柴归颗粒对慢性不可预知温和刺激抑郁大鼠模型肠道菌群的作用
2021-02-03 07:45赵映霞田俊生秦雪梅
中草药 2021年3期
赵映霞,许 腾,田俊生,秦雪梅
山西大学 中医药现代研究中心,山西 太原 030006
抑郁症属于情感性心境障碍,已成为严重影响患者身心健康的重大疾病,其发病率逐年增加,患病人数已超过3 亿,约占世界人口的4.4%[1]。但其病因和发病机制尚不清楚,现有抗抑郁药物的临床疗效不佳且不良反应明显,因此被列为重大疑难疾病之一。近年来,越来越多的研究报道肠道和大脑之间拥有一个包含内分泌、免疫、自主神经和肠神经等的双向通讯系统。肠道菌群与大脑之间的关系对抑郁症具有一定的影响,研究发现抑郁患者的粪便菌群组成与健康受试者存在明显差异[2];抑郁患者的粪便菌群移植到无菌小鼠可诱发抑郁样行为[3];抗抑郁药氟西汀、文拉法辛等可明显调节抑郁患者的肠道菌群组成[4-6];逍遥散可通过调节肠道菌群来改善慢性不可预知温和刺激(chronic unpredicted mild stress,CUMS)大鼠的抑郁样行为[7]。
柴归颗粒是课题组前期对宋代《太平惠民和剂局方》中的经典方剂逍遥散进行抗抑郁组分筛选和化学成分分析,通过药效学研究和临床试验观察,对原方进行化裁而研发的抗抑郁中药新药,处方由柴胡、当归、白芍、麸炒白术、炙甘草和薄荷组成。柴归颗粒的离体肠吸收特性研究发现,柴归颗粒可成倍增加甘草查耳酮B 和异甘草素在CUMS 大鼠肠道的吸收,甘草查耳酮B 和异甘草素是有效的抗抑郁活性成分[8]。柴归颗粒中含有的柴胡皂苷、白术内酯、香草酸、芍药苷、芍药新苷、芍药内酯苷、Z-丁烯基苯酞和甘草次酸均已被报道具有抗抑郁作用,可能是柴归颗粒发挥抗抑郁作用的有效成分,柴归颗粒已获得国家药物临床试验批件(2018L03149),目前正在开展药物临床试验研究。
课题组前期研究发现,柴归颗粒具有改善抑郁大鼠伴发的学习记忆能力下降、焦虑情绪和胃肠功能障碍的作用,推测柴归颗粒可能通过调节肠道菌群来改善胃肠功能和抑郁样行为。因此本研究采用16S rRNA 高通量测序技术,分析柴归颗粒对CUMS大鼠肠道菌群组成的影响,阐明柴归颗粒通过调节肠道菌群结构组成从而发挥抗抑郁作用的机制,为治疗抑郁症及抗抑郁药物的研究提供科学依据。
1 材料
1.1 动物
SPF 级雄性SD 大鼠,8 周龄,体质量180~200 g,购自北京维通利华实验动物技术有限公司,许可证号SCXK(京2016-0006)。动物于室温(22±2)℃,相对湿度(50±10)%,自然昼夜节律饲养,自由饮食和饮水。动物实验通过山西大学科学研究伦理委员会批准(批准号SXULL-2018009)。
1.2 药材
柴归颗粒(批号20181009)由柴胡16.38 kg、当归16.38 kg、白芍16.38 kg、麸炒白术16.38 kg、炙甘草8.19 kg、薄荷5.46 kg 组成,由山西省中医研究院制剂中心制备。以上药材均购于山西省华阳药业有限公司,经山西大学中医药现代研究中心秦雪梅教授鉴定分别为柴胡Bupleurum chinenseDC.、当归Angelica sinensis(Oliv.) Diels、白芍Paeonia lactifloraPall.、白术Atractylodes macrocephalaKoidz.、甘草Glycyrrhiza uralensisFisch.和薄荷Mentha haplocalyxBriq.,均符合《中国药典》2020年版标准。
1.3 药品与试剂
琼脂糖凝胶(批号75510-019)、琼脂糖凝胶电泳缓冲液(批号AM9870)、Quant-iT PicoGreen dsDNA Assay Kit(批号P7589)购自美国Invitrogen公司;Marker(批号DL2000)购自日本Takara 公司;Q5®High-Fidelity DNA 聚合酶(批号M0491L)购自美国NEB 公司。
1.4 仪器
Centrifuge TDL-5 高速离心机(上海安亭科学仪器厂);BSA124S 电子天平(赛多利斯科学仪器有限公司);大鼠旷场测试箱(长100 cm、宽100 cm、高70 cm,黑色,25 格,实验室自制);KQ-400KDE超声仪(昆山市超声仪器有限公司);NC2000 Nanodrop 紫外定量仪(美国Thermo Scientific Fisher公司);DYY-6C 电泳仪(北京六一生物科技有限公司);BG-gdsAUTO(130)凝胶成像系统(北京百晶生物技术有限公司);2720 PCR 扩增仪(美国ABI公司);FLX800T 酶标仪(美国BioTek 公司);Miseq测序仪(美国Illumina 公司)。
2 方法
2.1 动物分组与给药
大鼠适应性饲养1 周后,进行糖水偏爱实验、旷场测试和体质量基线测试,并随机分为对照组、模型组、柴归颗粒(8.3 g/kg,以生药量计)组,每组10 只。药物溶于蒸馏水,柴归颗粒组ig 药物(10 mL/kg),对照组和模型组ig 等体积蒸馏水,1 次/d,给药1 h 后造模,持续28 d。
2.2 CUMS 模型的建立
参照Willner[9]及本课题组方法[10],CUMS 模型使用9 种刺激因子,包括冰水浴、热刺激、禁食、禁水、超声刺激、夹尾、束缚、足底电击、昼夜颠倒,每天随机给予1 种应激,保证同一刺激不连续出现,持续28 d。
2.3 样本收集
给药及造模28 d 后,大鼠禁食不禁水12 h,ip 10%水合氯醛(5 mL/kg)麻醉[11],腹主动脉取血后处死,收集肠道内容物于灭菌冻存管中,液氮中速冻,随后转移至-80 ℃超低温冰箱保存备用。
2.4 16S rRNA V3~V4 区高通量测序
2.4.1 DNA 样本的提取与扩增 以行为学指标为依据,每组筛选6 只大鼠进行高通量测序。取大鼠肠道内容物200 mg,提取细菌总DNA,采用Nanodrop对DNA 进行定量分析,通过1.2%琼脂糖凝胶电泳检测DNA 纯度。扩增目的片段为V3~V4 区,上游引物序列为5’-ACTCCTACGGGAGGCAGCA-3’,下游引物序列为5’-GGACTACHVGGGTWTCTAAT-3’。PCR 条件为98 ℃预变性30 s,98 ℃变性30 s,50 ℃退火30 s,72 ℃延伸30 s,共27 个循环,72 ℃扩展延伸5 min。
2.4.2 PCR 产物定量 使用Quant-iT PicoGreen dsDNA Assay Kit 对PCR 产物在Microplate reader进行荧光定量。根据荧光定量结果,按照样本的测序量,对样本按相应比例进行混合。
2.4.3 Miseq 测序 采用Illumina 公司的TruSeq Nano DNA LT Library Prep Kit 制备测序文库,使用Miseq Reagent Kit V3 测序仪进行2×300 bp 的双端测序。利用Flash 软件(http://ccb.jhu.edu/software/FLASH/),通过质量初筛的双端序列根据重叠碱基进行配对连接,获得样本的有效序列;运用QIIME软件(http://qiime.org/)剔除疑问序列,采用Mothur软件通过Metastats 统计进行生物信息学分析。
2.5 统计分析
实验数据以±s表示,使用SPSS 22.0 软件进行数据统计分析,两组间比较采用独立样本t检验,多组间比较采用单因素方差分析(One-way ANOVA)。
3 结果
3.1 肠道菌群多样性分析
如图1-A 所示,各组间菌群操作分类单元(operational taxonomic unit,OTU)数量发生明显改变。丰度等级曲线将每个样本中的OTU 用折线连接,以此反映各样本OTU 丰度的分布规律,可直观地反映群落中高丰度和稀有OTU 的数量。折线在横轴上的长度反映了该样本中OTU 的数量,代表了群落丰度,折线的平缓程度反映了群落组成的均匀度,折线越平表示缓群落组成的均匀度越高,折线越陡峭则群落中OTU 间的丰度差异越大。如图1-B 所示,柴归颗粒组横轴上的折线较长,表明其OTU 数量较多。
α 多样性通过群落多样性指数与群落丰度指数反映样品的物种多样性,其中 Chao1 指数和Observed_species 指数表示丰度,Shannon 指数表示多样性,Pielou 指数表示均匀度。如表1 所示,与对照组比较,模型组Chao1 和Observed_species 指数显著降低(P<0.05),柴归颗粒可显著升高Chao1、Observed_species、Shannon 和Pielou 指数(P<0.05、0.01),表明柴归颗粒可有效增加肠道菌群丰度、多样性和均匀度。
β 多样性通过比较微生物群落构成来评估微生物群落间的差异。主坐标分析(principal coordinates analysis,PCoA)结果如图1-C 所示,对照组与模型组菌群结构组成差异明显,柴归颗粒组偏离模型组,接近对照组。表明CUMS 诱导大鼠肠道菌群结构组成发生变化,柴归颗粒可改善CUMS 大鼠肠道菌群结构组成和菌群多样性。
3.2 不同分类水平的肠道菌群组成分析
3.2.1 门水平肠道菌群组成 如图2 所示,从18个样品中共鉴定出厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)、栖热菌门(Thermi)、拟杆菌门(Bacteroidetes)、放线菌门(Actinobacteria)等20 个菌门,厚壁菌门和拟杆菌门的相对丰度较高,两者的比值(Firmicutes/Bacteroidetes,F/B)在一定程度上可以反映机体的健康状态。如图3 所示,与对照组比较,模型组大鼠肠道菌群栖热菌门、变形菌门、拟杆菌门和放线菌门相对丰度水平显著降低(P<0.05),F/B 值显著升高(P<0.01);与模型组比较,柴归颗粒组栖热菌门、变形菌门、拟杆菌门和放线菌门相对丰度水平显著升高(P<0.05、0.01),F/B 值显著降低(P<0.01),表明柴归颗粒可调节失调的肠道菌群水平和菌群的结构组成,从而发挥抗抑郁作用。

图1 各组大鼠肠道菌群的OTU 数量 (A)、丰度等级曲线 (B) 和PCoA (C)Fig.1 OTUs number (A),rank abundance curve (B),and PCoA (C) of gut microbiota in each group
表1 各组大鼠肠道菌群的α 多样性指数分析 ()Table 1 Alpha diversity indexes analysis of gut microbiota in each group ()

表1 各组大鼠肠道菌群的α 多样性指数分析 ()Table 1 Alpha diversity indexes analysis of gut microbiota in each group ()
与对照组比较:#P<0.05;与模型组比较:*P<0.05 **P<0.01#P < 0.05 vs control group; *P < 0.05 **P < 0.01 vs model group
组别 剂量/(g·kg-1) Chao1 指数 Observed_species 指数 Shannon 指数 Pielou 指数对照 — 260.44±99.87 266.67±90.02 3.12±0.75 0.35±0.07模型 — 131.07±17.25# 121.00±2.33# 2.23±0.36 0.28±0.02柴归颗粒 8.3 260.34±53.65** 261.20±40.80** 3.05±0.51* 0.35±0.04*
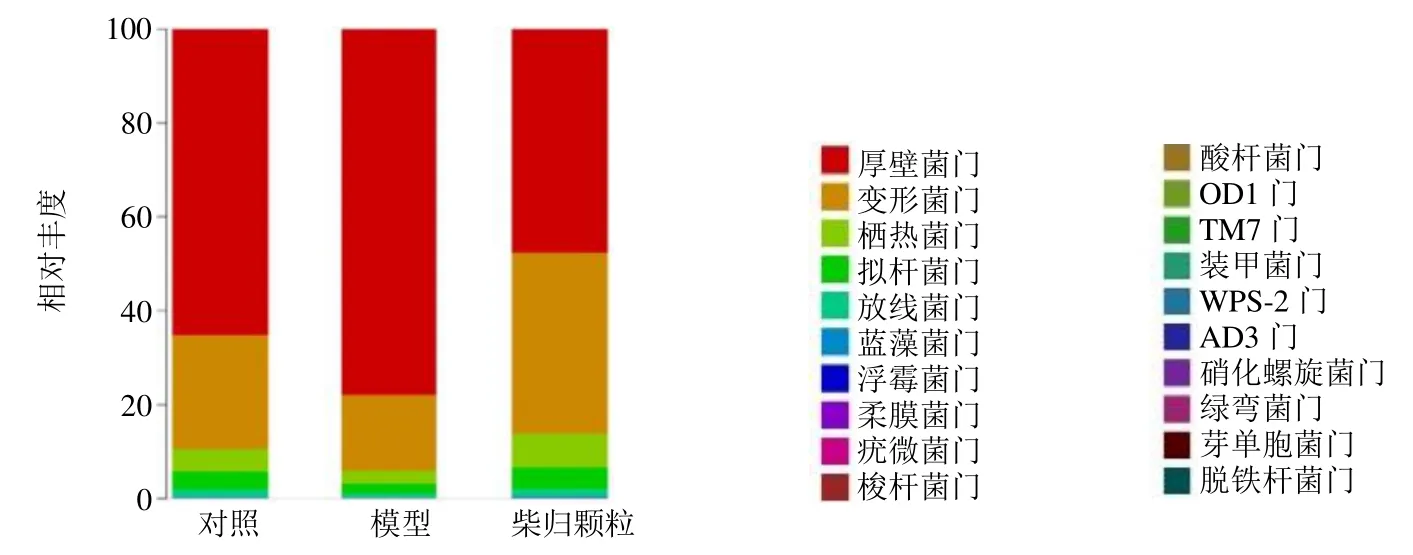
图2 各组大鼠肠道菌群在门水平的组成Fig.2 Classification and abundance of jejunum contents of phylum of gut microbiota in each group
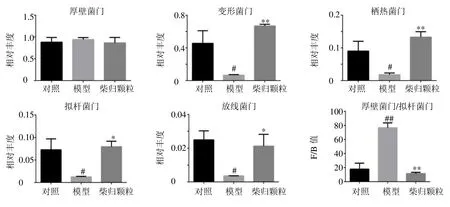
图3 各组大鼠肠道菌群在门水平的相对丰度比较Fig.3 Relative abundance of selected phylum with significant differences of gut microbiota in each group
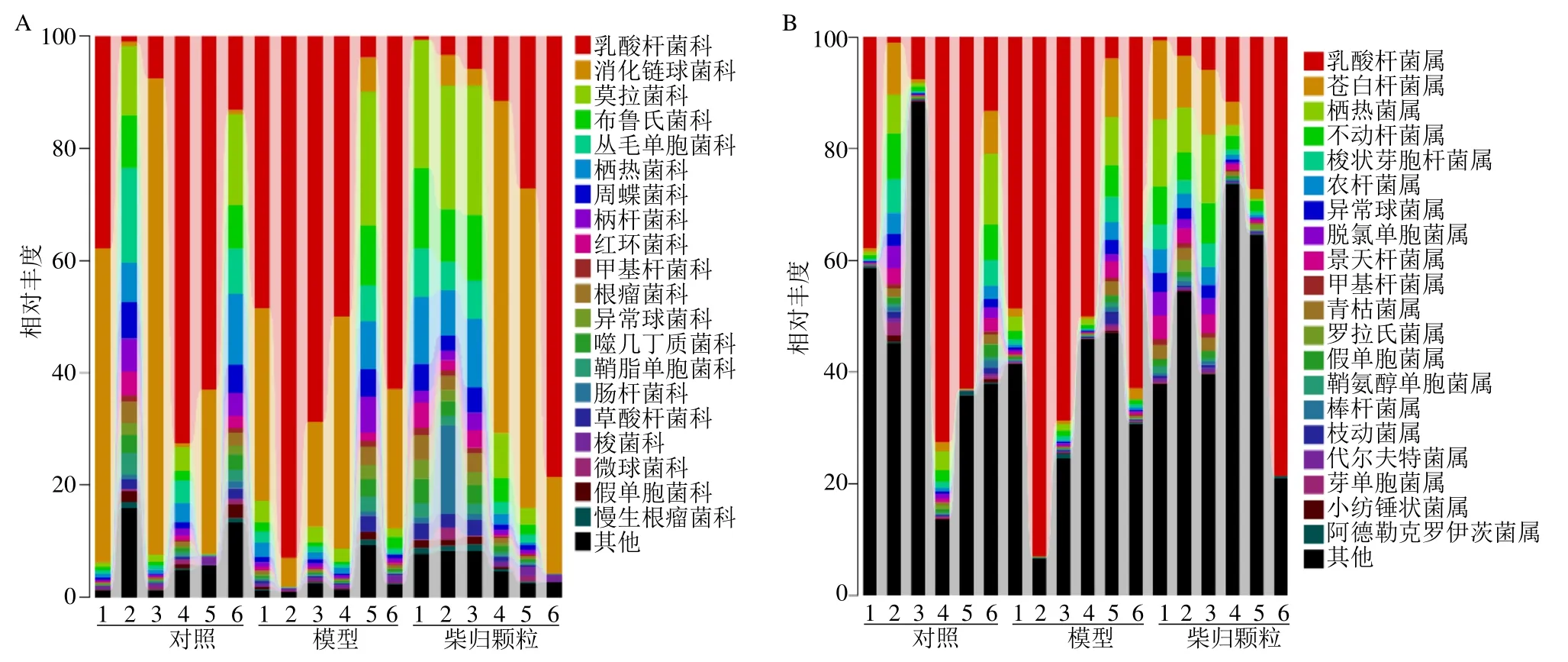
图4 各组大鼠肠道菌群在科水平 (A) 和属水平 (B) 的组成Fig.4 Classification and abundance of jejunum contents of family (A) and genus (B) of gut microbiota in each group
3.2.2 科及属水平菌群组成 从18 个样品中共得到135 个菌科和238 个菌属,其科和属水平组成见图4。如图5 所示,与对照组比较,模型组大鼠肠道菌群中布鲁氏菌科(Brucellaceae)、栖热菌科(Thermaceae)、莫拉菌科( Moraxellaceae )、甲基杆菌科(Methylobacteriaceae)和周蝶菌科(Weeksellaceae)相对丰度水平显著降低(P<0.05、0.01),梭杆菌科(Clostridiaceae)相对丰度水平显著升高(P<0.01);与模型组比较,柴归颗粒可显著回调以上菌科(P<0.01)。如图6 所示,与对照组比较,模型组大鼠肠道菌群中乳酸杆菌属Lactobacillus相对丰度水平显著升高(P<0.05),栖热菌属Thermus、不动杆菌属Acinetobacter、 甲基杆菌属Methylobacterium、青枯菌属Ralstonia、纤毛菌属Leptothrix、假单胞菌属Pseudomonas、棒状杆菌属Corynebacterium和苍白杆菌属Ochrobactrum相对丰度水平显著降低(P<0.05);与模型组比较,柴归颗粒可显著降低乳酸杆菌属相对丰度水平(P<0.01),显著升高栖热菌属、不动杆菌属、甲基杆菌属、青枯菌属、纤毛菌属和苍白杆菌属相对丰度水平(P<0.01)。表明柴归颗粒可显著调节抑郁大鼠肠道菌群,通过调节肠道菌群结构组成平衡维持机体健康。
4 讨论
本研究采用16S rRNA 高通量测序方法分析柴归颗粒对CUMS 大鼠肠道菌群的影响,从调节肠道菌群平衡的角度探讨柴归颗粒抗抑郁的作用机制。研究结果表明柴归颗粒可显著改善CUMS 大鼠肠道中乳酸杆菌属、不动杆菌属、苍白杆菌属和栖热菌属的相对丰度水平,其抗抑郁作用可能与调节上述菌群有关。课题组前期通过行为绝望模型(小鼠悬尾和强迫游泳实验)、药物诱导抑郁模型(利血平拮抗模型、5-羟色胺诱导甩头模型)和CUMS 模型对柴归颗粒的抗抑郁作用进行反复多次实验,以动物活动能力、糖水偏爱率、体质量等指标评价柴归颗粒的抗抑郁作用,发现8.3 g/kg 柴归颗粒抗抑郁效果最佳[12-13]。
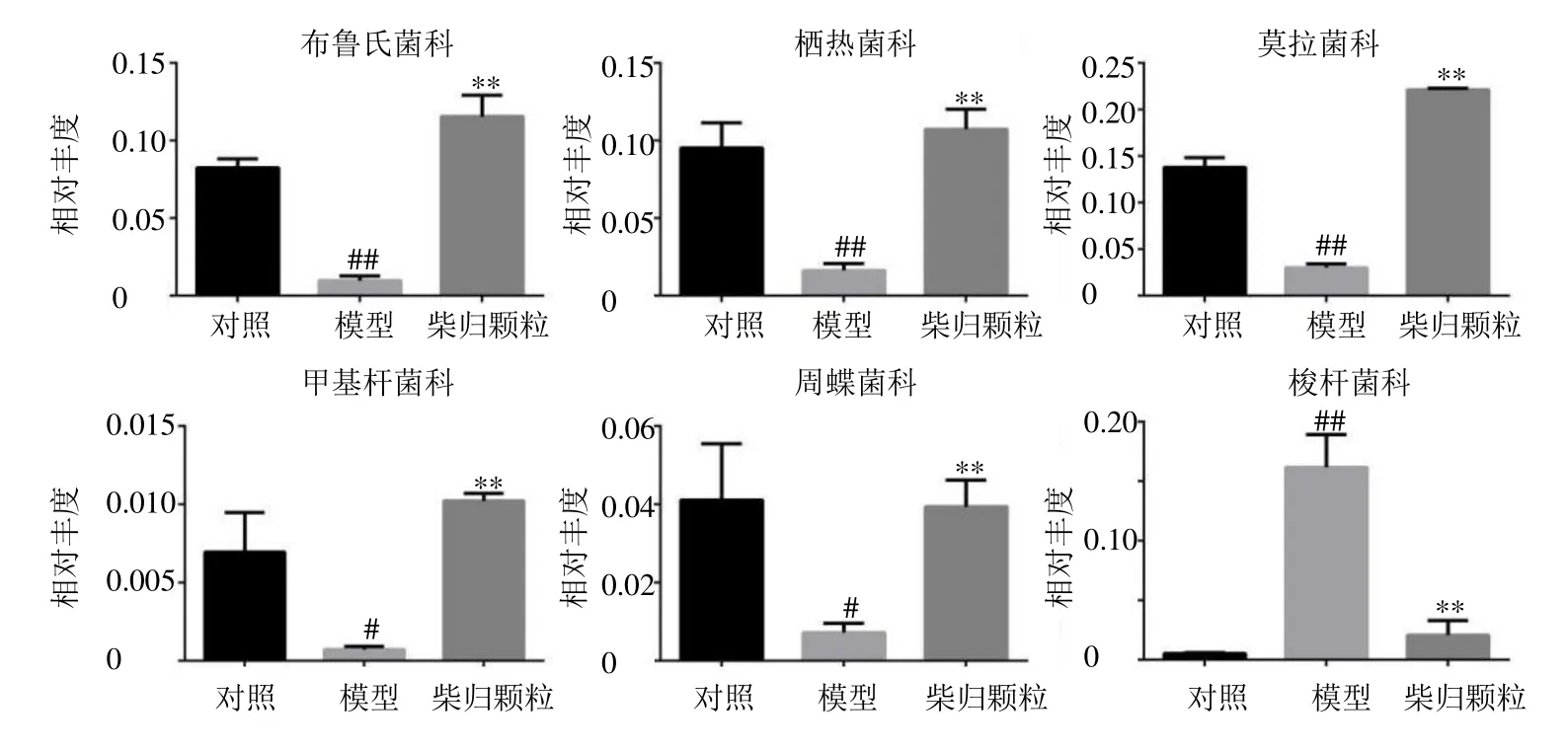
图5 科水平肠道菌群的相对丰度Fig.5 Relative abundance of selected family with significant differences of gut microbiota
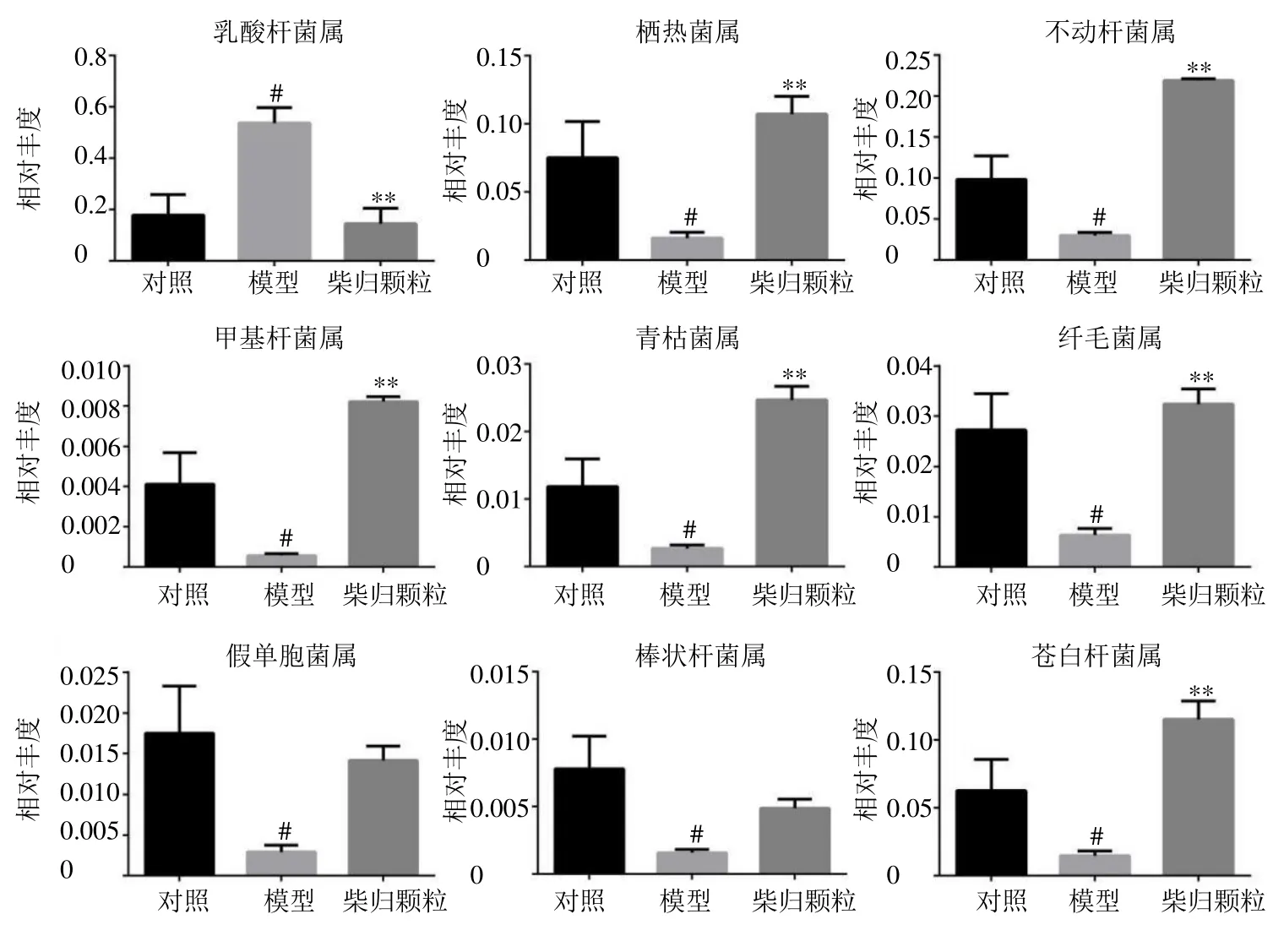
图6 属水平肠道菌群的相对丰度Fig.6 Relative abundance of selected gene with significant differences of gut microbiota
本研究发现,柴归颗粒对门分类水平的菌群具有一定的调节作用。90%肠道菌群集中在厚壁菌门和拟杆菌门,厚壁菌可以将碳水化合物代谢发酵成短链脂肪酸如丁酸盐、乙酸盐、戊酸盐等。短链脂肪酸盐诱导T 细胞分化[14],可作为具有免疫抑制[15]和抗炎[16]功能的药物;啮齿动物的临床前研究表明,抑制组蛋白脱乙酰酶能够在恐惧、焦虑、抑郁和创伤中起到增强认知的作用[17]。单胺类神经递质学说认为多巴胺能和肾上腺能神经系统功能异常是抑郁症的发病机制之一[18]。抑郁症患者脑内5-羟色胺水平降低,肠道菌群通过影响色氨酸代谢,激活吲哚胺2,3-双加氧酶,将色氨酸转化为犬尿氨酸,最终耗竭色氨酸,使5-羟色胺水平降低,从而引发抑郁。益生菌可降低吲哚胺2,3-双加氧酶活性,通过增加5-羟色胺前体色氨酸来增加5-羟色胺水平,从而改善抑郁[19]。课题组前期研究表明,柴归颗粒可上调血清中神经递质(5-羟色胺、去甲肾上腺素、5-羟吲哚乙酸)水平[20]。Aizawa 等[21]发现抑郁症患者和健康人群在肠道菌群的结构上存在明显差异;前期研究也发现,对照组和模型组大鼠的肠道菌群存在显著差异[22]。芍药苷可在粪肠球菌、金黄色葡萄球菌和短双歧杆菌的作用下转化为苯甲酸,从而发挥抗抑郁作用,表明肠道菌群与抗抑郁药物关系密切[23]。本研究结果显示,柴归颗粒可显著调节CUMS 大鼠肠道菌群的结构组成,表明柴归颗粒可能通过调节肠道菌群结构组成来发挥抗抑郁作用。
在属分类水平,柴归颗粒可降低CUMS 大鼠肠道中乳酸杆菌属相对丰度水平。乳酸杆菌作为益生菌具有较强的代谢机体内碳水化合物产酸的能力,可合成葡聚糖和杂多糖、抵抗肠道病原体的定殖、竞争性排斥病原体、抑制和消除条件致病菌、调节肠道菌群失衡[24-26]。γ-氨基丁酸(gammaaminobutyric acid,GABA)可由乳酸杆菌分泌,GABA 是存在于情感、行为调节脑区的物质,GABA受体受损导致GABA 能信号传递失衡,海马神经元受到抑制,从而产生抑郁[27]。乳酸杆菌可通过迷走神经调节神经递质(5-羟色胺和多巴胺)受体的表达来改善抑郁行为[28]。前期研究发现,健康人与抑郁患者的代谢存在差异,抑郁患者体内乳酸堆积,其可能原因是抑郁患者体内可产生乳酸的菌群增多,菌群平衡被破坏,进而诱发抑郁[29]。本研究发现抑郁大鼠中产生乳酸的乳酸杆菌水平显著增加,与课题组前期研究一致。
本研究发现,柴归颗粒能够回调抑郁大鼠肠道中不动杆菌属、苍白杆菌属和栖热菌属水平,这些差异菌属均参与了精氨酸的代谢途径。研究发现,CUMS 抑郁大鼠海马中精氨酸水平升高[30],而抑郁症患者血浆中精氨酸水平降低[31],Ozden 等[32]提出精氨酸代谢物与抑郁症的病理和生理有关,精氨酸代谢物腐胺水平可能是预测抑郁严重程度的生物标志物。Ikeda 等[33]发现腐胺的产生受肠道菌群影响,双歧杆菌可产生腐胺,动物肠道菌群中具有将精氨酸合成腐胺和亚精胺的酶。Nakamura 等[34]发现肠道内的腐胺通过多种代谢途径产生,主要依赖于肠道菌群,表明肠道菌群在精氨酸代谢过程中发挥作用。研究发现,胍丁胺作为腐胺的前体物质通过阻断抑郁动物模型中N-甲基-D-天冬氨酸受体起到快速抗抑郁作用[35-36],提示精氨酸代谢物在一定程度上可能具有抗抑郁作用。精氨酸在降解过程中涉及多种酶,肠道菌群主要通过精氨酸脱羧酶合成多胺,且多胺主要被小肠吸收[37]。多胺水平主要受限速酶调控,亚精胺/精胺N1-乙酰基转移酶是参与多胺分解代谢的关键酶,在抑郁症患者脑中其表达降低[38],提示抑郁的发生可能与多胺系统有关。慢性应激导致胞质多胺增多,促进兴奋,而长时间高浓度的胞质多胺使神经元过度兴奋,产生神经毒性[39]。课题组前期研究发现[40],逍遥散低级性部位可明显降低CUMS 大鼠亚精胺水平,而亚精胺是精氨酸的代谢产物,多胺中的腐胺、精胺、亚精胺、胍丁胺等均为精氨酸的降解产物,提示精氨酸降解途径在抑郁症中扮演重要的角色。本研究发现的不动杆菌属参与精氨酸途径,其柴归颗粒可显著调节该菌群的结构组成,提示柴归颗粒发挥抗抑郁作用可能与参与精氨酸途径的不动杆菌属有关。
综上所述,柴归颗粒的抗抑郁作用可能与精氨酸代谢途径及其相关的菌群有关,课题组后续将通过分子生物学方法对差异菌群和精氨酸降解途径相关限速酶进行验证。本研究有助于探讨柴归颗粒的抗抑郁作用及机制,为柴归颗粒在临床上的应用提供理论依据。
利益冲突所有作者均声明不存在利益冲突
猜你喜欢
中老年保健(2022年2期)2022-08-24
中老年保健(2021年5期)2021-12-02
科学(2020年4期)2020-11-26
天然产物研究与开发(2019年1期)2019-03-01
中成药(2018年4期)2018-04-26
中成药(2018年1期)2018-02-02
中外医疗(2015年11期)2016-01-04
动物营养学报(2015年10期)2015-12-01
中国当代医药(2015年30期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06