
牦牛ACAA2基因克隆及组织表达分析
2021-01-15 07:44柴志欣武志娟王吉坤王嘉博钟金城信金伟
东北农业大学学报 2020年12期
陈 美 ,柴志欣 ,武志娟 ,王 会,王吉坤,王嘉博,钟金城 ,信金伟
(1. 青藏高原动物遗传资源保护与利用四川省、教育部重点实验室,西南民族大学,成都 610041;2.青藏高原生态畜牧业协同创新中心,西南民族大学,成都 610041;3.省部共建青稞和牦牛种质资源与遗传改良国家重点实验室,拉萨 850000)
牦牛(Bos grunniens)别名旄牛,是主要分布于青藏高原海拔3 000~5 000 m 高寒高原的草食性反刍动物,我国牦牛数量占世界牦牛总数95%,因分布区生态环境、地理分布及人文经济各异,形成不同地方品种及类群[1-2]。类乌齐牦牛主要分布于西藏自治区东北部类乌齐县[3],位于海拔4 500 m以上高山草甸草原地区,是当地优良牦牛遗传资源,以产肉为主、肉乳兼用型牦牛。麦洼牦牛主要分布在海拔3 000 m以上高山草甸草原,是肉乳兼用的优良地方品种,产肉量高,是典型利用肉质优势优良牦牛品种。牦牛肉是一种高蛋白、低脂肪、高能量的优质肉资源,牦牛肉氨基酸含量高于黄牛肉[4],是高原牧区农牧民主要食用肉类和经济来源。因此,如何利用牦牛肉质优势资源,提高牦牛产肉量及肉品质,是目前牦牛定向育种重要研究方向。
乙酰辅酶A 酰基转移酶(Acetyl-Coenzyme AAcyltr-ansferase,ACAA)包括乙酰辅酶A 酰基转移酶1(Acetyl-Co A AcyltransFerase 1,ACAA1)和乙酰辅酶A酰基转移酶2(Acetyl-coenzyme Aacyltransferase 2,ACAA2),属于酰基辅酶A代谢酶超家族中硫解酶家族[5-6],而ACAA2又称线粒体3-羰基辅酶A硫解酶,是一种参与脂肪酸代谢过程酰基转移酶,主要分布于线粒体[7]。研究显示,ACAA2是脂肪酸氧化步骤中关键酶,催化脂肪酸氧化。李恒德等通过分析ACAA2 基因多态性发现,该基因在内含子8上存在一个SNP,且与日增重和眼肌面积均显著相关[8]。李文娟采用基因芯片技术方法筛选和分析肉品质相关脂肪代谢功能基因,得出ACAA2是参与脂类代谢通路关键基因[9]。De Boer等采用基因组芯片技术分析不同小鼠脂质代谢组织表达谱发现,ZPZ 等基因存在差异表达[10]。Dio 等研究发现,脂蛋白脂肪酶(LPL)受体基因激活剂NO-1886在治疗大鼠肥胖症中发挥作用,可使ACAA2 mRNA表达水平增加1.49 倍,且LPL 活化剂NO-1886 可加速脂肪酸氧化相关酶表达,导致呼吸商(RQ)降低,为治疗大鼠肥胖提供参考[11]。因此,ACAA2基因在肥胖、脂肪酸氧化或脂肪沉积等方面均发挥调节作用。
目前,关于ACAA2 基因相关研究在绵羊等物种中均有相关报道[12],探究该基因与不同物种肉质性状相关性。本研究通过克隆获得牦牛ACAA2 基因cDNA 序列,检测该基因在不同组织中表达情况,为该基因在牦牛脂肪合成、代谢等方面调控作用提供理论依据。
1 材料与方法
1.1 材料
1.1.1 试验动物
分别选取3 头健康6 月龄麦洼牦牛(四川省阿坝自治州红原县)和类乌齐牦牛(西藏自治区类乌齐县),将其宰杀后,迅速采集其臀肌、胸肌、脾脏、肺脏、肝脏、心脏和肾脏组织,用DEPC水冲洗干净,置于液氮中保存备用。
1.1.2 试验试剂及仪器
YBR Premix Ex TaqTMⅡ和反转录试剂盒(Ta-KaRa)、 pMD19- T 载体(TaKaRa)、 DL- 2 000 Marker、Trizol(Invitrogen)、QuantStudio®3 实 时荧光定量PCR仪(Thermo)。
1.2 方法
1.2.1 总RNA 提取
采用Trizol法对所需组织进行总RNA提取,利用1.0 %琼脂糖凝胶电泳和分光光度计分别检测RNA质量、浓度及D260 nm/D280 nm值,根据反转录试剂盒说明书合成cDNA,并作为克隆和荧光定量模板。
1.2.2 引物设计及合成
根据GenBank 已公布黄牛ACAA2 基因mRNA全长序列(NM_001035342.2),利用Primer 5 设计ACAA2基因克隆引物(见表1),根据克隆所得序列设计荧光定量引物。引物由成都擎科梓熙生物技术有限公司合成。

表1 引物序列Table 1 Primer sequence
1.2.3 ACAA2基因克隆
PCR 扩 增 体 系 为15 μL: 2 × Taq Green PCR MasterMix 7.5 μL,上、下游引物各0.6 μL,模板cDNA 0.6 μL,最后加入5.7 μL ddH2O 配至15 μL。扩增条件:95 ℃预变性5 min,95 ℃变性30 s,退火温度54 ℃,72 ℃延伸90 s,72 ℃终延伸5 min,35 个循环;1.0%凝胶电泳检测其扩增产物,使用DNA 纯化试剂盒作产物DNA 纯化回收,将胶回收产物与pMD- 19T 载体16 ℃过夜连接, 转化到DH5α 感受态细胞(大肠杆菌)中37 ℃、170 r·min-1震荡培养1 h,复苏后,将其涂布在含有Amp+的LB 培养基中,37 ℃培养12 h。从平板中挑取阳性菌落,置于含Amp+的LB 培养液中37 ℃、170 r·min-1震荡培养10~12 h,将PCR 鉴定后的阳性菌液送至成都擎科梓熙生物技术有限公司测序。
1.2.4 ACAA2基因序列生物信息学分析
利用NCBI 中ORF Finder 程序分析牦牛ACAA2基因开放阅读框;ExPASy中ProtParam预测该基因的理化性质;ExPASy中Prot ScaleNetPhos分析该蛋白疏水性/亲水性;PSORTII Prediction 分析亚细胞定位;TMHMM 和SignalP 分别预测蛋白跨膜结构和信号肽;Net Phos 2.0 预测该蛋白磷酸化位点;SOPMA预测蛋白二级结构;SWISS-MODEL预测蛋白三级结构;Lasergene MegAlign 软件作物种同源性序列比对。MEGA.7软件中最大似然法(ML)构建系统进化树;Graphpad prism 5 软件绘制组织表达定量图等。
1.2.5 ACAA2基因组织表达分析
利用克隆所得ACAA2 序列,设计实时荧光定量特异性引物。以GAPDH为内参基因,cDNA为模板,QuantStudio®3实时荧光定量PCR仪检测ACAA2基因在各组织相对表达量。反应体系为10 μL:上、下游引物各0.4 μL,cDNA 模板1 μL,ddH2O 3.2 μL,SYBR Premix Ex TaqTM(2×)5 μL。 反应程序:95 ℃预变性3 min,95 ℃变性10 s,55 ℃退火30 s,72 ℃延伸30 s,共40个循环。各样本相对表达量结果用2-ΔΔCt法计算,利用SPSS 19.0 软件单因素方差分析中Duncan法分析组织表达量显著性。
2 结果与分析
2.1 PCR扩增结果
PCR 扩增以类乌齐牦牛臀肌组织cDNA 为模板,结果发现扩增产物的条带清晰单一,长度为1 357 bp,与预期扩增片段一致(见图1)。利用NCBI 中ORF Finder 作开放性阅读框分析, 得出ACAA2 基因CDS 区序列长1 101 bp,共编码366 个氨基酸残基,序列已上传至NCBI,GenBank 登录号:MT861180。
2.2 理化性质分析
在分析ACAA2 蛋白的理化性质,发现该蛋白存在Ala 和Gly 含量最多,不含Pyl 和Sec。负电荷残基总数(Asp+Glu)和正电残基总数(Arg+Lys)均为37 个。蛋白分子式为C1686H2734N486O527S18,分子质量为38 822.27 u,理论等电点pI为7.07,亲水性平均值为-0.127,脂肪指数为85.55,不稳定指数(Ⅱ)为40.88,说明该蛋白为不稳定亲水性蛋白。
2.3 亲水性/疏水性及亚细胞定位预测
在分析ACAA2蛋白疏水性/亲水性过程中,发现ACAA2 信号蛋白最小值为-2.533 ,位于193位,其亲水性最强;最大值为1.889,位于229位,其疏水性最强,说明此蛋白为亲水性蛋白。在分析牦牛ACAA2 蛋白亚细胞定位过程中,发现ACAA2蛋白在细胞质中分布最多(47.8%);其次为线粒体和细胞核(26.1%和17.4%);在内质网和细胞骨架内分布较少,均为4.3%。
2.4 跨膜区与信号肽预测
分析ACAA2 基因编码蛋白跨膜区与信号肽,发现牦牛ACAA2 蛋白即不存在跨膜区域,也无信号肽区域,说明该蛋白质即为非跨膜蛋白,也是非分泌蛋白。
2.5 蛋白磷酸化分析
预测ACAA2 蛋白磷酸化位点, 发现牦牛ACAA2 基因编码氨基酸中共发现14 个丝氨酸(Ser)、10个苏氨酸(Thr)和4个酪氨酸(Tyr)磷酸化位点。
2.6 二级和三级结构预测
预测牦牛ACAA2蛋白二级结构,发现ACAA2蛋白主要结构为α-螺旋154 个,占总链42.08%;其次为无规卷曲116 个,占总链31.69%;延伸链和β-转角分别为64 个和32 个,占总链17.49%和8.74%。三级结构预测显示,该蛋白主要以α-螺旋为主,与二级结构预测结果一致,且两者序列一致性可达88.25%。
2.7 一致性比较与系统进化树分析
将牦牛ACAA2 基因CDS 区序列与其他物种该基因CDS 序列进行同源性比对,发现牦牛ACAA2基因序列与黄牛、马序列一致性较高,分别为92.0%和88.7%;与鼠类一致性较低,与家犬一致性最低(见表2)。由图3可看出,牦牛与黄牛亲缘关系最近,与马和家犬亲缘关系最远,符合物种进化规律。
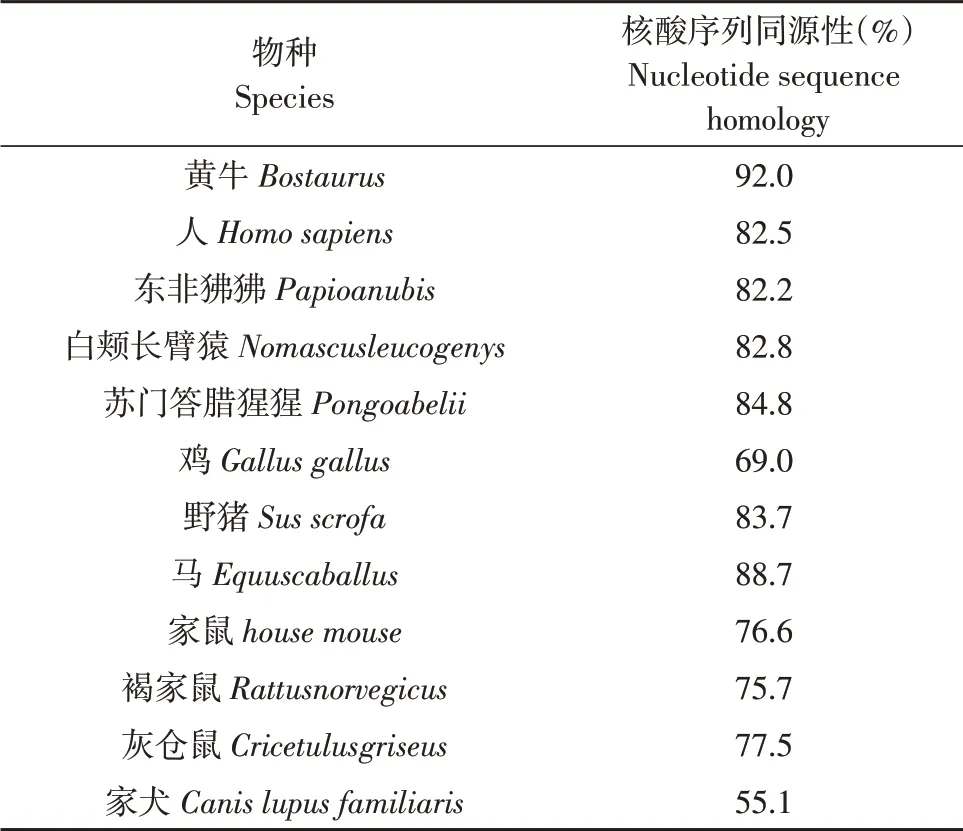
表2 各物种ACAA2基因序列的同源性比对Table 2 Homology alignment of ACAA2 gene sequence of each species
2.8 牦牛ACAA2基因组织表达分析
将麦洼牦牛心脏、肝脏、脾脏、肺脏、肾脏、胸肌和臀肌7 个组织总RNA 按照反转录说明书反转,获得cDNA,以GAPDH 基因为内参基因,利用实时荧光定量PCR 分析麦洼牦牛7 个组织差异,发现牦牛ACAA2基因在以上组织中均表达(见图4),且肝脏、肾脏与其他组织相比,差异极显著,胸肌和臀肌中表达量最低。
3 讨 论
随着人们生活水平提高,人们对肉质口感要求也越来越高,牦牛肉与黄牛肉相比,口感和嫩度相对较差,但富含丰富蛋白质、氨基酸及微量元素,且具有脂肪低、热量高等优势,深受人们喜爱。影响牛肉品质感官指标包括色泽、肌内脂肪含量及肉质柔韧性等[13],其中肌内脂肪(IMF)含量是主要影响因素[14],脂肪含量又受脂代谢调控[15],且关于IMF发育规律及脂肪沉积机制研究主要集中在肉牛[16]、鸡[17]和猪[18],在牦牛中研究相对较少。因此,如何提高牦牛肉品质和肌内脂肪含量也成为当前研究重点和热点之一。
3.1 牦牛ACAA2基因序列特性
ACAA2主要分布在线粒体中,参与体外脂肪细胞分化,是脂肪酸β-氧化途径关键酶,脂肪酸在β-氧化时产生乙酰辅酶A,在ACAA2作用下催化进入三羧酸循环彻底氧化,并释放能量,维持细胞活性,从而参与脂肪酸代谢、调节线粒体细胞信号转导、细胞凋亡信号等,为参与脂类代谢关键基因[19]。胰岛素是脂肪生成主要调节因素之一,可促进肝细胞内葡萄糖合成糖原和储存葡萄糖,但过量葡萄糖在肝脏中转化为脂肪,影响脂肪合成基因表达。王倩倩等在探讨ACAA2 基因与鹅肝脂肪代谢关系研究中发现,ACAA2基因表达与鹅肥肝形成密切相关,且在对鹅肝作不同浓度油酸、棕榈酸和胰岛素处理时,发现ACAA2 基因表达量均变化显著[20]。
本研究成功克隆牦牛ACAA2 基因CDS 区序列,分析牦牛ACAA2 基因理化性质和亚细胞定位,发现ACAA2 基因主要在细胞质中分布,细胞质是新陈代谢主要场所,也是绝大部分物质中间代谢(如醣酵解作用、氨基酸、脂肪酸和核苷酸代谢)场所,说明ACAA2蛋白在新陈代谢方面发挥作用,且与脂肪酸代谢有关。对其磷酸化分析显示,该蛋白磷酸化位点数量最多的是丝氨酸(Ser),丝氨酸可促进脂肪和脂肪酸新陈代谢,在肌肉组织合成过程中也发挥一定作用,此外,还是磷脂的主要成分之一,推测ACAA2可能调控牦牛脂肪代谢。
3.2 ACAA2基因在牦牛各组织中表达
分析ACAA2 基因在麦洼牦牛各组织中表达量,发现该基因在肾脏中表达量最高,其次是肝脏,在胸肌和臀肌中表达量最低。研究表明,研究固始鸡×安卡鸡F2资源群中胸肌IMF含量存在显著差异的两组极端样品蛋白质学,发现ACAA2 基因在试验个体对应的胸肌和肝脏中均高丰度差异表达[21]。在检测ACAA2基因是否受脂肪代谢相关因子调控中,发现脂肪酸和胰岛素均可显著影响该基因表达[20]。荧光定量结果显示,ACAA2基因在肝脏中表达量较高,推测该基因在肝脂肪代谢过程中发挥作用。肾脏具有重吸收水分、葡萄糖、蛋白质、氨基酸等作用,肾脏脂质含量与肾糖阈值(RTG)相关性研究发现,肾脏脂质沉积可能促进肾小管葡萄糖重吸收,导致尿糖排泄减少,且肾脏脂质沉积是造成RTG增高重要因素[22],而该基因在肾脏中表达量最高,推测该基因在肾脏脂质沉积中发挥调节作用。此外,心脏可为机体供应氧和各种营养物质,且心肌脂质含量可作为肥胖患者心脏病生物指标和治疗靶点[23];对心力衰竭患者心脏活检发现,心肌脂肪变性可能参与肥胖相关性心肌病变发生[24]。而本研究ACAA2在心脏和肾脏中表达量相对较高,也间接反映该基因在牦牛心脏、肾脏组织中与脂肪代谢相关性,同时ACAA2 参与线粒体能量代谢过程,为器官提供能量以维持其生理功能。
4 结 论
本试验成功获取牦牛ACAA2 基因CDS 序列,通过生物信息学分析,发现该蛋白是一种不具有信号肽和跨膜结构的不稳定亲水蛋白,蛋白结构以α-螺旋为主,主要存在于细胞质中。系统进化树和组织表达分析显示,牦牛与黄牛亲缘关系最近。与家犬亲缘关系最远,且ACAA2 基因在肾脏和肝脏组织中表达水平最高。
猜你喜欢
中老年保健(2022年3期)2022-08-24
中老年保健(2022年3期)2022-08-24
青海湖(2022年3期)2022-06-09
滇池(2022年5期)2022-04-30
汉语世界(The World of Chinese)(2022年1期)2022-02-25
老年医学研究(2021年5期)2022-01-19
散文诗(2021年22期)2022-01-12
健康博览(2021年11期)2021-12-10
现代临床医学(2021年6期)2021-11-20
中国生殖健康(2020年5期)2021-01-18
