
Herbal cake-partitioned moxibustion inhibits colonic autophagy in Crohn’s disease via signaling involving distinct classes of phosphatidylinositol 3-kinases
2020-12-11 07:10ShiYuanWangJjiMengZhaoCiLiZhouHanDanZhengYanHuangMinZhaoZhiYingZhangLuYiWuHuanGanWuHuiRongLiu
World Journal of Gastroenterology 2020年39期
Shi-Yuan Wang, Jji-Meng Zhao, Ci-Li Zhou, Han-Dan Zheng, Yan Huang, Min Zhao, Zhi-Ying Zhang, Lu-Yi Wu, Huan-Gan Wu, Hui-Rong Liu
Abstract
Key Words: Crohn’s disease; Colon; Moxibustion; Macroautophagy; Immunity; Phosphatidylinositol 3-kinase signaling
INTRODUCTION
Crohn's disease (CD), a chronic inflammatory gastrointestinal disease, is a form of inflammatory bowel disease (IBD) characterized by abdominal pain, diarrhea, anal lesions, and systemic symptoms of varying degrees of severity. CD features recurrent and remitting changes that can affect all parts of the digestive tract, with the most common being the terminal ileum and colon[1]. A population-based cohort study of diagnosed IBD patients from 13 countries in the Asia-Pacific region (2011-2013) has revealed that the overall incidence of IBD in Asia is 1.4/100000[2]. A 2018 systematic review of the epidemiology of IBD in mainland China has shown that the current incidence of IBD is 1.80/1000000, of which CD accounts for 0.46/1000000 cases[3]. The pathogenesis of CD has not been fully elucidated. At present, it is believed that CD onset is caused by abnormal factors such as genetic susceptibility, environmental factors, and intestinal flora imbalance that result in an abnormal intestinal mucosal immune response and impaired epithelial barrier function[4].
Macroautophagy (commonly shortened to “autophagy”) is an evolutionarily conserved biological process that occurs in eukaryotic cells and is mediated by lysosomal degradation and recycling of related cell components. Under normal conditions, autophagy occurs at a relatively low level, but it is activated when cells are stimulated by starvation, hypoxia, exposure to toxic molecules, or other stresses. Autophagy maintains intracellular homeostasis and promotes cell survival by decomposing related intracellular substrates[5]. Increasing evidence suggests that dysregulation of autophagy is inextricably linked to the onset of CD[6,11]. For example, autophagic dysfunction can reduce the restriction of intracellular replication of adherent-invasiveE. coli(AIEC) and increase the secretion of proinflammatory factors such as tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) in the intestine[12,13]. Autophagy in intestinal epithelial cells (IECs) may play a protective role in preventing TNF-induced apoptosis of IECs and thus play a role in limiting intestinal inflammation[14]. In-depth study of autophagy will help to further elucidate the pathogenesis of CD.
Mammalian target of rapamycin (mTOR) is a key mediator that regulates autophagy in response to cellular nutritional levels, hypoxia, and growth factors. As mTOR is a major regulator of autophagy, inhibiting mTOR is an important way to increase autophagy levels[15]. As a direct downstream target of phosphatidylinositol 3-kinase (PI3K) and Akt, mTOR can be activated by neurotrophic factors and growth factor receptors, thereby promoting cell growth, differentiation, and survival and simultaneously downregulating related apoptosis signals. Studies have shown that insulin, insulin-like growth factor (IGF), and epithelial growth factor (EGF) can activate PI3K[16-18], which promotes Akt (site 473) phosphorylation, and increases in Akt phosphorylation can directly or indirectly lead to increases in the phosphorylation of specific sites on mTOR downstream (such as Ser2448). Therefore, activation of the PI3K/AKT/mTOR pathway should in principle play a role in autophagy inhibition. In contrast, induction of autophagy (inhibition of mTOR) may impair cell survival by causing continuous activation of autophagy[19]. PI3K family members can be divided into 3 types; type I consists of regulatory and catalytic subunits (PI3Kα, β, γ, and δ), and its main pathway is the above mentioned PI3K/Akt/mTOR, whereas type III consists of only one member, the class III PI3K (PI3KC3/Vps34). Vps34 is also a major player in autophagy that mainly forms a complex with coiled-coil myosin-like BCL2-interacting protein (Beclin-1) to promote the formation of autophagosomes, and autophagy is inhibited when the formation of this complex is blocked. Therefore, the PI3K/Akt/mTOR pathway and the Vps34-Beclin-1 complex may play coregulatory roles in autophagy[20]. For example, administration of the mTOR-specific inhibitor rapamycin can lead to downregulation of mTOR phosphorylation and directly activate autophagy, and mTOR inhibition can directly lead to activation of the autophagyrelated protein unc-51-like kinase 1 (ULK1). When ULK1 is activated, Beclin-1 is phosphorylated, which enhances the activity of the Vps34-Beclin-1 complex and further enhances autophagic activity[21].
Herbal cake-partitioned moxibustion (HM), an important type of moxibustion, has the characteristics and advantages of moxibustion combined with traditional Chinese medicine, and its clinical application scope has exceeded that of conventional moxibustion. Our previous research has shown that HM improves intestinal epithelial morphology and epithelial cell structure and increases the expression of tight junction proteins to repair damage to the intestinal epithelial barrier by upregulating tumor necrosis factor alpha-induced protein 3 (A20) expression[22,23], alleviates colon tissue ulcers and relieves intestinal inflammation by downregulating IL-17 and IL-23 mRNA and protein expression in the colonic mucosa in CD model rats[24], alleviates colonic inflammation through regulation of Toll-like receptor 4 (TLR4), which induces nuclear factor κB (NF-κB) signal transduction[25], and ameliorates visceral pain and downregulates extracellular signal-regulated kinase, substance P, and neurokinin-1 protein and mRNA expression in the dorsal root ganglia in IBD[26]. However, there have been few studies on the regulation of autophagy in CD, and the related mechanism is unclear. Therefore, this study was conducted to explore the regulatory effects and mechanism of HM in CD rats from the perspective of colonic autophagy to provide a scientific basis for research on related signal transduction mechanisms and clinical applications of moxibustion in the treatment of CD.
MATERIALS AND METHODS
Chemicals and reagents
2,4,6-trinitrobenzenesulfonic acid (TNBS; P2297, Sigma), anti-IL-18 (ab191860, Abcam), anti-TNF-α (ab6671, Abcam), anti-NF-κB p65 (ab16502, Abcam), anti-microtubuleassociated protein 1 Light chain 3 beta (LC3B; #3868, Cell Signaling Technology, CST), anti-sequestosome 1 (p62; #23214, CST), anti-Beclin-1 (#3738, CST), anti-phosphomTOR (p-mTOR; #2971, CST), Cy3-AffiniPure goat anti-mouse IgG (H + L) (111-165-003, Jackson), HRP-labeled goat anti-rabbit IgG (H + L) (A0208, Beyotime), HRPlabeled goat anti-mouse IgG (H + L) (A0216, Beyotime), insulin (P3376, Beyotime), rapamycin (LC R-5000, LC Laboratories), anti-PI3K p85 (#4292, CST), anti-Akt (#9272, CST), anti-p-Akt (#9271, CST), anti-Vps34 (V9764, sigma), and Antifade mounting medium (Beyotime, P0126) were used in this study.
Animals
Sprague-Dawley (SD) rats (male, clean grade, 180 ± 20 g) were provided by the Experimental Animal Center of Shanghai University of Traditional Chinese Medicine. The rats were housed indoors in clean conditions under a 12-h light/dark cycle, a room temperature of 20 ± 2 °C, and a humidity of 50%-70%. After 1 wk of adaptive feeding, the formal experiments began. All animal experiments in this study were approved and performed under the guidelines of the Animal Ethics Committee of the Shanghai University of Traditional Chinese Medicine (No. PZSHUTCM200403009). Seventy rats were randomly divided into a normal control (NC) group, a CD group, an HM group, an insulin + CD (I + CD) group, an insulin + HM (I + HM) group, a rapamycin + CD (RA + CD) group, and a rapamycin + HM (RA + HM) group, with ten rats in each group. A CD rat model was established by using 2,4,6-TNBS enema (10 mg/100 g). To perform the enema, each rat was lifted upside down to expose the anus, and then a 1 mL syringe was connected to a gastric perfusion needle. The gastric perfusion needle was dipped in liquid paraffin and then slowly inserted into the anus of the rat to a depth of approximately 6-8 cm. After the injection was finished, the rats were kept in the head-down position for 5 min. The injection was repeated every 7 d for five times.
Intervention and HM treatment
Traditional Chinese medicines such asRadix Aconiti Lateralis Preparata,Cortex Cinnamomi, andRadix Salviae Miltiorrhizaewere made into powder. Yellow rice wine was poured into the powder and blended to make a paste. The paste was then made into an herbal cake with a special cake mold (0.5 cm in diameter and 1.0 cm in height). After the CD rat model was confirmed to be successfully established, the rats in the I + CD and RA + CD groups were given intraperitoneal injections of insulin and rapamycin, respectively, once daily for 7 d. The CD rats in the HM group were treated with moxa cones. The cones were made of refined mugwort floss (90 mg) placed on an herbal cake and were placed on the abdominal acupoints [Tianshu (ST25) and Qihai (CV6)] of the rats before being ignited. Two moxa cones were used for each acupoint once daily for 7 d. The CD rats in the I + HM and RA + HM groups underwent the same procedures as those in the HM group and were given intraperitoneal injections of insulin and rapamycin, respectively, once daily at the same time for 7 d.
Immunofluorescence
Rat colon samples were fixed in 40 g/L formaldehyde for 24 h, embedded in paraffin, cut into 5 μm sections, and then heated at 60 °C for 20 min. The sections were collected and soaked in dimethylbenzene for 15 min twice; soaked in 100%, 95% and 70% alcohol for 5 min each; and washed with water. Antigen repair was conducted using the microwave thermal repair method. Briefly, 0.01 mol/L citrate buffer (1000 mL with 3 g of trisodium citrate and 0.4 g of citric acid) was placed in a microwave oven and heated on a high setting. After the buffer began to boil, it was removed. Then, the sections were soaked in the buffer, put into the microwave oven, heated for 10 min, and cooled at room temperature for 1 h. The sections were then washed with phosphate buffer saline (PBS) for 5 min, blocked with goat serum for 30 min at room temperature, and incubated with a primary antibody (LC3B, 1:200) overnight at 4 °C. After the samples were incubated at 37 °C for 45 min and washed three times for 5 min with PBST, they were incubated with a secondary antibody (Cy3, 1:400) for 30 min at room temperature and then washed three times for 5 min with PBST. The samples were incubated with DAPI staining solution for 10 min and washed three times for 5 min with PBST. One drop of antifade mounting medium was added to each section, and laser confocal microscopy (Nikon, Japan) was used to detect the expression of LC3B.
Real-time qPCR
Total RNA was isolated using TRIzol reagent (Invitrogen), and cDNA was synthesized using a RevertAid First Strand cDNA Synthesis Kit (K1622, Invitrogen) according to the manufacturer’s protocol. Detection of mRNA was performed using a QuantityNova SYBR Green Kit (208052, Qiagen) and a Roche480II Real-Time PCR System (Roche). The mRNA expression levels were normalized to those of GAPDH and were calculated using the 2−ΔΔCtmethod. The primers used for PCR amplication are listed in Table 1.
Western blot analysis
Total proteins were extracted from colon tissues using RIPA buffer supplemented with protease and phosphatase inhibitors (P1046, Beyotime). Equal amounts of protein (50 µg) were separated by 8%-10% sodium dodecylsulphate polyacrylamide gel electrophoresis, and the separated proteins were transferred to polyvinylidene fluoride membranes. The membranes were blocked with 5% bovine serum albumin in Tris Buffered Saline with Tween-20 (TBST) and then incubated at 4 °C overnight with primary antibodies. Following several sequential washes, the membranes were incubated with secondary antibodies for 1 h at room temperature. The antibody concentrations/dilutions were prepared according to the instructions (IL-18 and p65, 0.5 µg/mL; Vps34, 2 µg/mL; TNF-α, LC3B, p62, Beclin-1, p-mTOR, p85, Akt, and p-Akt, 1:1000). The blots were then washed four times with TBST (10 min each time). The membranes were stained with enhanced chemiluminescence solution (Thermo) and visualized using an imager.
Transmission electron microscopy
Samples of the rat colonic mucosa were cut into 1 mm3strips and placed in glutaraldehyde fixation solution. The samples were washed for three times for 15 min each with 0.1 mol/L PBS, fixed with 2% glutaraldehyde fixative at 4 °C for at least 2 h, washed three times for 15 min each with 0.1 mol/L PBS, fixed with 2% osmium acid for 2 h, and again washed three times for 15 min each with 0.1 mol/L PBS. The samples were then dehydrated in 30%, 50%, and 70% ethanol solutions for 20 min each at 4 °C; incubated in uranyl acetate dihydrate overnight at 4 °C; placed sequentially into 80% and 90% ethanol solutions; soaked in 90% acetone: Embedding agent (1:1) for 20 min at 4 °C; soaked in 100% acetone three times for 20 min each at room temperature; Soaked in 100% acetone:embedding agent (1:1) for 2 h at room temperature; and Soaked in embedding agent two times for 1.5 h each at 37 °C. The samples were then heated in an oven at 37 °C for 12 h and at 60 °C for 48 h. For sectioning, the block of embedded sample was trimmed with a double-sided knife, and the top was cut into a pyramid shape. The top was flattened, and the tissue was exposed to facilitate slicing. A glass strip was fitted into an ultramicrotome knifemaking machine to make the glass knife, which was fixed on the stage with sticking plaster. The block of sample in embedding agent was placed on the ultramicrotome and sliced at a thickness of 80 nm. The slices were removed and placed on a support membrane of copper mesh. Lead citrate staining solution was used to dye the slices for 15 min, after which the slices were washed with distilled water three times, dried naturally, and observed under a transmission electron microscope (Leica EMUC 7, Germany). The embedding agent consisted of 618 epoxy resin 40.5 g, dodeceny succinicanhydride 36 g, dibutyl phthalate 2.7 g, and 2,4,6-Tris(dimethylaminomethyl)phenol 0.675 mL.
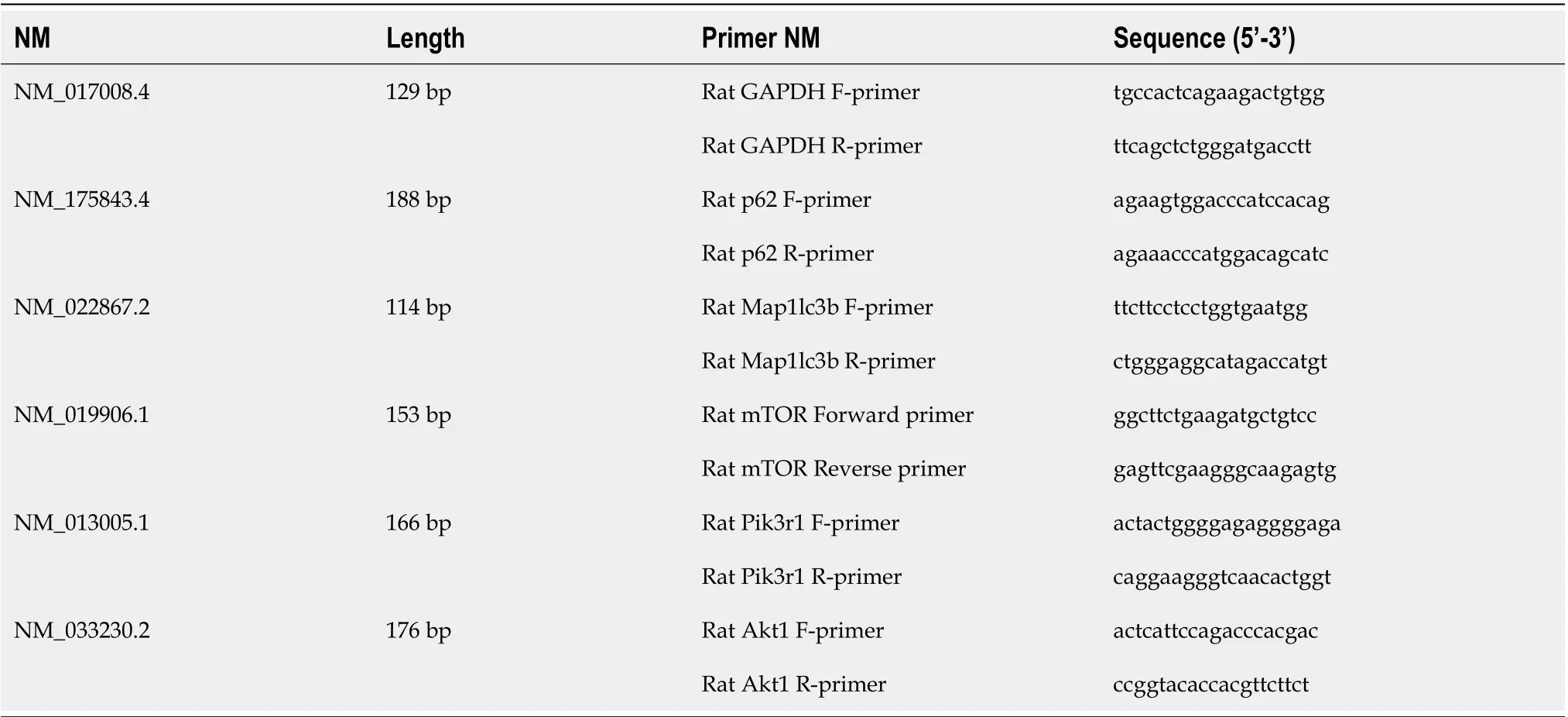
Table 1 Primers used for PCR
Statistical analysis
SPSS 20.0 statistical software was used for statistical analyses. One-way ANOVA was used for data that met the assumptions of a normal distribution and homogeneity of variance. For data that did not meet the normal distribution or homogeneity of variance assumption, a nonparametric test was used. Continuous data are presented as the mean ± standard deviation or as medians (P25, P75) according to the distribution;P< 0.05 were considered to indicate statistical significance.
RESULTS
HM ameliorates TNBS-induced inflammation and colon damage in CD rats
We used the Morris method[27]to establish a rat model of CD. After TNBS induction, the colon was shortened and exhibited megacolon or intestinal obstruction. Upon cutting of the intestine, wall thickening, mucosal surface roughness, and cobblestonelike changes were observed, accompanied by scattered edema, congestion, and ulceration, suggesting that the rat model had been successfully established (Figure 1A). After confirming the success of the modeling, we selected the Qihai (CV6) and bilateral Tianshu (ST25) acupoints for HM and performed pathological observation. In the NC group, the colon tissues showed clear and intact colonic epithelial structures, complete and continuous mucosae, and neatly arranged glands. In the CD group, considerable inflammatory cell infiltration, destruction of mucosal structure, partial mucosal shedding, disappearance or deformation of glands, and fissure-like ulcers were observed. Compared with rats in the CD group, the colon tissue of rats in the HM group showed relatively complete mucosal epithelial coverage, ulcer repair, and partial amelioration of gland abnormalities and lymphocyte infiltration (Figure 1B). Our previous research has demonstrated that HM at the Qihai (CV6) and Tianshu (ST25) acupoints in CD rats can relieve intestinal inflammation by downregulating IL-17 and IL-23[24]. In the present study, we tested the expression levels of IL-18, TNF-α, and NF-κB in the colon tissues of the rats in each group. Compared with the NC group, the CD group exhibited significantly higher levels of IL-18 protein and NF-κB p65 in colon tissue (P< 0.01 for both). Compared with the CD group, the HM group exhibited significantly lower levels of these proteins (PIL-18< 0.01 andPp65< 0.05). There were no significant differences in the expression of TNF-α protein in colon tissue among the groups. However, compared with the NC group, the CD group exhibited a trend toward higher TNF-α protein expression, and compared with the CD group, the HM group exhibited a trend toward lower TNF-α protein expression (Figure 1C).
HM facilitates the repair of colonic epithelial cells
Since the intestinal mucosa is composed of a simple columnar epithelium that serves as a physical and chemical barrier and an absorptive surface[28], we next observed the damage, repair, and autophagosome formation of colon tissue epithelial cells in the rats in each group by electron microscopy (Figure 2). In the NC group, the colonic epithelial cell structure was intact, the organelles and microvilli were arranged neatly without damage, and no autophagosomes were present. In the CD group, injury, swelling, and deformation of organelles were observed. Typical autophagic vesicles were found that had double-layer membrane structures and contained damaged organelles. In the HM group, typical autophagic vesicles were still present, and organelle deformation and swelling could be observed, but these features were milder than those in the CD group, and repair of microvillus damage could be seen. These findings suggested that inflammation activated autophagy and that HM promoted the repair of colonic epithelial cells, but it remained unclear whether HM could regulate autophagy.
TF/IDF方法只考虑了词在上下文中的统计特性,具有一定的局限性。一个特征项出现的频率越多,该特征项越能代表主题,包含该特征项的文档越少,该特征项越能代表主题,这是正确的,但也是片面的。从词性上讲,名词比动词或者其他词性更能代表主题,从词汇属性上讲,关键词、方言词比普通常用词汇更能代表主题。本文利用的是项目自定义的农业专业分词算法,该算法的分词结果包括该词的词性和词汇属性(常用词、方言词、专业关键词),将词汇按照其词性及词属性进行加权,增加了该算法的可操作性和准确度。
HM inhibits colonic autophagy in CD rats
Next, we observed the effect of HM on the colonic levels of autophagy proteins in each group of rats. As shown in Figure 3A, compared with the NC group, the CD group and the HM group exhibited significantly higher expression of LC3B protein in colon tissue (P< 0.01 for both). Compared with the CD group, the HM group exhibited significantly lower expression of LC3B protein in colon tissue (P< 0.01). Both p62 and LC3 are used to evaluate autophagy, and studies have shown that the expression levels of p62 decrease with increasing LC3 Levels[29]. Therefore, we tested the expression levels of p62 protein in colon tissues. As expected, compared with the NC group, the CD group and the HM group exhibited significantly lower expression of p62 protein in colon tissue (P< 0.01 for both). In addition, compared with the CD group, the HM group exhibited significantly higher expression of p62 protein in colon tissue (P< 0.05). Compared with the NC group, the CD group exhibited significantly higher expression of Beclin-1 protein in colon tissue (P< 0.01). Compared with the CD group, the HM group exhibited significantly lower expression of Beclin-1 protein in colon tissue (P< 0.05). Furthermore, compared with the NC group, the CD group and the HM group exhibited significantly lower expression of p-mTOR (ser 2448) protein in colon tissue (P< 0.01 andP< 0.05, respectively). Compared with the CD group, the HM group exhibited significantly higher expression of p-mTOR protein in colon tissue (P< 0.05). Immunofluorescence showed that LC3B was expressed in different layers of the colon, especially in the lamina propria and submucosa. Only a few positive puncta were observed in the NC group, while strong positive staining was observed in the CD group. Compared with the CD group, the HM group had obviously weaker positive staining (Figure 3B). The above results suggested that HM may have exerted a therapeutic effect by inhibiting autophagy in colon tissue in CD rats, but the mechanism was still unclear.
HM regulates autophagy through PI3K signaling

Figure 2 Ultrastructure of colonic epithelial tissue and autophagic vesicles in each group of rats. Scale bar: 2 μm. NC: Normal control; CD: Crohn’s disease; HM: Herbal cake-partitioned moxibustion.
The activation of the PI3K/Akt/mTOR signaling pathway by insulin has been confirmed[30]. Specifically, insulin upregulates Akt phosphorylation and activates mTOR complex 1 (mTORC1), thereby inhibiting autophagy[31-33]. The protein mTOR is downstream of the PI3K and AKT pathway[19], and studies have shown that rapamycin can inhibit cyclophosphamide-induced activation of the PI3K/Akt/mTOR signaling pathway, thus activating autophagy[34,35]. In addition, rapamycin and its derivatives can not only inhibit mTORC1, but also inhibit the phosphorylation of Akt by inhibiting mTORC2 after long-term application[36,37]. Thus, we used insulin to activate the PI3KC1/Akt1/mTOR pathway to inhibit autophagy and used rapamycin to inhibit the PI3KC1/Akt1/mTOR pathway to activate autophagy, and subsequently observed the changes in the PI3K signaling pathway in the colon tissues of the rats in each group. The hematoxylin-eosin staining results showed that the colon tissue structure was severely damaged and that the glands had disappeared in both the CD and RA + CD groups. In addition, fissure-like ulcers and granulomas were found. In the other groups, healing ulcers, epithelial hyperplasia, partial loss of glands, and lymphocyte infiltration were observed (Figure 4). Immunofluorescence showed that compared with the CD and RA + CD groups, the I + CD, I + HM, and RA + HM groups exhibited lower expression of LC3B (Figure 5). In addition, compared with the RA + CD group, the RA + HM group exhibited significantly higher expression ofPI3KC1,Akt1,mTOR, andp62mRNA (PPI3KC1< 0.01 andPAkt1,mTOR,andp62< 0.05) but significantly lower expression ofLC3BmRNA (P< 0.05) (Figure 6A). Compared with the RA + CD group, colon tissue in the RA + HM group exhibited significantly higher expression of PI3KC1 protein (P< 0.01), significantly higher expression of p-Akt1 and p-mTOR proteins (P< 0.05 andP< 0.01, respectively), and higher expression of p62 protein (P= 0.057). LC3B protein expression was significantly downregulated in the RA + HM group (P< 0.01) (Figure 6B). Previous research has indicated that administration of the mTOR-specific inhibitor rapamycin can downregulate mTOR phosphorylation, directly activate autophagy, and cause phosphorylation of the autophagy-related protein Beclin-1, thus enhancing the activity of the Vps34-Beclin1 protein complex[21]and further enhancing autophagic activity. In addition, studies have shown that insulin can inhibit the autophagy-related genePI3KC3/Vps34[38]. Since the expression of Beclin-1 could be inhibited by HM treatment (Figure 3A), thus, we observed the protein expression levels of Vps34 in the colon tissues of the rats in each group. The results showed that HM could significantly downregulate the expression of Vps34 protein in the colon tissues of CD rats. Compared with the CD group, the expression of Vps34 protein in RA + CD group was significantly upregulated (P< 0.01). Compared with the RA + CD group, Vps34 protein expression was significantly downregulated in the I + CD, I + HM, and RA + HM groups (P< 0.01 for all; Figure 6B).

Figure 3 Herbal cake-partitioned moxibustion at the Qihai (CV6) and bilateral Tianshu (ST25) acupoints regulates the expression of autophagy proteins in the colon tissues of Crohn’s disease rats. A: Expression changes of the microtubule-associated protein 1 light chain 3 beta, sequestosome 1, coiled-coil myosin-like BCL2-interacting protein, and phospho-mammalian target of rapamycin proteins evaluated by Western blot. Six independent experiments were analyzed, and the data are presented as the mean ± SD or medians (P25, P75). aP < 0.01, bP < 0.05 vs NC group; cP < 0.01, dP < 0.05 vs CD group; B: Immunofluorescence images for LC3B (Red) in each group. Scale bar: 200 μm. NC: Normal control; CD: Crohn’s disease; HM: Herbal cake-partitioned moxibustion; LC3B: Microtubule-associated protein 1 light chain 3 beta; p62: Sequestosome 1; Beclin-1: Coiled-coil myosin-like BCL2-interacting protein; p-mTOR: Phospho-mammalian target of rapamycin.
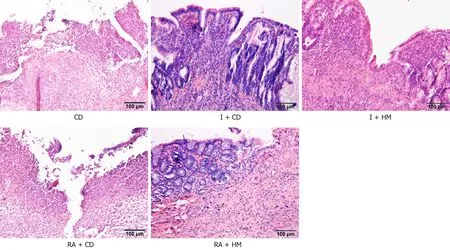
Figure 4 Histopathological observations of colon tissue from each group by hematoxylin and eosin staining. Scale bar: 100 μm. CD: Crohn’s disease; I + CD: Insulin + Crohn’s disease; I + HM: Insulin + herbal cake-partitioned moxibustion; RA + CD: Rapamycin + Crohn’s disease; RA + HM: Rapamycin + herbal cake-partitioned moxibustion.
DISCUSSION
At present, autophagy is broadly divided into three types: Macroautophagy, microautophagy, and molecular chaperone-mediated autophagy[39]. Among them, macroautophagy (commonly shortened to “autophagy”) has received the most attention in research. This type of autophagy can be triggered by different stimuli, such as starvation, inflammation, oxidative stress, hypoxia, and toxic molecules. Studies have shown that dysregulation of autophagy is inextricably linked to the occurrence and development of CD[6,7]. Thus, in the present study, we used the Morris method to establish a rat model of CD and observed the effects of TNBS on colonic autophagy in rats. The results showed that after TNBS stimulation, the colons of the rats showed severe inflammation and damage; they also showed significant activation of autophagy, as indicated by the upregulated protein expression of LC3B and Beclin-1 and the downregulated protein expression of p62 and p-mTOR and as further validated by transmission electron microscopy. In addition, treatment with rapamycin (a widely used autophagy activator) did not attenuate the pathological damage in the colon; however, HM and/or insulin treatment not only greatly ameliorated colon damage but also inhibited colonic autophagy, suggesting that under inflammatory conditions, overactivation of autophagy is harmful to colon tissues. This finding is consistent with the findings of previous studies[40,41].
Among the autophagy-related proteins, the kinase mTOR is a major regulator of autophagy. It can receive input from different signaling pathways and become inactivated under stress conditions, thereby activating ATG1 kinases (mammalian homologs of unc-51-like autophagy-activating kinases 1 and 2) and initiating autophagosome formation, thus activating autophagy[39]. Our results showed that the expression of p-mTOR protein in colon tissues was significantly downregulated in the CD group but that HM treatment reversed this downregulation, suggesting that TNBS may activate autophagy through the mTOR pathway and that mTOR may be one of the targets of HM therapy.

Figure 5 Immunofluorescence images for microtubule-associated protein 1 light chain 3 beta (Red) in each group. Scale bar: 200 μm. CD: Crohn’s disease; I + CD: Insulin + Crohn’s disease; I + HM: Insulin + herbal cake-partitioned moxibustion; RA+CD: Rapamycin + Crohn’s disease; RA + HM: Rapamycin + herbal cake-partitioned moxibustion; LC3B: Microtubule-associated protein 1 light chain 3 beta.
mTOR is downstream of PI3K/Akt signaling[42], and studies have shown that inhibiting the PI3K/Akt/mTOR pathway enhances autophagy and induces apoptosis[31,43], while activation of the PI3K/Akt/mTOR pathway inhibits autophagy[44]. PI3K is an enzyme that catalyzes the phosphorylation of inositol phospholipids at position 3 of the inositol ring. Its function is to generate specific molecular messengers on the cell membrane. There are eight types of PI3Ks in mammalian cells that are divided into three classes: Class I, which includes four isomers (PI3KC1α, β, γ, and δ); Class II, which includes three isomers (PI3KC2α, β, and γ); and Class III, which includes a single isomer (PI3KC3)[45,46]. PI3KC1 is the main type involved in the PI3K/Akt signaling pathway, and phosphorylation and activation of the serine kinase PKB (also known as AKT) appears to be a general response to class I PI3K activation[47]. Activation of the PI3K/Akt/mTOR pathway is initiated by interactions between PI3K and transmembrane G protein-coupled receptors or receptor tyrosine kinases such as IGF and fibroblast growth factor that lead to Akt phosphorylation; Akt phosphorylation then leads to activation of downstream mTOR[42]. Therefore, activation of the PI3K/Akt pathway can promote mTOR activation and inhibit autophagy, which in turn may lead to upregulation of autophagy. Based on the relationship between the PI3K/Akt/mTOR signaling pathway and autophagy mentioned above, we used insulin to inhibit autophagy and used rapamycin to activate autophagy. The results showed that after administration of rapamycin to CD rats, the mRNA expression ofPI3K,Akt,mTOR, andp62and the protein expression of PI3K, p-Akt, p-mTOR, and p62 in colon tissues were further downregulated, while the mRNA and protein expression of LC3B was further upregulated. These changes indicated that autophagy in colon tissue was further upregulated. Notably, the changes in these indicators were reversed by HM or/and insulin treatment. These results suggest that HM can inhibit excessively activated autophagy during CD by activating the PI3K/Akt/mTOR signaling pathway.

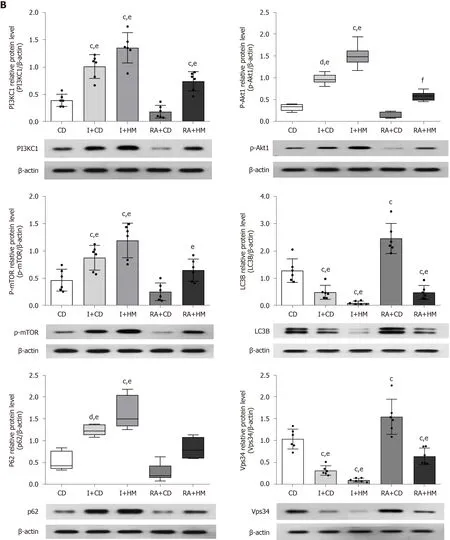
Figure 6 Herbal cake-partitioned moxibustion at the Qihai (CV6) and bilateral Tianshu (ST25) acupoints regulates the mRNA and protein expression of PI3KC signaling-related molecules in the colon tissues of Crohn’s disease rats. A: The mRNA levels of phosphatidylinositol 3-kinase class I (PI3KC1), Akt1, mammalian target of rapamycin (mTOR), microtubule-associated protein 1 light chain 3 beta (LC3B), and sequestosome 1 (p62) were determined by real-time PCR; B: Expression changes of the PI3KC1, p-Akt1, mTOR, LC3B, p62, and class III phosphatidylinositol 3-kinase proteins evaluated by Western blot. Six independent experiments were analyzed, and the data are presented as the mean ± SD or medians (P25, P75). cP < 0.01, dP < 0.05 vs CD group; e P < 0.01, fP < 0.05 vs RA+CD group. CD: Crohn’s disease; I + CD: Insulin + Crohn’s disease; I + HM: Insulin + herbal cake-partitioned moxibustion; RA+CD: Rapamycin + Crohn’s disease; RA + HM: Rapamycin + herbal cake-partitioned moxibustion; PI3KC1: Phosphatidylinositol 3-kinase class I; Akt1: Protein kinase B akt-1; p-Akt1: Phospho-protein kinase B akt-1; mTOR: Mammalian target of rapamycin; p-mTOR: Phospho-mammalian target of rapamycin; LC3B: Microtubuleassociated protein 1 light chain 3 beta; p62: Sequestosome 1; Vps34: Class III phosphatidylinositol 3-kinase.
PI3K class III currently contains only one member, PI3KC3, also known as Vps34[48], which is one of the main participants in autophagy[49]. Vps34 complexes with Beclin-1 (an autophagy-related protein 6 homolog) and provides a PI3P-rich domain that mediates autophagosome membrane formation[20,39]. Vps34-Beclin-1 complexes have been identified in mammals to play an important role in regulating autophagy[50]. For example, acetylation of Vps34 can inhibit the formation of the Vps34-Beclin 1 complex, thereby suppressing autophagy[51]. In contrast, phosphorylation of Beclin-1 can enhance the activity of the Vps34-Beclin1-ATG14L complex, thereby enhancing autophagy[52]. Studies have shown that insulin has an inhibitory effect on Vps34[38]and that inactivation of Vps34 enhances insulin-related effects[53]. Rapamycin can directly activate ULK1 by inhibiting mTOR; after activation of ULK1, rapamycin phosphorylates Beclin-1 at the Ser14 site, thereby strengthening Vps34-Beclin1-ATG14 complex activity, which is essential for autophagic vesicle formation and initiation of autophagy[21,54]. Based on the above findings, we observed the effect of HM on the Vps34-Beclin-1 complex core protein Vps34 in colon tissues. As expected, compared with the CD group, the CD + RA group exhibited further upregulation of VPS34 protein expression, and the Vps34 protein expression level was significantly downregulated after HM and/or insulin treatment. These results suggest that HM can inhibit overactivated autophagy by inhibiting the PI3KC3 (Vps34)-Beclin-1 protein complex.
CONCLUSION
In summary, our results suggest that HM can inhibit TNBS induced excessive autophagy in colon tissues of CD rats by activating PI3KC1/Akt1/mTOR signaling while inhibiting the PI3KC3 (Vps34)-Beclin-1 protein complex. Additionally, our findings raise the question of whether insulin has dual therapeutic effects on diabetic patients with CD. However, since the PI3KC signaling pathway may have different mechanisms between the two diseases, further clinical observations are needed to answer this question.
ARTICLE HIGHLIGHTS


Research results
Our experiments revealed that TNBS-induced inflammation activated autophagy in rat colon tissues. HM at the Qihai (CV6) and bilateral Tianshu (ST25) acupoints reversed the expression changes in the autophagy proteins LC3B, p62, Beclin-1, and pmTOR in colon tissues while ameliorating colonic inflammation (indicated by IL-18, TNF-α, and NF-κB p65 levels) and damage. Furthermore, HM facilitated the repair of colonic epithelial cells. We hypothesized that the mechanisms by which HM alleviates CD may involve the PI3KC1/Akt1/mTOR pathway and the PI3KC3 (VPS34)-Beclin-1 protein complex and confirmed this hypothesis through our experiments.
Research conclusions
HM can activate the PI3KC1/Akt1/mTOR signaling pathway while inhibiting the PI3KC3 (VPS34)-Beclin-1 protein complex, thereby inhibiting overactivated colonic autophagy in CD rats.
Research perspectives
Our findings provide insights into the molecular mechanism of HM and shed new light on a cost-effective and safe therapy for CD.
猜你喜欢
上海人大月刊(2022年4期)2022-04-14
作文通讯·初中版(2022年2期)2022-02-05
校园英语·月末(2021年13期)2021-03-15
中华胰腺病杂志(2021年1期)2021-02-26
人大建设(2020年5期)2020-09-25
人大建设(2020年5期)2020-09-25
智富时代(2019年6期)2019-07-24
智富时代(2019年6期)2019-07-24
文理导航·趣味课堂(2016年5期)2016-07-21
文理导航·趣味课堂(2016年4期)2016-06-01
 World Journal of Gastroenterology2020年39期
World Journal of Gastroenterology2020年39期
- World Journal of Gastroenterology的其它文章
- Use of artificial intelligence in improving adenoma detection rate during colonoscopy: Might both endoscopists and pathologists be further helped
- Real-world treatment attrition rates in advanced esophagogastric cancer
- Metastatic pattern in esophageal and gastric cancer: Influenced by site and histology
- Relationships of early esophageal cancer with human papillomavirus and alcohol metabolism
- Dynamic contrast-enhanced magnetic resonance imaging and diffusion-weighted imaging in the activity staging of terminal ileum Crohn's disease
- Clinical assessment and management of liver fibrosis in non-alcoholic fatty liver disease