
水稻不同穗型品种幼穗分化相关基因表达分析
2021-01-15 07:44金正勋王思宇张忠臣李钢夑朴钟泽
东北农业大学学报 2020年12期
金正勋,王 剑,王思宇,王 珊,张忠臣,李钢夑,朴钟泽
(1. 东北农业大学农学院,哈尔滨 150030;2. 韩国农村振兴厅农业科学院,全罗北道 全州 54874;3. 上海市农业科学院作物育种栽培研究所,上海 201403)
穗型与水稻产量和群体结构等关系密切,是水稻理想株型育种和栽培研究关注的热点问题。水稻穗型主要包括穗长、枝梗数、每穗粒数、着粒密度和粒型等[1-2],增加枝梗数和穗粒数或增大籽粒是选育超级稻新品种、获取水稻超高产重要途径之一。细胞分裂素是影响水稻穗发育和生长重要激素。研究表明,幼穗发育受激素调控,特别是与细胞分裂素代谢密切相关[3]。高温下幼穗中激素代谢紊乱,特别是细胞分裂素含量显著降低,抑制颖花形成[4-5]。OsLOGL2和OsLOGL3是细胞分裂素合成酶相关基因,OsCKX5和OsCKX9是细胞分裂素氧化酶相关基因,OsRR2、OsRR5是细胞分裂素信号转导相关基因,这些基因均与水稻幼穗分化和形成关系密切。OsCKX2 基因表达下调导致花序分生组织细胞分裂素积累,改变花序分生组织和花轴分支组织,增加生殖器官数量,提高籽粒产量[6];OsCKX3和OsCKX5缺失突变体增加花器官体积以及胚珠数量,最终增加产量[7];OsRR2 结合在细胞分裂素氧化酶基因OsCKX4启动子上直接调控该基因表达[8]。LOG 基因突变体分生组织细胞分裂能力明显降低,茎端分生组织体积明显减小,枝梗分化受到强烈抑制,且仅在每个枝梗顶部形成一朵颖花[9]。
鉴于CKX、RR、LOG 等家族基因直接或间接调控穗部性状发育和形成,且DNA 转录是结构基因表达调控重要节点,其转录速度和数量均影响关联酶活性强弱,进而调控性状表现强弱。因此,本试验选择与水稻幼穗分化相关基因和不同穗型粳稻品种,通过盆栽试验比较分析不同穗型水稻品种间穗部性状以及幼穗分化相关基因转录表达量遗传差异及相互间关系,旨在为解析水稻穗部性状形成转录调控机理和开发超高产栽培技术等提供理论依据。
1 材料与方法
1.1 材料
供试品种为多粒型晚熟品种GWS 15 和松粳9号,较多粒型晚熟品种东农7366和龙稻27号,少粒型晚熟品种GWS 14 和中熟品种龙稻5 号,其中,龙稻27 号和龙稻5 号由黑龙江省农业科学院耕作与栽培研究所提供,其他品种由研究室选育新品种。
1.2 方法
本试验于2018 年和2019 年在黑龙江省哈尔滨市香坊区东北农业大学校园盆栽场开展。试验采用盆栽种植方法,盆规格为长60 cm、宽40 cm、高40 cm,盆栽用土过筛混匀后等量装盆。4月5日播种,大钵体盘育苗,每个孔播两粒催芽籽,大棚旱育秧管理。5月15日选取长势一致秧苗插秧,每盆插2 行8 穴,每穴插2 棵苗,缓苗后定植1 棵苗,每个品种3盆,其中两盆用于调查及测定穗发育情况,另一盆用于室内考种,正常水管理。全年施氮量为纯氮105 kg·hm-2,按盆表面积折算成每盆施氮量,N∶P2O5∶K2O 比为1∶0.5∶0.8。氮肥为尿素,磷肥为磷酸二铵,钾肥为硫酸钾。施肥方法是磷肥全部作基肥;钾肥50%作基肥,50%作穗肥,氮肥50%作基肥,20%作分蘖肥,30%作穗肥。插秧前3 d 施入基肥与上层20 cm 土壤混拌均匀,灌水浸泡,秧苗开始分蘖时施入分蘖肥,穗肥在幼穗分化始期一次性施入。
1.3 取样方法
插秧后每个品种秧苗按照分蘖发生顺序依次标记分蘖等级,仅标记主茎及第1 次和第2 次分蘖。待幼穗分化开始时剥离标记的茎鞘观察幼穗分化情况,分别于幼穗长度为5、10、20、30 mm时取标记茎幼穗,迅速液氮处理后放入-80 ℃冰柜内保存备用。
1.4 基因转录表达量测定
从水稻基因库获取基因定位区域,使用Primer 5.0及NCBI内Primer BLAST功能,在基因保守区内设计高特异性Actin1内参基因和幼穗分化相关基因引物,其引物序列见表1。
选 用NOVA®Taq SYBR®Green qPCR Premix(NOVA,愚公生命科技有限公司,江苏连云港)定量试剂盒及Roche Light Cycler TM 荧光定量PCR仪,以Actin1 基因为内参基因,采用Delte-DelteCt法分析基因转录表达量。参照基因ΔCT法,依公式ΔCT=2Ct(reference)-Ct(target)计算基因相对表达量, 其中Ct(reference)-Ct(target)分别为内参基因表达量和目标基因达到设定荧光值时扩增循环次数。

表1 RT-qPCR反应引物设计Table 1 Design of RT-qPCR reaction primer
1.5 数据分析
2018 年与2019 年试验结果一致,本研究仅选用2019 年试验数据作分析,数据处理采用Excel 2007和SPSS 23.0软件,LSD法显著性测验。
2 结果与分析
2.1 不同品种间穗部性状比较及穗型分类
供试不同品种间穗部性状方差分析及多重比较结果见表2,同时根据供试6 个不同品种计算穗部性状间简单相关系数。由表2可知,供试不同品种除每穗一次枝梗数外,每穗总粒数、穗长、每穗二次枝梗数、每穗一次枝梗粒数、每穗二次枝梗粒数F值均达极显著,说明不同品种间除每穗一次枝梗数外,其他穗部性状均呈极显著遗传差异。多重比较结果表明,每穗总粒数最多品种是GWS15 和松粳9 号,其次是东农7366 和龙稻27号,而GWS 14 和龙稻5 号最少,且最多的两个品种和其他品种间及次多的两个品种和最少的两个品种间差异均显著,说明不同穗型品种间每穗总粒数遗传差异显著。据此本试验将供试6个不同品种划分为3 种穗型,即GWS 15 和松粳9 号为多粒型品种,东农7366 和龙稻27 号为较多粒型品种,GWS 14和龙稻5号为少粒型品种。
不同穗型品种每穗二次枝梗数和每穗二次枝梗粒数均是多粒型品种>较多粒型品种>少粒型品种,不同穗型品种间差异均显著。相关分析表明,每穗总粒数与每穗二次枝梗数、每穗二次枝梗粒数间相关系数分别为0.9124 和0.9552,均相关显著。说明每穗总粒数与每穗二次枝梗数、每穗二次枝梗粒数间关系密切,增加每穗二次枝梗数和每穗二次枝梗粒数有利于增加每穗总粒数,进而增加库容。由表2可见,不同穗型品种每穗一次枝梗粒数依次为龙稻5号>龙稻27号>GWS14>松粳9号>GWS 15>东农7366,少粒型品种每穗一次支梗粒数多于其他穗型品种,尤其是总粒数最少的龙稻5号均显著多于其他穗型品种,说明每穗一次支梗粒数多的穗型品种与每穗总粒数并不一致。
2.2 不同穗型品种间幼穗形成相关基因转录表达量比较
2.2.1 不同穗型品种间幼穗OsLOGL2 基因表达量变化动态比较
在幼穗生长过程中供试不同穗型品种幼穗OsLOGL2 基因mRNA 表达量比较列于表3。由表3可见,随幼穗生长,供试不同穗型品种幼穗OsLOGL2 基因mRNA 表达量均持续升高,与幼穗长度5 mm 相比,30 mm 幼穗该基因转录表达量平均上升幅度多粒型品种为88.2%、较多粒型品种为91.4%、少粒型品种为70.6%,上升幅度均较大,且少粒型品种小于多粒型和较多粒型品种。幼穗长度5、10、20、30 mm 时OsLOGL2 基因mRNA 表达量多粒型品种和少粒型品种间平均极差分别为0.482、0.921、1.177、1.293,在幼穗生长过程中OsLOGL2基因mRNA表达量多粒型品种高于较多粒型和少粒型品种,较多粒型品种又高于少粒型品种,与少粒型品种相比,多粒型和较多粒型品种平均表达量分别高1.33 倍和1.22 倍。相关分析结果表明,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与平均幼穗OsLOGL2 基因mRNA 表达量间相关系数分别为0.9920、0.9433和0.9745,均达极显著。说明不同穗型品种间幼穗生长不同时期幼穗OsLOGL2 基因mRNA 表达量遗传差异较大,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与幼穗OsLOGL2 基因mRNA 表达量关系密切,幼穗OsLOGL2 基因通过mRNA 表达量正向调控每穗颖花分化和形成,基因表达量上调有利于增加每穗颖花数。
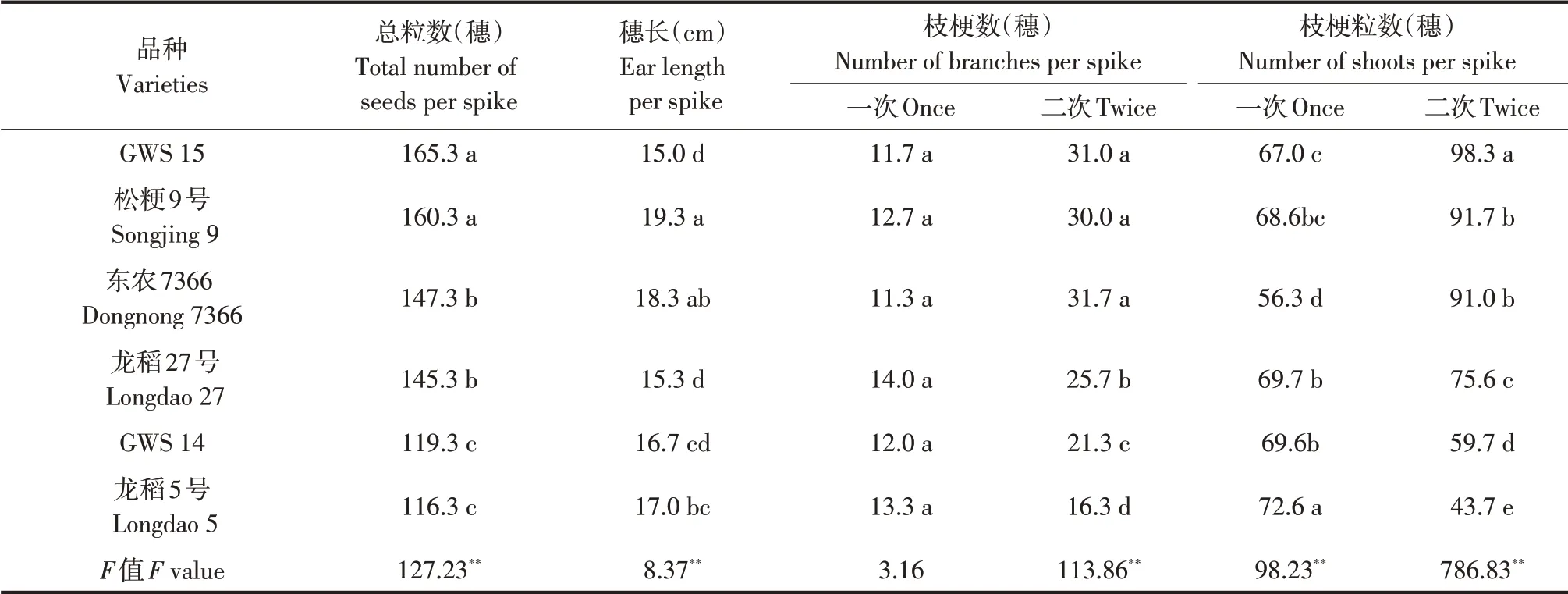
表2 不同穗型品种间穗部性状比较Table 2 Comparison of ear traits among different types of cultivars

表3 不同穗型品种间幼穗OsLOGL2基因mRNA表达量比较Table 3 Comparison of mRNA expression of OsLOGL2 gene among different panicle type varieties
2.2.2 不同穗型品种间幼穗OsLOGL3 基因表达量变化动态比较
在幼穗生长过程中供试不同穗型品种幼穗OsLOGL3 基因mRNA 表达量比较见表4。由表4 可见,随幼穗生长供试不同穗型品种幼穗OsLOGL3基因mRNA 表达量均持续上升,与幼穗长度5 mm相比,30 mm幼穗该基因转录表达量平均上升幅度多粒型品种为32.8%、较多粒型品种为29.2%、少粒型品种为30.1%,上升幅度均较大,其上升幅度多粒型品种大于较多粒型和少粒型品种。幼穗长5、10、20、30 mm时OsLOGL3基因mRNA表达量多粒型和少粒型品种间平均极差分别是0.333、0.420、0.530、0.479,幼穗生长过程中OsLOGL3 基因mRNA 表达量多粒型品种高于较多粒型品种和少粒型品种,较多粒型品种又高于少粒型品种,与少粒型品种相比,多粒型品种和较多粒型品种平均表达量分别高1.31倍和1.15倍。相关分析结果表明,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与平均幼穗OsLOGL3 基因mRNA 表达量间相关系数分别为0.9540、0.8668和0.9240,均达显著或极显著。说明不同穗型品种间幼穗生长不同时期幼穗OsLOGL3基因mRNA表达量遗传差异较大,每穗总粒数及每穗二次枝梗数和每穗二次枝梗粒数与幼穗OsLOGL3基因mRNA 表达量关系密切,幼穗OsLOGL3 基因是通过mRNA 表达量正向调控每穗颖花分化和形成,基因表达量上调有利于增加每穗颖花数。

表4 不同穗型品种间幼穗OsLOGL3基因mRNA表达量比较Table 4 Comparison of mRNA expression of OsLOGL3 gene among different panicle type varieties
2.2.3 不同穗型品种间幼穗OsCKX5 基因表达量变化动态比较
幼穗生长过程中供试不同穗型品种间幼穗OsCKX5基因mRNA表达量比较结果见表5。由表5可见,随幼穗生长,供试不同穗型品种幼穗OsCKX5基因mRNA 表达量均持续上升,与幼穗长度5 mm相比,30 mm幼穗该基因转录表达量平均上升幅度多粒型品种为32.8%、较多粒型品种为35.4%、少粒型品种为28.3%,上升幅度均较大,其上升幅度少粒型品种小于多粒型和较多粒型品种。幼穗长度5、10、20、30 mm时OsCKX5基因mRNA表达量多粒型和少粒型品种间平均极差分别为0.193、0.271、0.291、0.301,幼穗生长过程中OsCKX5基因mRNA表达量多粒型品种高于较多粒型和少粒型品种,较多粒型品种又高于少粒型品种,与少粒型品种相比,多粒型和较多粒型品种平均表达量分别高1.22 倍和1.13 倍。相关分析结果表明,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与平均幼穗OsCKX5 基因mRNA 表达量间相关系数分别为0.9680、0.9273 和0.9615,均达极显著。说明不同穗型品种间幼穗生长不同时期幼穗OsCKX5 基因mRNA表达量遗传差异较大,每穗总粒数及每穗二次枝梗数和每穗二次枝梗粒数与幼穗OsCKX5基因mRNA 表达量关系密切,幼穗OsCKX5 基因通过mRNA表达量正向调控每穗颖花分化和形成,基因表达量上调有利于增加每穗颖花数。

表5 不同穗型品种间幼穗OsCKX5基因mRNA表达量比较Table 5 Comparison of mRNA expression of OsCKX5 gene among different panicle type varieties
2.2.4 不同穗型品种间幼穗OsCKX9 基因表达量变化动态比较
幼穗生长过程中供试不同穗型品种间幼穗OsCKX9基因mRNA表达量比较结果见表6。由表6可见,随幼穗生长,供试不同穗型品种幼穗OsCKX9基因mRNA 表达量均持续升高,与幼穗长度5 mm相比,30 mm幼穗该基因转录表达量平均上升幅度多粒型品种为52.6%、较多粒型品种为65.5%、少粒型品种为59.3%,上升幅度较大,其上升幅度多粒型品种小于较多粒型和少粒型品种。幼穗长度5、10、20、30 mm时OsCKX9基因mRNA表达量多粒型品种和少粒型品种间平均极差分别为0.251、0.310、0.358、0.274,幼穗生长过程中OsCKX9 基因mRNA 表达量多粒型品种始终高于较多粒型和少粒型品种,较多粒型品种又高于少粒型品种,与少粒型品种相比,多粒型和较多粒型品种平均表达量分别高1.63倍和1.35倍。相关分析结果表明,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与平均幼穗OsCKX9 基因mRNA 表达量间相关系数分别为0.9730、0.9320 和0.9733,均达极显著。说明不同穗型品种间幼穗生长不同时期幼穗OsCKX9基因mRNA表达量遗传差异较大,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与幼穗OsCKX5基因mRNA 表达量高低关系密切,幼穗OsCKX9 基因通过mRNA 表达量正向调控每穗颖花分化和形成,基因表达量上调有利于增加每穗颖花数。
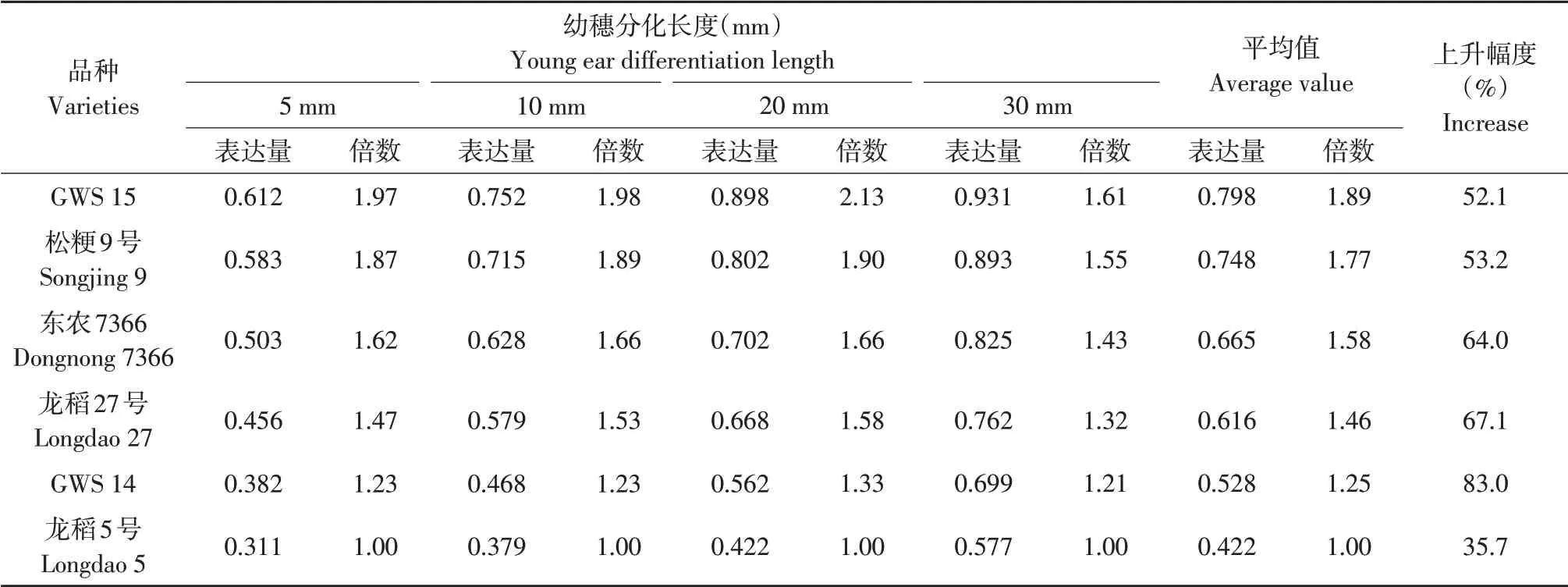
表6 不同穗型品种间幼穗OsCKX9基因mRNA表达量比较Table 6 Comparison of mRNA expression of OsCKX9 gene among different panicle type varieties
2.2.5 不同穗型品种间幼穗OsRR5基因表达量变化动态比较
幼穗生长过程中供试不同穗型品种幼穗OsRR5基因mRNA 表达量比较列于表7。由表7 可见,随幼穗生长,供试不同穗型品种幼穗OsRR5 基因mRNA 表达量均持续升高,与幼穗长度5 mm 相比,30 mm幼穗该基因转录表达量平均上升幅度多粒型品种为90.9%、较多粒型品种为81.0%、少粒型品种为78.6%,上升幅度均较大,其上升幅度多粒型品种>较多粒型品种>少粒型品种。幼穗长度5、10、20、30 mm 时OsRR5 基因mRNA 表达量多粒型和少粒型品种间平均极差分别为0.397、0.497、0.777、0.897,幼穗生长过程中OsRR5基因mRNA表达量多粒型品种始终高于较多粒型和少粒型品种,较多粒型品种又高于少粒型品种,与少粒型品种相比,多粒型和较多粒型品种平均表达量分别高1.44 倍和1.23 倍。相关分析结果表明,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与平均幼穗OsRR5基因mRNA表达量间相关系数分别为0.9720、0.8955 和0.9455,均达显著或极显著。说明不同穗型品种间幼穗生长不同时期幼穗OsRR5基因mRNA表达量遗传差异较大,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与幼穗OsRR5基因mRNA表达量关系密切,幼穗OsRR5基因通过mRNA 表达量正向调控每穗颖花分化和形成,基因表达量上调有利于增加每穗颖花数。

表7 不同穗型品种间幼穗OsRR5基因mRNA表达量比较Table 7 Comparison of mRNA expression of OsRR5 gene among different panicle type varieties
2.2.6 不同穗型品种间幼穗OsRR2 基因表达量变化动态比较
幼穗生长过程中供试不同穗型品种幼穗OsRR2基因mRNA 表达量比较见表8。由表8 可见,随幼穗生长,供试不同穗型品种幼穗OsRR2基因mRNA表达量均持续升高,与幼穗长度5 mm相比,30 mm幼穗该基因转录表达量平均上升幅度多粒型品种为45.7%、较多粒型品种为47.8%、少粒型品种为61.9%,上升幅度均较大,但少粒型品种>较多粒型品种>多粒型品种。幼穗长度5、10、20、30 mm时OsRR2 基因mRNA 表达量多粒型和少粒型品种间平均极差分别为0.360、0.379、0.494、0.444,幼穗生长过程中OsRR2 基因mRNA 表达量多粒型品种高于较多粒型品少粒型品种,较多粒型品种又高于少粒型品种,与少粒型品种相比,多粒型品种和较多粒型品种平均表达量分别高1.63 倍和1.33倍。相关分析结果表明,每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与平均幼穗OsRR2基因mRNA 表达量间相关系数分别为0.9710、0.9160和0.9602,均达显著或极显著。说明不同穗型品种间幼穗生长不同时期幼穗OsRR2基因mRNA表达量遗传差异较大,每穗总粒数及每穗二次枝梗数和每穗二次枝梗粒数与幼穗OsRR2基因mRNA表达量关系密切,幼穗OsRR2 基因通过mRNA 表达量正向调控每穗颖花分化和形成,基因表达量上调有利于增加每穗颖花数。
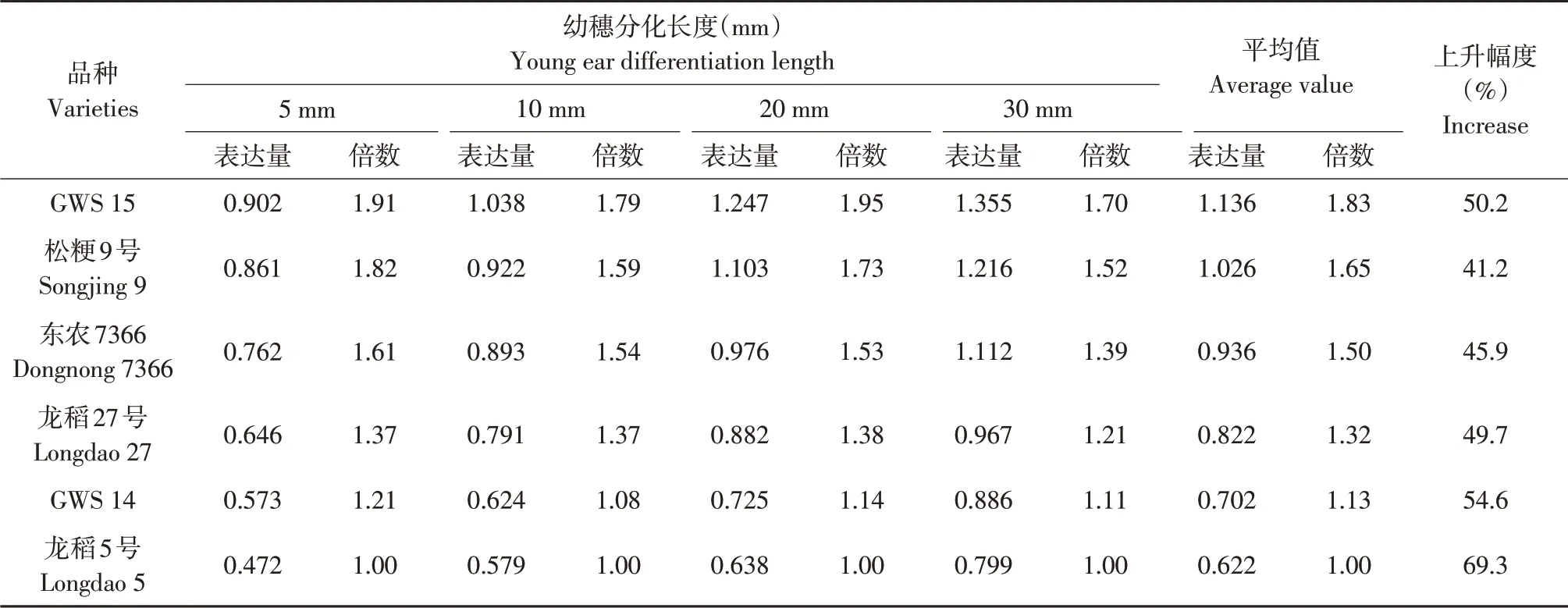
表8 不同穗型品种间幼穗OsRR2基因mRNA表达量比较Table 8 Comparison of mRNA expression of OsRR2 gene among different panicle type varieties
2.2.7 不同穗型品种间幼穗细胞分裂素合成酶和氧化酶基因表达量总和变化动态比较
幼穗生长过程中供试不同穗型品种幼穗细胞分裂素合成相关酶基因OsLOGL2、OsLOGL3和氧化酶相关基因OsCKX5、OsCKX9转录表达量总和比较结果分别列于表9和10。
由表9 可知,随幼穗生长,供试不同穗型品种幼穗细胞分裂素合成相关酶基因OsLOGL2、OsLOGL3和氧化酶相关基因OsCKX5、OsCKX9转录表达量总和均同步持续上升,上升幅度均较大,但在幼穗生长过程中幼穗细胞分裂素合成相关酶基因OsLOGL2、OsLOGL3转录表达量总和高于氧化酶相关基因OsCKX5、OsCKX9转录表达量总和,两者比值为2.32~2.91,即细胞分裂素合成相关酶基因转录表达量总和比氧化酶相关基因转录表达量总和高2倍以上(见表10),且多粒型品种高于较多粒型和少粒型品种,较多粒型品种又高于少粒型品种。说明幼穗生长过程中虽幼穗细胞分裂素合成相关酶和氧化酶相关基因转录表达量同步持续上升,但合成相关酶基因转录表达量总和远高于氧化酶相关基因转录表达量总和,幼穗细胞分裂素合成代谢优于降解代谢。
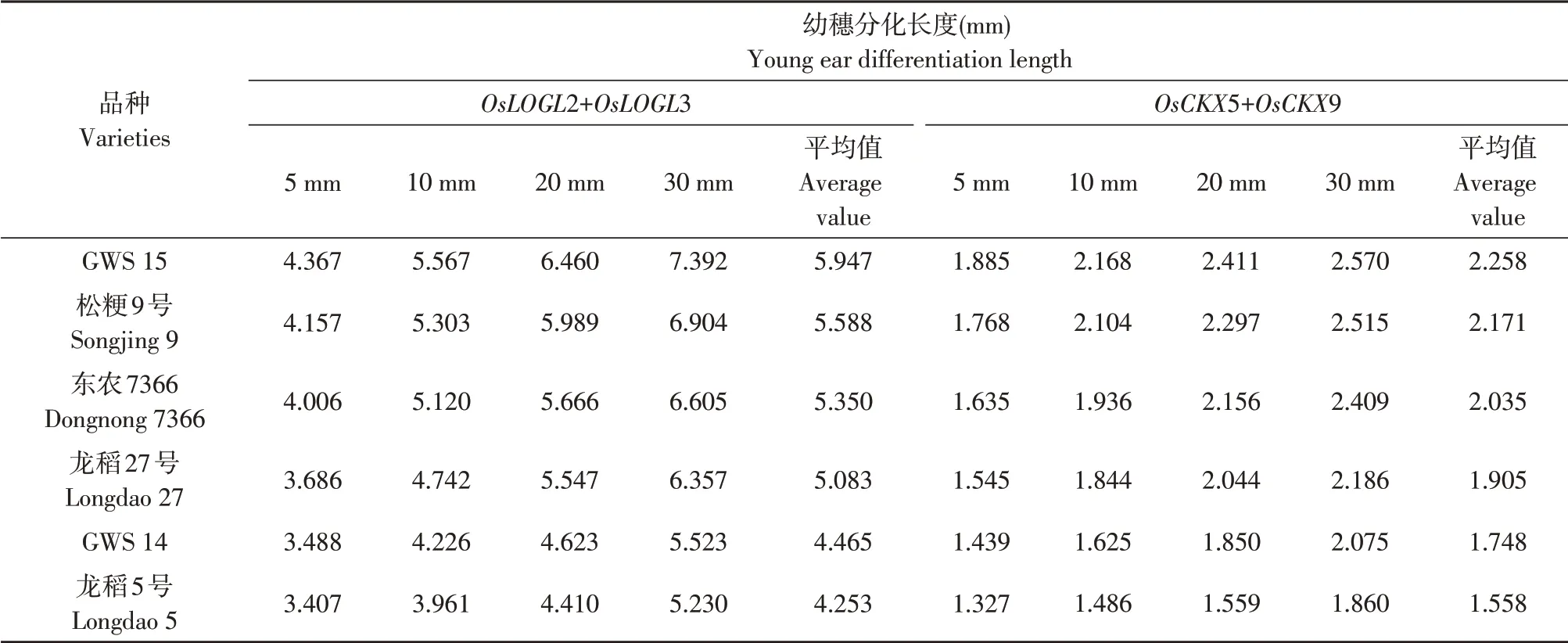
表9 不同穗型品种间幼穗OsLOGL2、OsLOGL3和OsCKX5、OsCKX9基因mRNA表达量总和比较Table 9 Comparison of mRNA expression total amount of OsLOGL2, OsLOGL3 and OsCKX5, OsCKX9 genes among different panicle type varieties
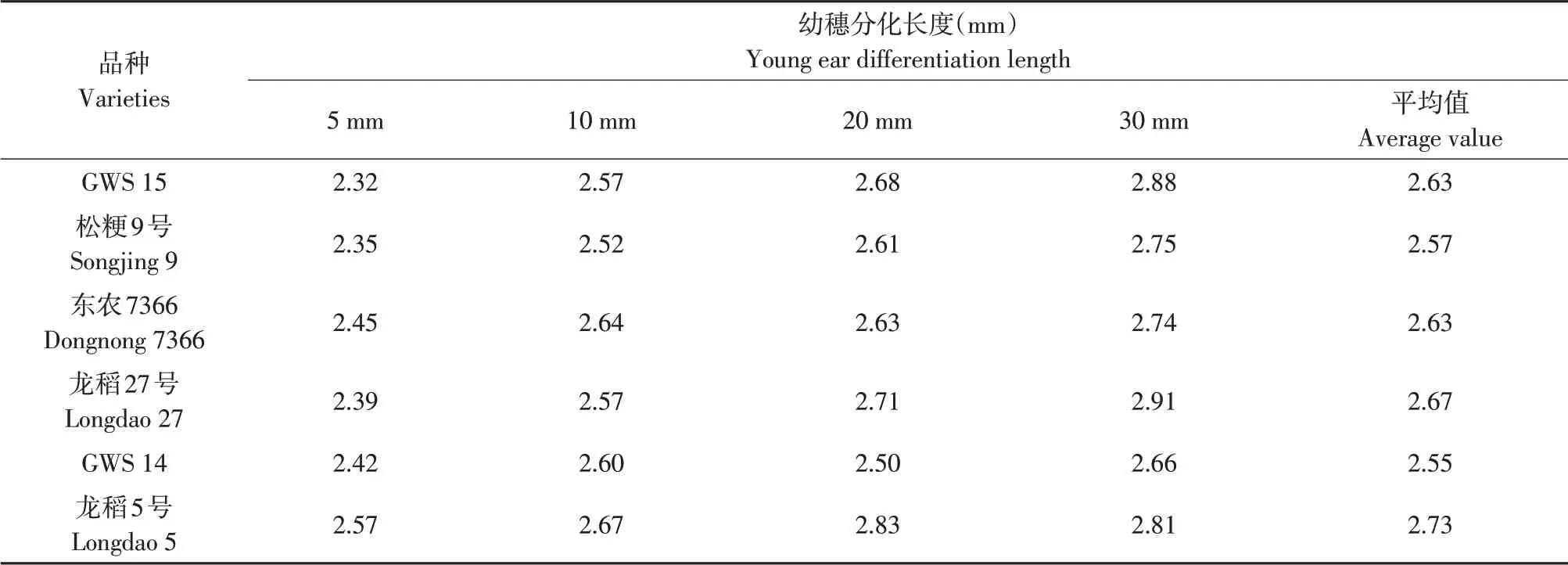
表10 不同穗型品种间幼穗OsLOGL2、OsLOGL3和OsCKX5、OsCKX9基因mRNA表达量总和比值比较Table 10 Comparison of mRNA expression total amount ratio of OsLOGL2, OsLOGL3 and OsCKX5, OsCKX9 genes among different panicle type varieties
3 讨论与结论
水稻穗型与多个穗部性状相关,在穗型分类中,由于研究目的及方向等不同,不同研究者所选取性状也有所差异,分类结果各异,如直立穗、弯曲穗、半弯曲穗、密穗、稀穗、长穗、短穗、轻穗、重穗等多种穗型。通过改良穗型和增加每穗粒数提高水稻单产已成为水稻超高产品种选育和栽培技术研究重要共识,因此深入研究穗粒数形成内外调控机制对水稻穗型塑造及提高水稻产量具有重要意义。鉴于此,本研究根据供试品种间每穗总粒数遗传差异特点,将供试品种划分为多粒型、较多粒型、少粒型3种穗型,以便阐明不同穗型形成分子调控机制。
基因转录是基因表达调控重要环节之一,其表达量不仅受自身遗传因素控制,且受氮素营养、温度、水分等外源环境因素影响[10-11]。水稻中细胞分裂素氧化酶基因OsCKX2表达缺失引起细胞分裂素在花絮分生组织中积累,导致颖花分化数目增加,有利于水稻大穗形成[12]。水稻穗型相关基因DEP1、LP 及DST 直接或间接调控OSCKX2 表达水平,调控水稻每穗穗粒数改变水稻产量[13]。Gao等研究结果表明,OsRR2结合在细胞分裂素氧化酶基因OsCKX4 启动子上直接调控该基因表达[8]。长日照条件下,Ghd7 基因表达增强,引起水稻穗型变大和穗粒数增多[14]。陈燕华等研究结果表明,高温下细胞分裂素合成有关基因OsLOGL2 和OsLOGL3表达降低,喷施0.15 mg·L-1油菜素内酯(EBR)同时促进高温和适温条件下OsLOGL2 和OsLOGL3表达[15]。与适温相比,高温抑制OsRR2、OsRR5、ORR2 和ORR4 表达,喷施0.15 mg·L-1EBR 显著提高4 个基因高温下表达量,且高于适温下表达量。由上可知,穗型相关基因表达水平不仅与基因自身结构有关,且还受其他调控基因及基因产物影响,基因表达水平变化引起与穗部性状发育和生长有关激素、营养物质等含量变化,最终引起穗部性状表型变异。由本研究结果也可知,随幼穗生长供试不同穗型品种幼穗OsLOGL2、OsLOGL3、OsCKX5、OsCKX9、OsRR2、OsRR5 基因mRNA 表达量均持续上升,其上升幅度均较大,且不同穗型品种间幼穗生长不同时期幼穗上述6 个基因mRNA表达量遗传差异较大,在幼穗生长过程中上述6个基因mRNA表达量多粒型品种高于较多粒型和少粒型品种,较多粒型品种又高于少粒型品种,且每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与幼穗上述6个基因mRNA表达量均呈显著或极显著正相关。说明每穗总粒数、每穗二次枝梗数和每穗二次枝梗粒数与幼穗OsCKX5、OsCKX9、OsRR2、 OsRR5、 OsLOGL2、 OsLOGL3 基 因mRNA表达量关系密切,上述基因通过mRNA 表达量变化正向调控每穗颖花分化和形成,基因表达量上调有利于增加每穗颖花数,穗型相关基因转录表达量变化是穗部性状发生数量变异的重要内在调控机制。且幼穗分化相关的同工型基因间转录表达量差异较大。 本试验中OsLOGL2、 OsCKX5、OsRR5 基因转录表达量均高于同工的OsLOGL3、OsCKX9、OsRR2 基因。因此,通过不同作用性质穗型相关基因聚合,调控穗型相关基因转录表达水平,增强幼穗分化和形成所必需细胞分裂素水平,以增加每穗总粒数形成强大库容,是选育水稻超高产品种重要途径。
比较本试验不同功能基因转录表达量总和可知,幼穗生长过程中虽幼穗细胞分裂素合成相关酶基因和氧化酶相关基因转录表达量均同步持续上升,但合成相关酶基因转录表达量总和远高于氧化酶相关基因转录表达量总和,说明在幼穗生长过程中细胞分裂素合成代谢和降解代谢同步发生,但合成代谢强于降解代谢,以维持幼穗生长所必需细胞分裂素浓度。所以,通过外源激素调控幼穗内源激素水平,增加每穗粒数进而形成强大库容是提高水稻单产或减轻高温灾害等重要外源调控技术途径。
猜你喜欢
烟草科技(2022年9期)2022-09-24
河南农业·综合版(2022年2期)2022-03-18
河南农业(2022年2期)2022-03-14
作物学报(2022年3期)2022-01-22
河南农业·综合版(2021年7期)2021-08-23
河南农业(2021年7期)2021-07-30
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02
山西农业科学(2021年4期)2021-04-19
江苏农业科学(2020年22期)2020-03-03
中学教学参考·理科版(2016年2期)2016-04-05
