
The Effect of Attendance Patterns on the Lek Mating System in the Omei Treefrog (Rhacophorus omeimontis)
2020-12-30 06:59YueGUOWenxiaWANGWeiZANGHuiWANGChenliangLIMianZHAOandHuaWU
Asian Herpetological Research 2020年4期
Yue GUO,Wenxia WANG,Wei ZANG,Hui WANG,Chenliang LI,Mian ZHAO and Hua WU
Institute of Evolution and Ecology,School of Life Sciences,Central China Normal University,Wuhan 430079,Hubei,China
Abstract Attendance patterns are a key factor affecting sexual selection and mating systems in lekbreeding species.However,there are few systematic studies on attendance patterns in lek-breeding anurans.In this study,we monitored a population of Omei treefrogs (Rhacophorus omeimontis) for four consecutive years to reveal in detail the effects of attendance patterns on mating systems.We found that (1) female nightly attendance was affected by male nightly attendance,and operational sex ratios (determined by male and female nightly attendance) ultimately determined the extent of female polyandry; (2) males surviving more than one breeding season maintained high chorus attendance and mating success,suggesting that male chorus attendance is a good predictor of male genetic quality; (3) the distributions of male mating success were not significantly different from a Poisson distribution,indicating a random mating pattern;and male mating success was not related to body size(size-assortative mating was not evident) but was significantly correlated with chorus attendance.Since the chorus attendance is a predictor of male genetic quality,females performing random mating were more likely to mate with more frequently present males and could therefore gain indirect benefits from mating with higher-than-average quality males,thereby producing good offspring without incurring the significant costs involved in prolonged searching for,and assessing males.Our systematic survey on the relationship between attendance patterns and the lek mating system in R.omeimontis could help us better understand in-depth anuran lek mating systems.
Keywords attendance,genetic quality,lek mating,random mating,Rhacophorus omeimontis
1.Introduction
In lek mating systems,males aggregate at sites where they perform their competitive displays to attract females,who can assess the quality of potential mates allowing them to choose the best male to mate with (Hoglund and Alatalo,1995).In lek-breeding species,male mating success is primarily influenced by two distinct mechanisms of sexual selection:endurance rivalry and female choice (Andersson and Iwasa,1996).Endurance rivalry generally favors males with higher-than-average lek attendance while female choice primarily selects for males with certain phenotypes (Murphy,1998).In some lek-breeding species showing endurance rivalry,females mate randomly with males and gain only sperm from the males,suggesting that females do not obtain obvious direct benefits from leks (Hoglund and Alatalo,1995;Andersson and Iwasa,1996; Botto and Castellano,2016).Therefore,an important question is whether females can gain indirect benefits in these random-mating species.
Anuran amphibians provide a good model to address this question.Many anuran species have a prolonged breeding season and show a lek mating systems where females mate randomly with males who provide no obvious contribution besides the sperm (Hoglund and Alatalo,1995).On each breeding night,males arrive at the breeding site first and then commonly join in competitive and/or enticing choruses to attract females; females arrive later,asynchronously,and may show consistent preferences for mating with particular partners.The ratio of male to female nightly attendance (the nightly number of males and females at the breeding site,hereafter referred to as ‘nightly attendance’) can represent a nightly operational sex ratio (OSR),as determined by the ratio of sexually competing males to sexually receptive females at the site and time when mating occurs (Clutton-Brock,2007).This can reflect the intensity and direction of sexual selection (Byrne and Roberts,2004; Weiret al.,2011;Janicke and Morrow,2018).The increased male biased OSR in anuran species commonly leads to intense sexual selection (Byrne and Roberts,2004; Hase and Shimada,2014).Males that spend more time in leks appear to have a higher chance of meeting and mating with females.Accordingly,a positive correlation of male mating success and male chorus attendance (the number of nights males spent calling at the breeding site,hereafter referred to as ‘chorus attendance’) has been found in many lekbreeding anuran species,even under random female mating preference (Friedl and Klump,2005; Castellanoet al.,2009).However,high chorus attendance does not always indicate genetic quality,but may instead suggest good current body conditions; anuran males may show a trade-off between current and future reproductive output favoring short-time high chorus attendance when they have no advantage in terms of genetic quality (Friedl and Klump,2005).Consequently,male chorus attendance can be regarded as a reliable indicator of male genetic quality when males with high chorus attendance maintain this behavior in the next breeding season.In this situation,random mating is a beneficial strategy for females because they gain benefits without investing time and energy or exposing themselves to predators in search of potential mates.To date,few studies have investigated anuran male chorus attendance over successive years(Friedl and Klump,2005).
The Omei treefrog (Rhacophorus omeimontis) is endemic to central and southwestern China,found at the elevations of 700-2000 m (Liao and Lu,2010; Feiet al.,2012).As a prolonged breeder,R.omeimontisbreeds between April and July (Liao and Lu,2010; Feiet al.,2012).On each breeding night,males commonly gather around a permanent or temporary pond earlier than females,and then join in choruses to attract mates(Liao and Lu,2010).Females entering the lek are generally grasped quickly by a male,and amplectant pairs climb up nearby trees whose leaves protrude above the pond,providing substrate for spawning and hatching of eggs(Liao and Lu,2010).Any peripheral males present commonly join the spawning group after the oviposition has begun(Liao and Lu,2010).The copulation generally lasts for several hours.After spawning,mated males leave the mating group first (Liao and Lu,2010).The female secretes a foamy substance which combines with the eggs and the sperms to form a foam nest on the leaf.The eggs then develop and hatch in the nest before dropping into the pond below.Each female produces a single egg clutch per breeding season.
Previous studies have revealed female polyandry inR.omeimontispopulations(Liao and Lu,2010; Zhaoet al.,2016).Compared to female choice,endurance rivalry between males plays a more important role in sexual selection and group spawning inR.omeimontis,likely because the cost of choosing mates is high forR.omeimontisfemales(Luoet al.,2016).However,detailed information on what determines the extent of female polyandry and the outcome of mating without female choice is lacking.In this study,we observed and recorded the breeding behavior of a singleR.omeimontispopulation for four consecutive breeding seasons (2011-2014).We first analyzed the relationship between the operational sex ratio and the extent of female polyandry.We then analyzed whether females indeed preformed random mating or whether the pattern resulted in size-assortative mating.Finally,we assessed whether male chorus attendance reflected male genetic quality based on data from multiple years.
2.Materials and Methods
2.1.Study siteWe conducted fieldwork a t the Badagongshan National Nature Reserve (29°38′-29°49′ N,109°41′-110°10′ E) in Sangzhi County,Hunan Province,central China.It’s located in the zone of subtropical monsoon climate,characterized by humid summers.According to data from the China Meteorological Data Service Center,the mean ambient temperature is 16-24 ℃and the average relative humidity is 78%-81% from April to June.The average rainfall increased since April and reached the maximum value in June and July.We focused on a population ofR.omeimontisthat gathered around a permanent pond (29°47′ N,110°05′ E,elevation:1413 m)at the foot of a hill during the breeding period.The pond was relatively small,measuring 5 m2and < 50 cm deep.Branches of cedar trees around the pond stretched out over the water surface,offering oviposition sites forR.omeimontisfemales.
2.2.Field observation and data/sample collectionWe monitored the attendance patterns ofR.omeimontisat the pond during the breeding seasons from 2011-2014.The breeding period lasted 47 nights in 2011 (4/18-6/3),43 nights in 2012 (4/14-5/26),36 nights in 2013(4/22-5/27),and 42 nights in 2014 (4/12-5/23).Each night (from 7:00 p.m.to 7:00 a.m.the next morning),we searched forR.omeimontisindividuals using a 12 V flashlight.We recorded the behaviors of the frogs and sexed them based on their nuptial pads (males:presence,females:absence) and body size (males:smaller,females:larger).We marked everyR.omeimontisindividuals using color threads and recorded their nightly attendance at the pond during the breeding season.Specifically,colored threads were wrapped around the waists of frogs tight enough to prevent them slipping over the thighs without causing discomfort on frogs.We measured the snoutvent length (SVL) of each available frog three times to the nearest 0.01 mm using a Vernier caliper and then used the mean values to represent its body size.We then clipped approximately 1 mm3of tissue from the hindfoot of each individual.All the individuals were released immediately after sampling.Frogs returning to the breeding site could easily be discriminated from newcomers due to the probably maintained marks and/or the lacked clipped toe tips.Frogs that lost the thread marks were resampled through toe clipping.We collected all foam leaf nests and cultivated each clutch in a plastic bucket (with a 10-20 cm depth of water) by the side of the pond.After the hatching process finished,we counted the total number of offspring(including tadpoles,the dead embryos,and unfertilized eggs) from each clutch.We selected 24 tadpoles randomly from each clutch and took small tissue samples by tailclipping at Gosner stage 40 (Gosner,1960).Finally,all tadpoles were released back to the pond.All samples were immersed in 95% ethanol and stored at -20 °C for DNA extraction.
2.3.Genotyping of microsatellites and paternity analysesBased on the protocol of Zhaoet al.(2016),we first extracted genomic DNA from tissue specimens,and then amplified and genotyped eight polymorphic microsatellite loci to obtain the polymorphic information of sampled adults and tadpoles (Zhaoet al.,2012,2016).The COLONY program (Jones and Wang,2010) was used to determine the paternity of the sampled tadpoles.
2.4.Data analysesFirst,we assessed whether female nightly a ttenda nce was a ffected by male nightly attendance and whether the mating pattern was affected by OSR inR.omeimontis.Subsequently,we assessed whether male chorus attendance reflected male genetic quality through data across years.Incomplete data on male chorus attendance in 2011 meant that we could not include this year in the analysis.According to the high fidelity of breeding site commonly found in anuran species (Pittman and Dorcas,2009),migration inR.omeimontisis negligible.In this situation,the recapture rate could be regarded as the survival rate.Individual identification of the resampled frogs could be achieved through comparing microsatellite genotypes.We compared arrival time (the day when the male was present at the breeding site for the first time since the breeding period began),chorus attendance,mating success and body size between surviving males and non-surviving males (males absent in the breeding site in the next year)and between survived males and new males (males first present in the breeding site) by using the Wilcoxon Mann-Whitney rank sum tests (Lam and Longnecker,1983).Finally,we analyzed whether females performed random mating by evaluating the Poisson distribution of male mating success.Based on the results of paternity analyses and the total number of offspring per clutch,we calculated the number of the tadpoles sired by each candidate males for each mating group.Summing the number of the offspring sired by a male from all copulations within a breeding period,we obtained male reproductive output.The Kolmogorov-Smirnov goodness-of-fit test was used to test the distribution of male mating success (Conover,1972).We then conducted correlation analyses between mating success and male chorus attendance,male body size.Moreover,the relationship between female and male body size in each mating pair/group was also investigated to determine whether size-assortative mating occurred.All the statistical analyses were conducted using the software package R (R Core Team,2019; http://www.r-project.org/).Single variables were summarized by measurements including ranges (minimum valuesmaximum values) and arithmetic means ± SD (standard deviation).Variables used in the comparisons were first tested for normality and homogeneity variance.
3.Results
3.1.Nightly attendance and operational sex ratiosThe number of males and females present at the breeding site differed among nights and the nightly OSR was highly male biased (Table 1).Notably,significant positive correlations were found between male and female nightly attendance in all four years (Table 1).Correlation analyses revealed that the mean number of males within a spawning group per night was significantly positively correlated with the nightly OSR in three of the studied years,but not in 2014 (Figure 1; Table 1).
3.2.Male chorus attendance and male qualityThe numbers of nights that males attend the chorus at the breeding site were 36,37,36,and 42 in 2011-2014,respectively.Five sampled males returned to the breeding site in 2012,two in 2013,and five in 2014,giving return rates of 12.5%,10.0%,and 23.8%,respectively,per year.Compared with non-surviving males,there were nonsignificant trends that surviving males were present in the breeding site earlier or showed larger body size (Table 2).In contrast,the chorus attendance of surviving males was significantly higher in 2012 but not in 2013 (Table 2).Data of male chorus attendance was lacking in 2011,so there was no similar comparison analysis in 2011.Consistent with this,males that survived to successive breeding seasons showed significantly higher mating success in 2011 and 2012 but not in 2013 (Table 2).Compared with new males,survived males commonly arrived at the breeding site earlier,showed larger body size,and joined in the male chorus more frequently (Table 3).As a result,survived males showed slightly higher mating success,although the differences were only significant in 2013(Table 3).

Figure 1 The relationship between the nightly mean number of males per group and nightly operational sex ratio (OSR).
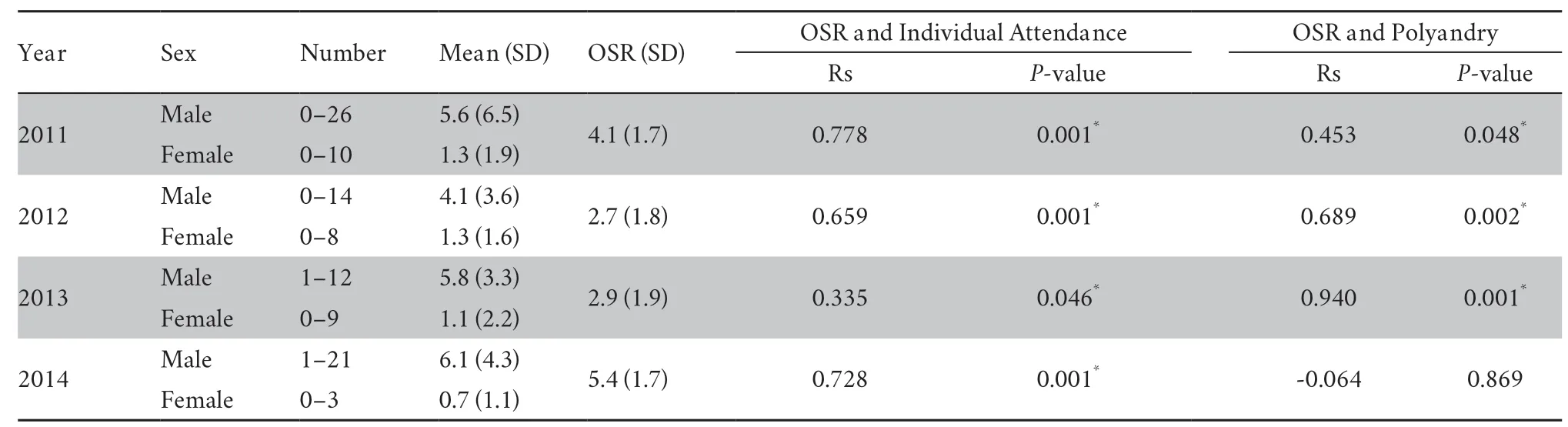
Table 1 The numbers of the two sexes present at the breeding site and the relationships between operational sex ratio (OSR) and individual nightly attendance,the extent of female polyandry.

Table 2 Comparison between males surviving and those not surviving.
3.3.Mating system and random matingDuring each breeding period,a female ofR.omeimontisonly produced one clutch but could have several mates at a time(simultaneous polyandry).In contrast,a male could be present at the breeding site on several nights and thus participated in several copulations (sequential polygyny).The number of mates per female ranged from 1 to 6 (mean± SD=2.4 ± 1.4,n=96),and for a male ranged from 0 to 8 (mean ± SD=2.1 ± 1.6,n=110).The distributions of male mating success were not significantly different from a Poisson distribution in all the four years(Kolmogorov-Smirnov tests of goodness-of-fit:2011,P=0.860; 2012,P=1.000; 2013,P=0.991; 2014,P=0.512).However,our analyses showed a significant positive correlation between male mating success and male chorus attendance (Figure 2; Table 4),suggesting that females mate randomly with males present at the breeding site more frequently.Additionally,male mating success was significantly correlated with male body size in 2013 and 2014,but not in 2011 or 2012 (Table 4),suggesting that males with larger body sizes do not always mate more frequently.The arrival time was found to be significantly correlated with body size except in 2014 for both males and females (Table 5),indicating that larger frogs arrived first at the early stage of the breeding period.However,no significant positive correlation was found betweenfemale body size and the paired male body size (Table 5),providing no evidence for size-assortative mating.

Table 3 Comparison between survived and new males.
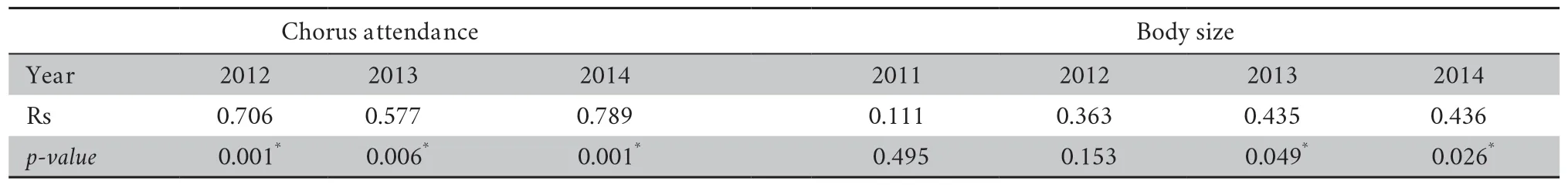
Table 4 Male mating success as a function of chorus attendance and body size.
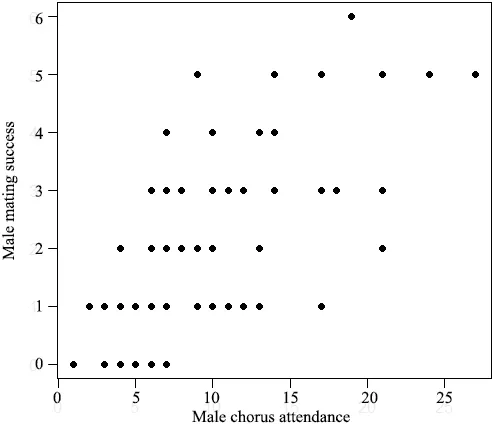
Figure 2 The relationship between male mating success and male chorus attendance.
4.Discussion
The breeding behavior ofR.omeimontisinvolves a lek mating system characterized by male aggregation and chorusing and female asynchronous arrivals (Hoglund andAlatalo,1995).Females showed simultaneous polyandry that a female can mate with more than one male in one spawning.It has been reported in many studies that a male-biased OSR was the main driver for anuran polyandry (Hase and Shimada,2014).Our analyses revealed a significant positive correlation between mean numbers of female mates per night and nightly OSR inR.omeimontis,supporting previous studies (Byrne and Roberts,2004; Hase and Shimada,2014).Nightly OSR was determined by male and female nightly attendance which were commonly correlated with each other.In our study,significant positive correlations between male and female nightly attendance were evident,consistent with previous studies (Shelly,2001; Murphy,2003).Two main hypotheses have previously been proposed to explain such correlations.The first hypothesis proposes that the nightly attendance of one sex may be directly influenced by that of the other sex (Henziet al.,1995; Murphy,2003).This is a plausible explanation of our observations of female nightly attendance because female anurans commonly arrive at the breeding site later than males.Females may decide whether they visit the breeding site after assessing the number of active males at the breeding site through choruses.The second hypothesis considers environmental conditions,such as air temperature and rainfall,as important factors affecting individual nightly attendance and chorusing activity (Brookeet al.,2000; Oseen and Wassersug,2002; Byrne and Roberts 2004).Males and females respond similarly to the same environmental conditions,thus leading to the accordance in nightly attendance between males and females (Murphy,2003).
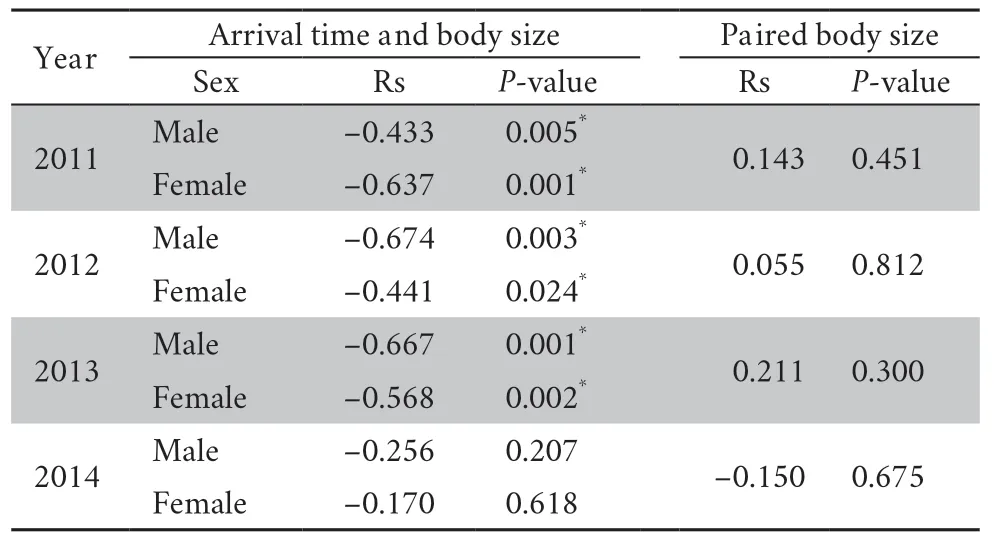
Table 5 The relationship between arrival time and body size,female body size and paired male body size.
In many anuran leks,chorus attendance has been shown to be the determinant of male mating success,regardless of whether females choose mates or mated randomly (Castellanoet al.,2009; Botto and Castellano,2016).Male chorus attendance can therefore be a good indicator of male quality.Males with higher genetic quality are at an advantage in terms of survival,which leads to higher male mating success.However,males with average genetic quality could also display traits correlating with higher-than-average mating success in short periods when taking life history strategic tradeoff between current and future fitness.In this situation,male chorus attendance just reflected male current body conditions.Thus,multiple-year data on male survival and reproduction were required to decide whether male chorus attendance reflected male genetic quality.In our study,R.omeimontismales that survived from one breeding period to another generally showed higher chorus attendance and mating success than males that did not survived.This relationship between male chorus attendance and male genetic quality was supported by another evidence:compared to the new males,survived males showed significantly earlier arrival time,larger body sizes and higher chorus attendance,and non-significantly higher mating success.Consequently,chorus attendance is related to male genetic quality inR.omeimontis.In our study,R.omeimontismale mating success was found to follow a Poisson distribution in all four years,suggesting that mating events were independent,that is,females performed random mating (Friedl and Klump,2005).In fact,females were observed in amplexus as soon as they were present at the breeding site and we found no evidence for size-assortative mating inR.omeimontis,consistent with previous studies on lek-breeding species(Bertramet al.,1996; Friedl and Klump,2005; Diaset al.,2017; Székelyet al.,2018).Additionally,male chorus attendance was the most important determinant of mating success in the Omei treefrog,similar to other lekbreeding species (Hoglund and Alatalo,1995; Castellanoet al.,2009; Jaquieryet al.,2010),male mating success in the Omei treefrog was positively correlated with chorus attendance.Females in natural anuran populations have been shown to spend a lot of time searching for and assessing males in leks to choose the fittest mates due to the complexity of the environment (Bourne,1992).However,extended mate searching might expose females to higher risk of predation (Grafe,1997).As a result,only a proportion of anuran species perform this behavior and are able to obtain benefits from mate choice despite the considerable costs involved (Ryan,1983).For the other anuran species in which benefits of female mate choice do not compensate for the costs,females spend little time searching for and assessing mates,instead commonly mating with the first males that they meet (Grafe,1997).Accordingly,in these species,chorus attendance might be relatively important for male mating success in natural populations.In our study,individuals ofR.omeimontiswere undoubtedly under predation risk at the studied breeding site becauseAmphiesma craspedogaster(a known predator of frogs) were observed on several occasions.Moreover,R.omeimontisfemales obtained from mates except sperms,they may attempt to minimize time and energy spent searching for,assessing,and choosing mates.Consistent with this assumption,females were clasped by the primarily amplectant males when they arrived at the breeding site.Consequently,R.omeimontismales with more frequent chorus attendance have higher genetic quality and more chances to meet females.Females could therefore easily obtain higher-than-average quality males even if they mated randomly to minimize the costs of mate searching and assessing.As genes conferring higher fitness from males could then be passed on to offspring(Doty and Welch,2001; Kokkoet al.,2002; Huntet al.,2004),females obtain indirect genetic benefits even through random mating.
In conclusion,we found that inR.omeimontis,female nightly attendance was affected by male nightly attendance.The correlated male and female nightly attendance then determined OSR which subsequently affected the pattern of female polyandry.Male mating success was significantly positively correlated with male chorus attendance which reflects male genetic quality.Furthermore,male mating success was not related to body size and there was no evidence for size-assortative mating under female random mating.As a result,R.omeimontisfemales could still obtain indirect benefits from random mating as they were more likely to mate with more frequently present males that had higher-thanaverage quality.Random mating appears to be a beneficial strategy forR.omeimontisfemales as they can mate with high-quality males without incurring the costs involved in searching for and assessing males.This pattern of mating,which is mainly affected by the nightly attendance of individuals while females show a ‘no choice’ strategy in mate selection,may be common in lek-breeding anurans.This could be confirmed through further studies on different species.
AcknowledgementsThis research was supported by the National Natural Science Foundation of China(No.31470442).We thank the staff of the Badagongshan National Nature Reserve for assistance in field studies.
 Asian Herpetological Research2020年4期
Asian Herpetological Research2020年4期
- Asian Herpetological Research的其它文章
- Characterization of Skin Symbiotic Bacteria of Sympatric Amphibians in Southeastern China
- Taxonomic Revision of Raorchestes menglaensis (Kou,1990) (Amphibia:Anura),with Descriptions of Two New Species from Yunnan,China
- Description of a New Species of the Asian Newt Genus Tylototriton sensu lato(Amphibia:Urodela:Salamandridae) from Southwest China
- Reevaluation of the Holotype of Odorrana schmackeri Boettger,1892(Amphibia:Anura:Ranidae) and Characterization of One Cryptic Species in O.schmackeri sensu lato through Integrative Approaches
- Toad-headed Lizard Phrynocephalus forsythii (Squamata,Agamidae) as a Potential Ring Species Inferred from Population Genetic Differentiation
- No Male Preference for Large Females in the Asian Common Toad(Duttaphrynus melanostictus):Effect of the Sex Ratio and Breeding System