型血红蛋白基因生物信息学分析
利用Lasergene7.0软件, 对基因序列进行阅读框分析及氨基酸序列推定; Blastx程序(http://blast.ncbi.nlm.nih.gov/Blast.cgi)进行蛋白相似性搜索; 利用DNA MAN 6.0进行氨基酸序列同源性的比较, 利用利用MEGA6.0软件构建邻接(Neighbor-joining,NJ)系统发育树[14]。
1.8 RNA探针制备及整胚原位杂交
通过对已获得的4个胚胎型血红蛋白基因序列设计特异性探针引物(表 1), 并对目的基因进行扩增, 将目的片段插入pGEM-T Easy 载体, 阳性克隆送生工生物测序。测序完成后, 碱裂解法提取质粒,提取的质粒总含量要确保达到10 μg。对测序结果进行比对, 根据比对结果中正向引物的连接方向选择相应的内切酶(单酶切)。若正向引物在T7端, 则用SacⅡ进行酶切; 若正向引物在SP6端, 则用SpeⅠ进行酶切。DNA preparation反应体系如下:质粒DNA10 μg、10×Buffer Trager(add BSA)10 μL、Enzyme(SpeⅠ orSacⅡ)5 μL, 最后用RNase-free H2O 补充至100 μL。37℃孵育2h, 酶切产物经1.0%的琼脂糖凝胶电泳检测后测定浓度以用于RNA探针制备。RNA 探针合成: Digested DNA 1 μg, 10×Transcription buffer 2, NTP-DIG-RNA labeling 2 μL,RNase inhibitor 1 μL, Polymerase SP6 or T7 2 μL,ddH2O 10.5 μL。离心5—10s, 封口膜封口37℃水浴3h之后, 加入2 μL DNaseⅠ(RNase-free), 混匀后37℃孵育15min。加入5 μL 4 mol/L LiCl (RNasefree)和37 μL 100%的无水乙醇(RNase-free), –20℃沉淀过夜。4℃, 12000 r/min, 离心30min, 弃上清。加入70%的乙醇(RNase-free)1 mL洗涤沉淀, 来回颠倒EP 管使沉淀悬浮, 4℃, 12000 r/min, 离心10min,风干沉淀。用50 μL提前预热至55℃的DEPC H2O溶解沉淀,即为标记好的RNA探针, 紫外分光光度计测定浓度, 1%琼脂糖凝胶电泳鉴定RNA探针的质量, 分装并于–80℃保存。
整胚原位杂交步骤详见文献[15, 16], 采用显色液(NBT/BCIP、10×Staining buffer)进行染色, 显色完成后, 终止液(25% tween-20、0.5 mol/L EDTA、pH 5.5 PBS)和PBST各洗涤3次终止染色, 然后用70%甘油进行透化置于体式显微镜下观察拍照。

表 1 引物序列Tab. 1 Primer information in this study
1.9 Real-time PCR法检测胚胎期的相对表达量
用Trizol研磨法提取不同发育阶段的黄河裸裂尻鱼胚胎总RNA, 用DNase Ⅰ(RNase-free)酶解可能残余的基因组DNA, 用1%琼脂糖凝胶电泳鉴定所提取RNA的完整性, 紫外分光光度法测定RNA样品的浓度和纯度。以不同发育阶段的胚胎总RNA为模板, 利用 PrimeScript™ RT reagent Kit (Perfect Real Time)合成第一链模板cDNA, 根据各基因内含子和外显子结构特征, 通过已获得cDNA序列跨第一内含子设计特异性引物, 并进行qRT-PCR反应。qRT-PCR反应体系10 μL: TB Green Premix ExTaqⅡ 5 μL, 上下游引物(10 μmol/μL)各0.3 μL, cDNA模板1 μL, ddH2O 3.4 μL。胚胎型血红蛋白基因qRT-PCR反应条件均为: 95℃ 30s; 60℃ 30s; 72℃1min。β-actin基因qRT-PCR反应条件为: 95℃ 30s;60℃ 30s; 72℃ 1min。实验结果采用2–∆∆Ct法分析[17],引物见表 1。
2 结果
2.1 黄河裸裂尻鱼胚胎型血红蛋白基因CDS序列特点
通过本地数据库搜索, 共鉴定到黄河裸裂尻4个胚胎型血红蛋白基因, 分别为2个α珠蛋白基因(hbae1和hbae4)和2个β珠蛋白基因(hbbe1和hbbe3)。hbae1、hbbe1、hbae4和hbbe3 四个基因均获得CDS全长序列, 长度分别为432、444、426和441 bp, 分别编码143、147、141、146个氨基酸(图 1A和1B)。搜索结果显示, 黄河裸裂尻鱼转录组中没有发现与斑马鱼hbae3、hbae5和hbbe2基因同源的序列。通过对黄河裸裂尻鱼同斑马鱼胚胎型血红蛋白基因构建系统发育树及氨基酸序列同源性分析, 结果显示: α珠蛋白基因Sphbae1/Drhbae1和Sphbae4/Drhbae4分别汇聚在第一和第三支上, 且对应的氨基酸序列同源性为88.11%、75.59%(图 1A);β珠蛋白基因Sphbbe1/Drhbbe1和Sphbbe3/Drhbbe3分别汇聚在第一和第三支上, 且对应的氨基酸序列同源性分别为82.99%和86.01%(图 1B)。另外, 通过cDNA序列比对发现黄河裸裂尻鱼4个胚胎型血红蛋白基因hbae1与hbbe1、hbbe3和hbae4核酸序列相似性为50%、45.7%和53.94%;hbbe1与hbbe3和hbae4核酸序列相似性为62.44%和43.43%;hbbe3与hbae4核酸序列相似性位为44.59%。
2.2 黄河裸裂尻鱼胚胎型血红蛋白基因结构特点
黄河裸裂尻鱼hbae1、hbbe1、hbae4和hbbe3基因序列长度分别为613、649、830和670 bp, 均由3个外显子和2个内含子构成。hbae1、hbbe1、hbae4和hbbe3基因第1个外显子长度均为90 bp, 第2长度分别为211、224、205和220 bp第3个外显子序列长度分别为131、130、131和134 bp。胚胎型hbbe1和hbae1基因以头对头(3′-5′—5′-3′)的转录方式串联在一起, 中间间隔序列长度为1411 bp; 而胚胎型hbbe3和hbae4基因则以尾对尾(5′-3′—3′-5′)的转录方式串联在一起, 中间间隔序列长度为1396 bp(图 1C)。
2.3 黄河裸裂尻胚胎期胚胎型血红蛋白基因表达特点
荧光定量PCR(qRT-PCR)检测利用qRTPCR检测了胚胎发育过程中胚胎型血红蛋白基因的表达情况, 结果显示,hbae1和hbbe1基因的表达量都是从胚胎发育第5天(120 hpf)开始明显上升(图 2),其中从胚胎发育第5天(120 hpf)至破膜后第9天(432 hpf)期间持续高水平表达, 但是从破膜后第4天(312 hpf)开始表达量逐步下降(图 2A);hbae4基因的表达量也是从胚胎发育第5天开上有上调趋势, 而破膜后(220 hpf)开始表达量下降甚至不表达, 总体而言hbae4基因在整个胚胎发育期维持较低的表达量;hbbe3基因从胚胎发育第5天(120 hpf)至破膜后第6天(360 hpf)均有表达, 而从破膜后第7天(384 hpf)开始其表达量下降甚至不表达。
整胚原位杂交检测制备得到hbae1、hbbe1、hbbe3和hbae4 RNA探针核酸序列长度分别为:304、246、207和238 bp,hbae1与hbbe1、hbbe3和hbae4核酸序列相似性为35.2%、32.7%和30.79%;hbbe1与hbbe3和hbae4核酸序列相似性为34.38%和38.08%;hbbe3与hbae4核酸序列相似性为35.29%。整胚原位杂交检测发现, 在黄河裸裂尻鱼胚胎发育早期阶段胚胎型血红蛋白基因的表达主要以hbae1和hbbe1为主,hbbe3次之, 而hbae4的表达极其微弱。hbae1、hbbe1、hbae4和hbbe3基因都是从胚胎发育第5天(120 hpf)开始出现明显的杂交信号, 其中hbae1和hbbe1基因从胚胎发育第5天(120 hpf)至破膜后第9天(432 hpf)(持续存在较强的杂交信号, 而在胚胎发育第5天(120 hpf)至破膜后第3天(288 hpf)期间杂交信号最为强烈, 从破膜后第4天(312 hpf)开始信号逐渐变弱;hbae4基因在胚胎发育的整个阶段表现出较弱的杂交信号, 特别是破膜后基本观察不到杂交信号; 而hbbe3基因从胚胎发育第5天(120 hpf)至破膜后第6天(360 hpf)都能观察到杂交信号, 其中从胚胎发育第5天至破膜后第2天(264 hpf)期间信号最为强烈, 而从破膜后第3天(288 hpf)开始信号逐渐变弱, 从破膜后第7天(384 hpf)开始基本观察不到杂交信号(图 3)。
3 讨论
3.1 黄河裸裂尻鱼胚胎型血红蛋白基因的组成及结构特征
已有研究表明, 斑马鱼基因组拥有7个胚胎型或仔鱼型血红蛋白基因, 分别为4个α珠蛋白基因(hbae1、hbae3、hbae4和hbae5)和3个β珠蛋白基因(hbbe1、hbbe2和hbbe3)[5]。本研究通过本地数据库搜索, 在黄河裸裂尻鱼转录组中只发现4个胚胎型血红蛋白基因(hbae1、hbae4、hbbe1和hbbe3), 与斑马鱼基因组相比, 黄河裸裂尻鱼缺少3个胚胎型血红蛋白基因(hbae3、hbae5和hbbe2), 那么黄河裸裂尻鱼基因组中是否缺失这3个基因?针对这一问题, 我们根据斑马鱼的hbae3、hbae5和hbbe2基因序列设计了特异性引物, 以黄河裸裂尻鱼基因组DNA为模板进行了目标基因扩增, 均没有获得特异性条带。为了进一步证实这一现象, 我们以斑马鱼hbae3、hbae5和hbbe2基因序列为搜索条件, 在NCBI数据库中利用Blast扫描了已公布的黄河裸裂尻鱼近源种-尖裸鲤(Oxygymnocypris stewartii)的基因组, 结果发现尖裸鲤基因组中也不存在这3个基因。
脊椎动物α- /β-珠蛋白基因家族是由祖先珠蛋白基因经过串联复制而来, 复制事件发生在450—500(Million years ago, MYA)[18]。大约在320—400MYA,硬骨鱼类经历了第三轮基因组复制, 这次硬骨鱼类特异性基因组复制(Teleost-specific genome duplication, TGD)事件对硬骨鱼类生理、形态和行为多样性的形成产生了深刻影响, 同时对硬骨鱼类血红蛋白基因赋予了多样性[19]。在硬骨鱼中, 血红蛋白基因分为两大基因簇, 分别位于不同的染色体, 而且胚胎型血红蛋白编码基因的种类和数量在种间存在较大差异, 这种差异是在硬骨鱼快速进化过程中基因的复制或丢失事件所造成的[19,20]。因此, 本研究初步判断hbae3、hbae5和hbbe2基因在裂腹鱼亚科鱼类进化过程中可能发生了丢失。
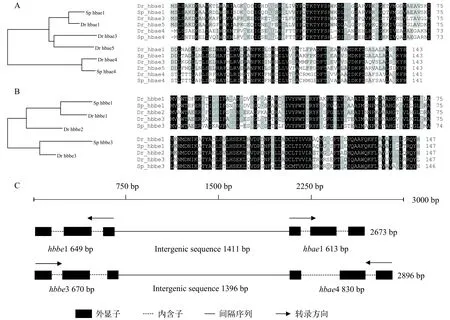
图 1 黄河裸裂尻鱼胚胎型血红蛋白基因氨基酸序列和基因结构分析Fig. 1 Amino acid sequences and gene structure analysis of embryonic hemoglobin of S. pylzovi
黄河裸裂尻鱼的4个胚胎型血红蛋白基因与其他硬骨鱼类一样具有相对保守的结构, 均由3个外显子和2个内含子组成, 但是在外显子和内含子长度上表现出其特异性[2,5,21]。如, 大黄鱼(Pseudos-ciaena crocea)等硬骨鱼类中的研究表明血红蛋白基因的第一内含子序列长度明显小于第二内含子, 在黄河裸裂尻鱼中研究发现hbae1、hbbe1和hbbe3基因符合这一特征, 但有趣的是hbae4基因的第二内含子序列长度明显小于第一内含子[21]。研究表明,哺乳动物α-/β-珠蛋白分别由位于不同染色体的基因所编码, 而在鱼类中成对α-/β-珠蛋白基因以头对头(3′-5′—5′-3′)或以尾对尾(5′-3′—3′-5′)的转录方式串联在一起[2]。如, 在斑马鱼中,hbbe1和hbae1基因以头对头的转录方式串联在3号染色体上, 而hbbe2和hbae5基因以头对头、hbbe3和hbae4基因则以尾对尾的转录方式串联在12号染色体[5,22,23]; 在青鳉(Oryzias latipes)中,hbae1和hbbe1、hbae2和hbbe2、hbae3和hbbe3三对基因都以头对头的方式串联在8号染色体上[24—26]。本研究中,hbbe1和hbae1、hbbe3和hbae4基因的串联转录方式与斑马鱼相同, 同时也表明近源种间胚胎型α-/β-珠蛋白基因具有相似或相近的表达调控方式。
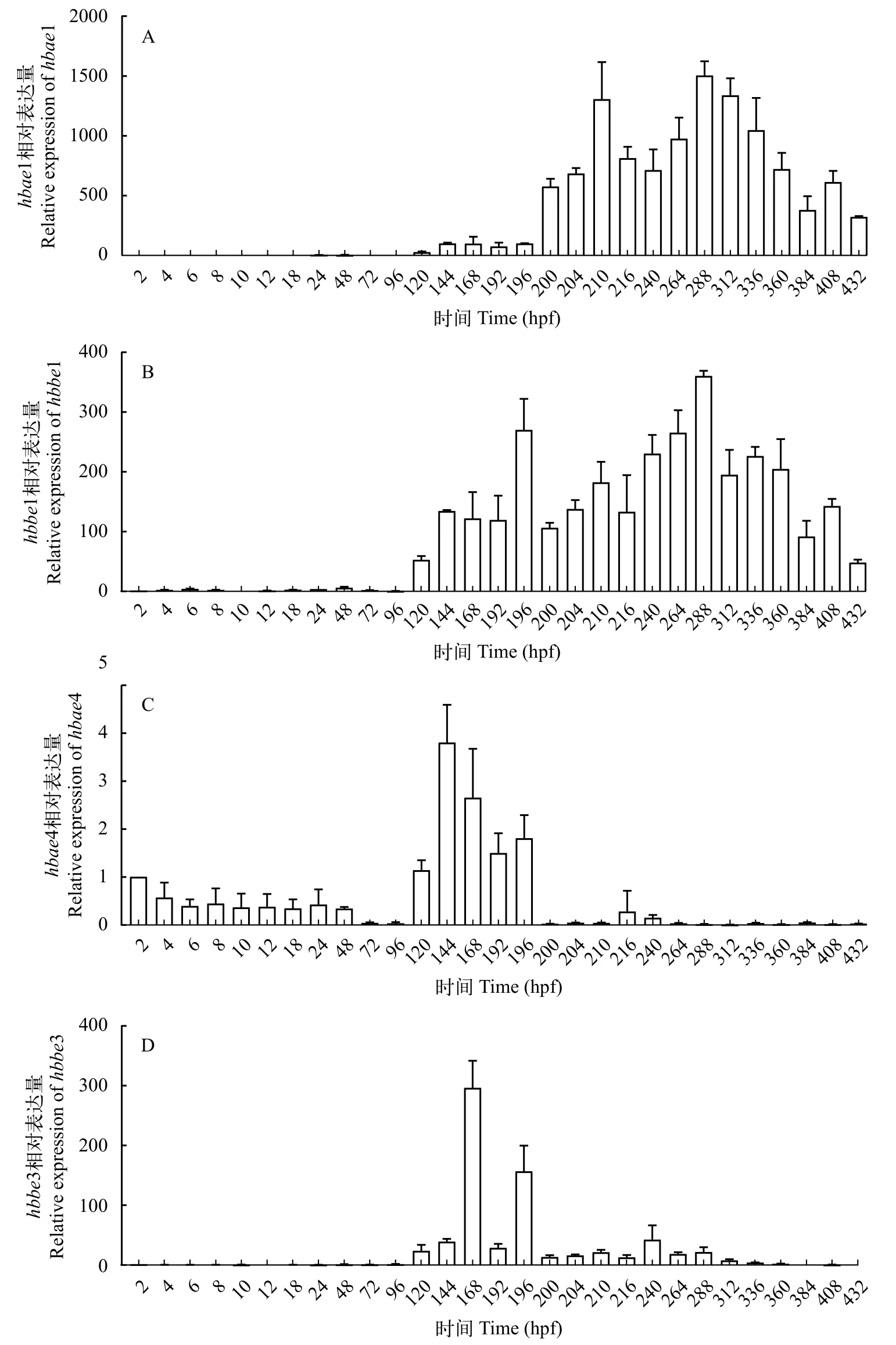
图 2 黄河裸裂尻鱼胚胎发育过程中胚胎型血红蛋白基因的相对表达量Fig. 2 Relative mRNA levels of embryonic hemoglobin genes during embryonic development of S. pylzovi
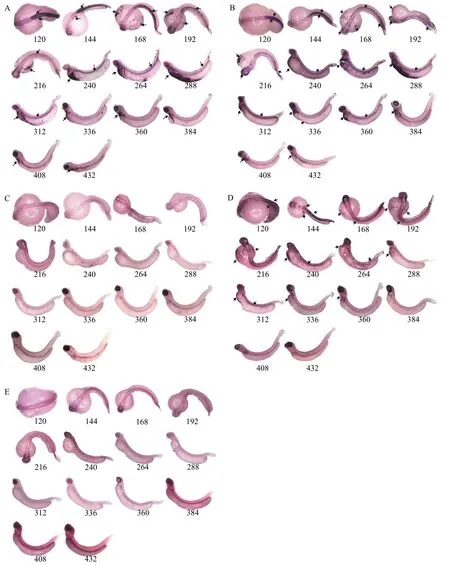
图 3 整胚原位杂交检测胚胎型血红蛋白在黄河裸裂尻鱼胚胎发育过程中的表达Fig. 3 The expression patterns of embryonic hemoglobin genes by whole mount in situ hybridization during embryonic development of S.pylzovi
3.2 黄河裸裂尻鱼胚胎型血红蛋白的时空表达特征
脊椎动物拥有多个血红蛋白编码基因, 在个体发育的不同阶段所表达的血红蛋白基因有所不同,从而实现血红蛋白的转换表达(Hemoglobin switching)。脊椎动物血红蛋白基因的多样性保证了个体发育不同阶段产生不同氧亲和力的血红蛋白, 从而满足生物体应对氧环境的改变[1,2,4]。比如, 斑马鱼基因组拥有7个胚胎型或仔鱼型血红蛋白基因, 在个体发生的16体节阶段开始就能检测到hbbe1和hbbe3基因表达, 但是胚胎期主要以hbbe3的高表达为特征; 仔鱼期hbbe2基因表达显著上升并伴随hbae5基因的表达, 同时hbbe3基因表达急剧下降,这种由胚胎型到仔鱼型血红蛋白转换表达充分满足了斑马鱼胚胎发育过程中对氧环境的适应[3,5]。另外, 青鳉基因组拥有包括两个假基因在内的11个胚胎型或仔鱼型血红蛋白基因, 在胚胎期主要以hbae2、hbae3、hbbe3、hbae4和hbbe4的表达为主要特征; 在仔鱼期以hbae0、hbae1、hbae2和hbbe1基因的表达为主要特征[24]。在本研究中, 黄河裸裂尻鱼只有4个胚胎型或仔鱼型血红蛋白基因, 而且结合qRT-RCR和整胚原位杂交结果分析表明4个基因的表达量都是从受精后第5天(120 hpf)开始明显上升, 但是hbae4和hbbe3基因的高表达只维持在受精后第5天(120 hpf)至破膜期(216 hpf), 随后急剧下降, 从破膜(216 hpf)开始hbae1和hbbe1基因表达急剧上升。由此可以看出, 黄河裸裂尻鱼从胚胎型到仔鱼型血红蛋白转换发生在受精后216h(即破膜期), 其显著特征是hbae4和hbbe3基因表达下降伴随hbae1和hbbe1基因表达急剧上升, 这与斑马鱼中以hbbe3表达下调并伴随着hbbe2上调为标志的从胚胎型向仔鱼型血红蛋白的转换表达特征截然不同[5]。在黄河裸裂尻鱼胚胎发育过程中,hbae1和hbbe1基因表达具有相似的特征, 这与这2个基因以头对头串联表达, 共享启动子和增强子引起的协同表达机制有关[24,26]。而hbbe3和hbae4基因虽然也串联在一起, 但是以尾对尾的串联方式使得2个基因分别具有不同的启动子和增强子, 从而使2个基因的表达各具特点, 这与斑马鱼中的研究一致[6]。
斑马鱼胚胎发育至16体节时, 可以检测到hbae1、hbae3、hbbe1和hbbe3基因表达, 发育至24h时, 原始前红细胞(Primitive proerythroblasts)进入血液循环[27], 但是, 在受精后24—48h内, 胚胎型血红蛋白的表达持续发生变化, 如hbae3基因的表达下降, 而hbbe2的表达上升, 这种变化与红髓系祖细胞(Erythromyeloid progenitor)的出现相一致[28]。黄河裸裂尻鱼在受精后第5天(120 hpf)左右胚胎达到相当于斑马鱼的16体节期, 发育至第8天(192 hfp)左右能够观察到明显的血液循环和心跳, 期间胚胎型血红蛋白的表达具有明显的改变, 这种特征与斑马鱼中的研究相似, 但是黄河裸裂尻鱼胚胎发育期发生表达变化的基因类型与斑马鱼完全不同[5], 这可能与黄河裸裂尻鱼特殊的进化历程和高原环境的适应有关。
猜你喜欢
农村科学实验(2022年16期)2022-11-19
检验医学与临床(2022年18期)2022-09-27
中国典型病例大全(2022年12期)2022-05-13
转化医学杂志(2022年1期)2022-03-07
水产科学(2020年4期)2020-07-28
检验医学与临床(2018年13期)2018-07-12
中国现代医学杂志(2017年22期)2017-10-11
家庭百事通·健康一点通(2017年2期)2017-02-23
大众健康(2015年5期)2015-09-10
现代检验医学杂志(2015年1期)2015-02-06
