
北部湾大风江与南流江河口红树林空间分布格局研究
2021-01-15 08:17王日明戴志军黄鹄梁喜幸黎树式胡宝清周晓妍吴天亮
海洋学报 2020年12期
王日明,戴志军,黄鹄,梁喜幸,黎树式,胡宝清,周晓妍,吴天亮
( 1. 北部湾大学 广西北部湾海岸科学与工程实验室/钦州市环境生态修复重点实验室,广西 钦州 535011;2. 华东师范大学 河口海岸学国家重点实验室,上海 200062;3. 南宁师范大学 北部湾环境演变与资源利用教育部重点实验室,广西南宁 510310)
1 引言
红树是生长在热带亚热带河口或海岸潮间带的耐盐性木本植物,在渔业、海岸保护、营养盐循环及捕获碳通量等方面具有极其重要的生态服务价值[1–2]。因全球海平面上升和高强度人类活动的作用,沿海红树林海岸遭受侵蚀,红树林面临大规模消失的风险[2]。红树林变化、红树林潮滩冲淤以及红树林生境及生态服务等已成为众多学者和相关政府关注的重大理论与社会问题[1,3–5]。较多研究针对前述问题开展了相关工作[1–6],对推动红树林生态、生理及生境和响应全球变化研究具有重要价值。与此同时,作为影响红树能否发育与成活的关键因子,盐度不仅胁迫红树组织结构,影响抗氧化酶活性,而且直接控制红树林种属地域分布特征[7–12]。故在影响红树生理与形态方面的环境因子研究中,有不少是盐度对红树林的影响研究[8–9]。譬如,受潮汐淹水造成的盐度在潮滩分布不一,红树植物在潮间带分布呈现出低滩带、中潮带和高潮带带状分布特征[3]。河口水体盐度的规律性变化成为影响红树植物沿入海河段分布的限制因子[7,10–11]。此外,一些研究指出桐花树(Aegiceras corniculatum(L)Blanco)作为先锋红树植物的一种,盐度会影响其幼苗萌发和生长的进程[12],其最适宜海水盐度为8~15[13],桐花树在低盐度下生长量最大,在中等盐度海滩可以种植桐花树[14]。然而,到目前为止一直较少有研究涉及入海河口段红树林自海向陆是如何分布以及不同因素在其间扮演的角色,这在很大程度上影响了红树林宜林地选择与国家“南红北柳”—“南红”重大生态工程的开展。基于此,本文以北部湾大风江和南流江为代表,研究北部湾入海河口红树林沿程空间分布格局,探讨外部因素对红树林的影响过程,旨在揭示区域入海河口红树林分布状态及驱动机制,在理论上补充河口段红树林发育与演化体系,在应用上为“南红”工程与生态堤防建设提供技术支撑。
2 研究区域概述
大风江和南流江同属于北部湾山溪型入海河流(图1)。该区域位于北热带季风气候区,夏季高温多雨,冬季温和少雨,雨热同期。年降水量为1 693 mm,降雨主要集中在6−9 月[15],10 月至翌年5 月为枯水期。潮汐属于不规则全日潮[16],受潮汐影响的南流江入海河口发育面积约550 km2的三角洲,主入海河段16 km,河床落差达3.34‰。河口段河岸多处修建有人工河堤,自海向陆河口段形成的潮滩由发育相对平缓到坡度较陡,呈狭长的断续带状分布。受控于周边地形,大风江河口段形成深入内陆的溺谷海湾环境。河口上游东场构建有挡潮闸,导致径流较少过闸入海,仅沿线汇流区内降水进入该区域。河岸坡度较陡,河口潮滩自海向陆由发育稀疏过渡到狭长而呈断续带状分布特征。

图1 研究区域红树林分布与采样站点Fig. 1 Mangrove distribution and survey stations of the research area
3 实验设计与研究方法
为研究两个河口段自海向陆红树林的空间分布特征及其耐盐性,实验分为室内和野外两个阶段。其中室内实验选择本地河口区优势种桐花树胚胎进行不同盐度环境的沉水测试。野外实验则选择在冬季大潮高潮期间沿河口向陆地测定不同位置水体表层盐度,同时在低潮期间调查所测位置河口潮滩红树林分布及滩地坡度等。
3.1 桐花树胚胎沉水实验
3.1.1 实验材料
(1)实验所用桐花树胚胎来自南流江河口七星岛岛尾潮滩桐花树种群。这些胚胎全部是当地生长健壮的成年树树冠外围中上部向阳面的成熟胚胎。胚胎采集时间为2019 年9 月22 日。
(2)实验所用不同盐度配比水体:不同盐度水体包括0、20、30、40、50,其中盐度为0 实验用水为静放24 h 的自来水,其他盐度用水采用从实验区内所取盐度为10.9 的天然海水加海盐与经静放24 h 的自来水调配。不同盐度水体均用Pocket PAL-ES3 盐度计标定盐度,用PZ-D-5 型液体比重天平标定比重。
3.1.2 实验设计
于 2019 年9 月23 日08:00 把从实验区采回的供试新鲜胚胎进行浸泡下沉实验。以盐度为0 的自来水浸泡为对照,分别设盐度为20、30、40、50 共4 个处理,每个盐度处理设5 个重复,每组样品为10 颗桐花树胚胎。为减少水分蒸发导致的水体盐度升高,保证桐花树胚胎所处水体盐度恒定在所设计盐度,每间隔2 d 对各盐度水体进行标定、换水。每天8:00、10:00、12:00、14:00、16:00、18:00、20:00、22:00 定 时观测和记录红树林胚胎吸水下沉数量与下沉时间。
3.2 河口实地实验
为较准确地反映河口红树林的空间分布及生境特性,本文选择在枯季径流最弱期间、大潮高潮涨平阶段进行水体采样,在大潮低平期对沿岸潮滩红树进行调查。枯季大潮高潮涨平阶段河口盐水上溯抵达位置最远,即枯季河口潮区界所在。水样采集站分别沿南流江主入海河口和大风江河口河岸自口外海滨到潮区界布设,调查取样点分别为13 个及15 个(图1)。同时利用RTK 对各个站点之间的坐标进行量测,随后基于坐标计算得到各站点之间的距离。
3.2.1 河口表层水体盐度采样设计
(1)南流江入海口河段盐度调查取样:为了取得枯水季节大潮高潮位期间涨潮水流对南流江河口段水体盐度的影响,本实验于2019 年12 月15 日05:58−10:17 测定各站点(图1)表层水体盐度。为较完整地记录高潮位盐度变化特征,于06:00 在各站点水面与滩地交界处插设标杆,标杆上每隔5 cm 贴标记,以记录站点水位上升、下降情况。涨潮时水面每上升5 cm时取一次水样,直到达到水位最高点;随后落潮时每下降5 cm 时取一次水样,连取3~5 个水样。把所取水样带回实验室经过充分搅拌后用Pocket PAL-ES3盐度计测定各水样盐度。
(2)大风江河口河段盐度调查取样:实验分别在2019 年12 月29 日和30 日涨潮至最高潮位期间驾船往返于1~15 号点,每隔5 min 取一次水样带回实验室测定河水盐度。
3.2.2 河口红树林植物分布调查取样
红树林植物沿河口向上游分布具有明显差异,为便于研究和比较,本文将河口潮间带植被定性归纳为5 类。
Ⅰ类:红树植物纯林(含多种红树植物混交林)区段,红树植物在河口段沿岸潮滩地大范围分布,极少有非红树植物混杂。
Ⅱ类:红树植物大斑块区段,红树植物在河口段沿岸潮滩地聚集组团分布形成红树植物大斑块,红树植物大斑块间有半红树植物、非红树斑块,但以红树植物斑块为主。
Ⅲ类:局部红树植物混交林区段,有成年红树植物小斑块镶嵌于其他非红树混交林中,以非红树植物为主。
Ⅳ类:有一二年生红树植物小苗区段,但未发现成年红树植物。
Ⅴ类:有红树植物胚胎区段,潮滩地发现有红树植物胚胎,但未发现红树植物小苗与成年树。
基于上述标准,实验在2019 年12 月29−30 日对南流江和大风江入海河口河段沿岸红树植物分布状况采用按站点分区段普查方法进行现场调查。其中,按上述13 个站点把南流江主入海河口21.3 km 河段从口外海滨上溯,分成12 区段,按上述15 个站点把大风江河口38.1 km 河段从海向陆上溯,分成14 个区段(图1)。由此分别统计低潮时两个河口各区段潮滩出现的红树植物的分布状况。
4 结果分析
4.1 桐花树胚胎浸水下沉对盐度胁迫的响应
所采回供实验用桐花树成熟新鲜胚胎比重为0.923,置于盐度为0 的水中,漂浮于水面,有部分悬浮于水中。随后用盐度为20、30、40、50 的人工盐水对桐花树胚胎浸泡处理,用盐度为0 的水作为对照,观测胚胎在不同的盐度水体中吸水下沉时间与数量,结果见图2。

图2 新鲜桐花树胚胎对不同盐度胁迫的响应Fig. 2 The stress response of the fresh Aegiceras corniculatum embryos to different salinity condition
图2 表明,新鲜的桐花树胚胎经不同盐度的人工海水处理,随着浸泡时间的延长出现不同的变化特征。其中,桐花树胚胎在低盐度水体中下沉速度明显快于高盐度水体(图2,表1)。具体而言,在0~10 h时段内,对照组有约25%胚胎下沉,而50 盐度处理组没有下沉;20、30、40 盐度处理组有不同程度下沉,但差异不显著。10~30 h、30~50 h 时段内,各处理组中30 盐度处理组下沉最快,50 盐度处理组下沉最慢,分别为15%、25%与9%、18%。到第50 h 时,对照组已全部下沉,对照组与各处理组表现出显著差异。到第230 h 时,20 盐度处理组已全部下沉,但30、40、50 盐度的处理组仍有约15%胚胎处于漂浮(悬浮)状态,表明桐花树胚胎在高盐度水体中漂浮(悬浮)时间更长。
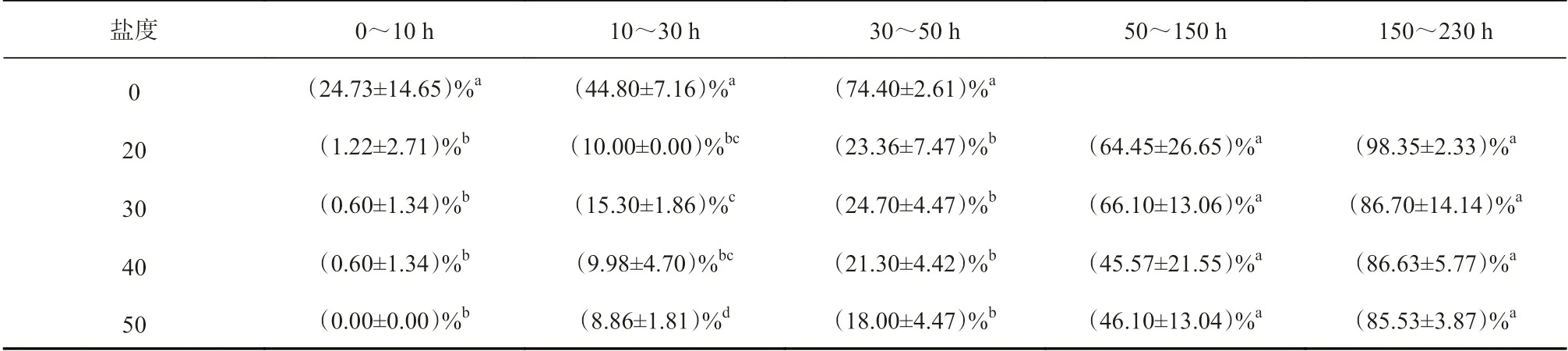
表1 不同盐度水体中新鲜桐花树胚胎在不同时段内吸水下沉差异性分析Table 1 Difference analysis for water absorption of the fresh Aegiceras corniculatum embryos in different salinity conditions during the different periods
4.2 入海河口段水体盐度空间分布
4.2.1 南流江河口段盐度空间分布特征
枯水期大潮涨平阶段,潮汐向河段内推进约15 km,其中口外海滨N01−N03 最高盐度在23 以上,盐度最高达25.8,平均盐度为23.2,潮区界左右的区段盐度平均为3.6。向海距离越远,盐度值越大,向陆距离越远,盐度值越小(图3)。
4.2.2 大风江河口段盐度空间分布特征
类似地,大风江河口口外海滨D01−D04 最高盐度在26 以上,向海距离越远,盐度值越大,向河内距离越远,盐度值越小。然而,与南流江河口有所差异,向陆到33.88 km 附近,其盐度仍高达15.8,平均盐度为10.5(D15)。有必要指出的是,D15 恰好位于大风江河流东场拦潮闸下,因闸的阻拦作用,进入闸下淡水径流极少,导致潮汐作用最远距离为33.9 km,且盐度偏高(图3)。
4.3 河口潮滩红树林植物空间分布特征
4.3.1 南流江河口潮滩地红树植物群落空间分布
南流江干流河口红树林面积为109.13 hm2(图1)。南流江主入海河段潮滩红树植物分布最集中的河段为N03−N05,沿线3.5 km 的河岸潮滩与江心洲满布红树植物秋茄(Kandelia candel(Linn.))与桐花树,另有少量的无瓣海桑(Sonneratia apetala)、海漆(Excoecaria agallochaLinn.)分布;距N03 站点口外的N01−N03沿线约4.0 km 的海滨潮滩分布有大量的互花米草与秋茄片林;距N03 站点口内3.5 km 的N05−N06 沿线1.5 km 的潮滩,桐花树生长良好,但有部分半红树植物(黄槿(Hibiscus tiliaceus)等)与非红树植物混杂;N06−N07 河段潮滩内偶布有桐花树成年苗,再入上游几乎没有成年苗;N12 河段潮滩为桐花树分布上限,只有少量的一二年生桐花树小苗分布。总而言之,南流江口潮滩表现出离河口距离越远,红树植物分布越少,长势越差的趋势(图4A)。
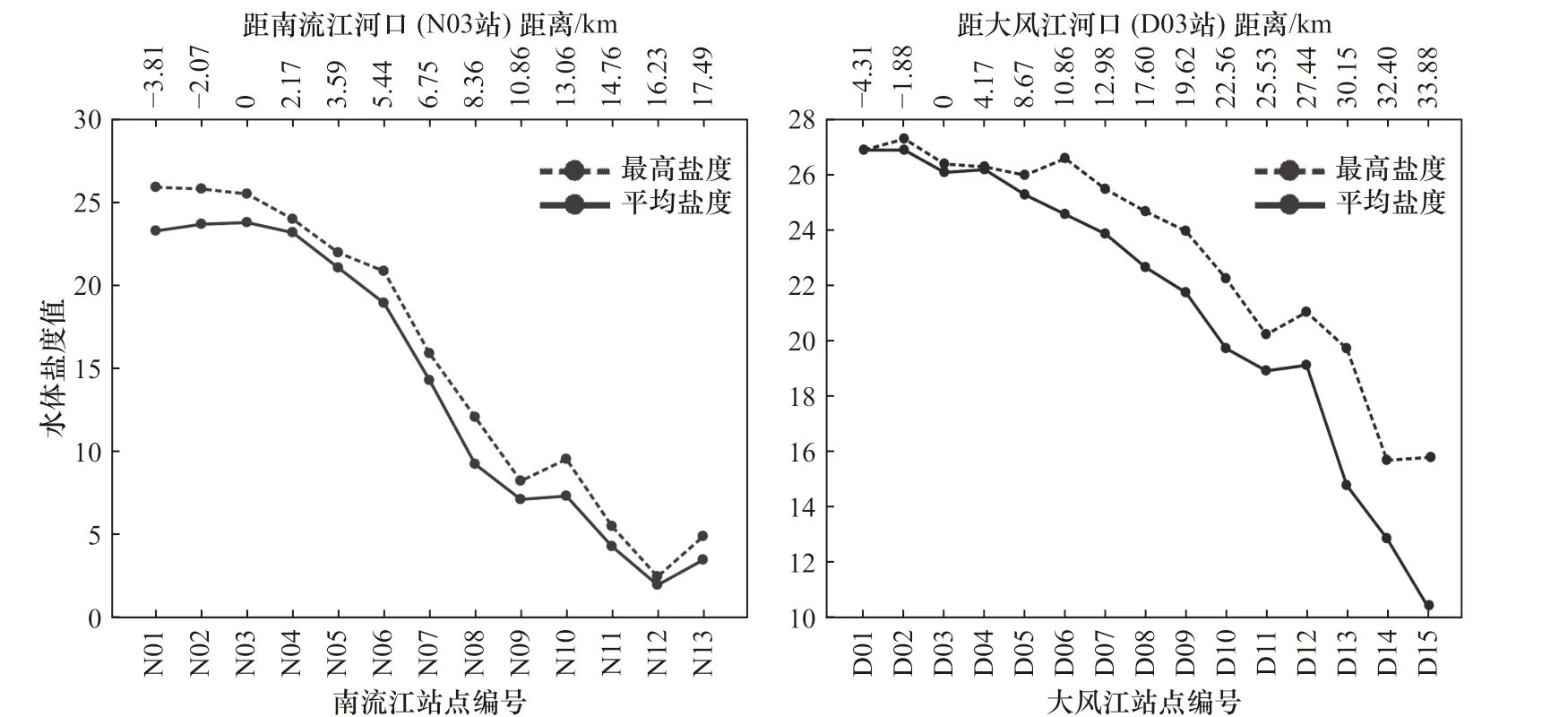
图3 观测站点盐度Fig. 3 The salinity of the observed sites
4.3.2 大风江河口潮滩地红树植物群落空间分布
大风江河口红树林面积为620.62 hm2(图1)。大风江入海河口段潮滩红树植物分布基本表现出与南流江河口相似的趋势。D03−D10 河岸潮滩与江心洲为纯红树林,其中秋茄、桐花树为绝对优势种,高潮位以上有黄槿、海漆与苦郎(Clerodendrum inerme (L.) )等半红树植物;距D03 站点口外的D01−D03 沿线约4.5 km 的海滨潮滩大多为光滩,有少量的海榄雌(Avicennia marina)与桐花树一年生苗分布;距D03 站点口内22.0 km 的D10−D12 沿线5.0 km 潮滩与江心洲皆以桐花树为优势种群,间或有半红树植物(黄槿等)与非红树植物;距D03 站点口内27 km 以上的D12−D15 桐花树成年苗三五成群镶嵌于半红树植物、非红树植物之间形成混交林(图4B)。然而,相比南流江入海口,大风江入海口红树林分布范围更广,沿河上溯距离远达距海口33.88 km 的D15 站点拦潮闸(图4B),且向陆界限主要以红树、红树幼苗及半红树混生为主,向陆界限不明显。
5 讨论
受潮汐强度、堤岸坡度、滩地高程等因素的影响及人为干扰作用,入海河口段沿岸不同潮滩地红树林植物的分布可能存在不同差异。
5.1 涨潮时间与胚胎吸水下沉时间
桐花树胚胎浸水实验表明,桐花树胚胎在低盐度的水中浸泡将吸水下沉,同时下沉的数量又和其浸泡时间相关(图2)。在相对适宜盐度的水体中,宜林潮滩桐花树胚胎成熟脱落后,可悬浮于水体且随涨潮水动力向河口内陆延伸。基于此,在非整个全日潮影响下,涨潮12 h 内,南流江河口的红树胚胎能被带到受潮汐作用影响的河段最上游N13 段(图4A)。基于先前研究关于河流受潮汐影响的上限是红树植物沿河分布的上限[17]表明,如果在该上限区有合适的潮滩,胚胎落在潮滩上可以生根发育。然而,当没有适宜潮滩时,红树胚胎在河床底着地后,将随着落潮又被河水冲刷返回下游,继续随波逐流。故南流江河口上限仅发现少量的一二年生桐花树小苗分布。红树植物分布的最上限很可能由潮汐周期涨潮上溯时长与胚胎吸水下沉时间决定。此外,在盐度为40、50 的水体中浸泡230 h,仍有部分胚胎悬浮于水体中表明,桐花树胚胎在高盐度水体中悬浮时间长,这为桐花树胚胎在落潮时随海水回流而远距离传播提供了可能。
5.2 潮滩地坡度
桐花树胚胎培育实验发现,胚胎在完全淹水条件下不进行生根发育过程。除受淹水影响,滩地也可能是限制性因子。基于调查中发现的南流江河口段N06−N08 较陡滩地极少出现桐花树胚胎与一二年生幼苗,而在N09−N12 河段缓滩发现有一二年生幼苗,在N12−N13 低潮位出露的滩涂缓滩发现有胚胎。这就表明,当桐花树胚胎被涨潮时的潮水推进到河段上游,如果胚胎停留在过陡且有潮滩的位置,落潮后胚胎并不能有效停留在滩地,而将在落潮水动力作用下带回下游。从而胚胎不能在相对较陡的滩地有效停留。在大风江河口的D10−D15 河段亦有类似情况,外缘没有适宜于胚胎停留的潮滩地,而仅在现有桐花树林下有少量一二年生幼苗出现。因而,入海河口段适宜坡度的宜林潮滩地是红树植物正常生长发育的必要条件。
5.3 水体盐度与潮水浸淹时间
桐花树胚胎在盐度为10 以下时,生根发育状况最优,桐花树对低盐度表现出很强的适应性[18]。南流江河口的现场调查表明,红树植物长势最好的河段为位于河口 的N03−N05 段,在N01−N02 红 树 长势最差。产生这一现象的原因在于:N01−N02 区域,水体盐度长期大于23,处于高盐度水平,桐花树胚胎萌发率极低[12],故红树植物极少,以光滩为主。N03−N05区域地处南流江河口径潮流交汇频繁作用的地带,全年盐度不仅在10 左右,而且发育相对较大的潮滩给红树提供了栖息场所。本研究所调查盐度为南流江河口枯水季大潮涨潮期间的水体表层盐度,故盐度相对偏高,并不能代表该河段全年平均盐度水平。
然而,大风江河口D03−D10 区段红树长势最好,所测盐度为20 以上,且由于没有河水冲淡的作用,将常年维持这一盐度水平,但红树植物依然长势最好。其主要原因在于河岸潮滩与江心洲红树植物生长范围处于中高潮位,受潮汐浸淹时间较短,而较少受到高盐度水体影响。其中低潮位与低低潮位没有红树植物生长正解释了这一现象,即长期被高盐度水体浸泡,胚胎不能萌根,从而没有小苗发育。同时,大风江河口上段在东场建有挡潮闸,正常时段闸外水位高于闸内水位,常年没有淡水补充,该河段相当于一个较深入内陆的狭长海湾,致使河段内盐度严重偏高。经现场实测,在距离海口33.88 km 的挡潮闸处D15 站点的最高盐度为15.8,平均盐度为10.5,这就导致大风江口红树林分布上限比南流江口远,分布范围比南流江口广(图1)。这也是红树植物分布沿两条河河口段分布有明显不同的主要原因。故水体盐度与潮水浸淹时间是决定河段红树种类的关键因子。
6 结论
受控于河口复杂的水文与地貌条件,河口红树林空间分布较海岸复杂。本文结合室内盐度实验,通过对北部湾大风江和南流江入海河口红树林及栖息环境的分析,主要结论包括:
(1)南流江和大风江河口红树林自海向陆都展现出距离河口越远,红树群落趋于简单和长势较差的格局:红树林纯林(或桐花树、秋茄、无瓣海桑种类混生)→红树植物与半红树植物(黄槿、苦朗等)混生→红树植物、半红树植物与非红树植物混生→红树植物镶嵌→红树林小苗。
(2)两个河口水体盐度自海向陆递减直接控制红树的分布格局,潮水浸淹时间是影响红树胚胎发育的关键因素,涨潮上溯时长决定了红树向陆生长的极限位置,河口段适宜坡度的宜林潮滩地为红树植物正常生长发育的必要条件。
(3)大风江口向陆构建拦潮闸,导致闸下盐度常年高于南流江,从而导致大风江口相较南流江河口的红树没有明显向陆界限,且主要分布红树、红树幼苗与半红树等。
猜你喜欢
小猕猴学习画刊(2022年4期)2022-05-05
老年博览·上半月(2021年10期)2021-11-02
海洋通报(2020年5期)2021-01-14
中学生百科·小文艺(2020年6期)2020-01-13
儿童故事画报(2019年12期)2019-01-07
东坡赤壁诗词(2018年3期)2018-07-16
小天使·二年级语数英综合(2018年3期)2018-06-29
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·少年版(2015年11期)2015-12-17
少儿科学周刊·少年版(2015年11期)2015-12-17
