
优质水稻新种质ZY56的创制及评价
2021-03-25 07:00邱东峰葛平娟刘刚杨金松陈建国张再君
中国农业科学 2021年6期
邱东峰,葛平娟,刘刚,杨金松,陈建国,张再君
优质水稻新种质ZY56的创制及评价

1湖北省农业科学院粮食作物研究所/粮食作物种质创新与遗传改良湖北省重点实验室,武汉 430064;2湖北大学生命科学学院,武汉 430062
【】为避免未知基因和遗传背景等不可控因素对育种实践的影响,将高通量SNP分型技术与传统育种相结合,培育新品种,以提高育种效率,实现育种方法的改进,为优质食味稻米新种质的鉴定和创制方法提供参考。以鉴真2号和鄂中5号为亲本,通过杂交、回交及系谱法选育,在BC1F2—BC1F4群体开展稻米品质分析评价,至BC1F7初步选定优良株系4W1-056,进一步利用捕获测序的方法,对鉴真2号、鄂中5号及66个4W1-056单株中的68个DNA片段进行PCR扩增,并测序,分析908个SNP位点。基于SNP位点的基因型,采用p-distance方法,使用MEGA7软件进行系统进化分析。结合系统进化分析、农艺性状和稻米品质的鉴定,对新品系的选育和鉴定进行评价。经杂交、回交和自交获得株型好、分蘖力强、茎秆粗壮、抗倒性好、外观品质优的稳定优良株系4W1-056。系统进化分析表明,从4W1-056优良株系中筛选的66个单株分成3个类群:类群Ⅰ、类群Ⅱ和类群Ⅲ。3个类群之间的碱基替代率为0.1398,类群Ⅰ和类群Ⅱ之间的碱基替代率为0.0662,而在各类群内的碱基替代率为0,表明同一类群内的单株没有遗传差异。结合农艺性状和稻米品质的鉴定,将类群Ⅱ作为新的品系,命名为ZY56,与4W1-056相比,其外观品质更好,垩白度为0.9,更接近鄂中5号。利用水稻8K SNP芯片检测ZY56及2个亲本(鉴真2号和鄂中5号),显示ZY56有14.13%的染色体片段来源于鉴真2号,85.87%的染色体片段来源于鄂中5号,进一步说明ZY56的主要基因源于鄂中5号,验证了杂交、回交和自交后的选择结果。特征特性研究结果显示,ZY56完成基本营养生长所需要的最低有效积温为760.5℃,生殖生长的临界光照长度为14 h 13 min,完成幼穗分化需要的有效积温高于711.5℃。不同播期的品质分析结果表明,ZY56对光照长度的反应明显弱于鄂中5号,但生长平稳,有利于稻米品质的形成,表明ZY56的稻米品质具有更好的稳定性。在资源创制高代选择中,利用高通量SNP分型技术进行遗传一致性筛选,将通过农艺性状测定等常规方法难以细分的材料进行类群细分,最终确定符合目标要求的株系。该方法克服了传统系谱法在高世代选择中针对农艺性状难以继续选择的困难,避免了同类单株重复选择和不同类单株漏选的问题,降低了高世代选择工作量,提高了选择效率,具有推广价值。
水稻;种质创新;资源评价
0 引言
【研究意义】水稻是重要的粮食作物,为世界人口提供了约25%的能量[1]。从人类最早驯化野生稻到现在的水稻遗传改良,稻种资源的鉴定评价、创新和利用以及育种方法的改进等,始终是种质资源研究的重要内容,也是水稻品种培育的基础工作[2]。在现代生物技术条件下,如何利用分子技术评价资源,如何更高效地创制新材料将是未来水稻资源创制和遗传改良的重要研究方向。【前人研究进展】经典的种质创新方法包括有性杂交和回交、辐射和化学诱变、花培、组培、原生质融合和基因转移等[3]。综合多种水稻品种培育方法,通常包含有利基因的聚合和优异基因型的纯合2个过程。基于形态学,以经验为主的传统育种进程慢、效率低,已不能满足日益增长的粮食需求[4]。近年来,随着水稻功能基因组学领域的快速发展[5-7],水稻育种技术已逐步实践精准和高效的分子设计育种理念。分子设计育种以探究尽可能多的重要农艺性状形成的分子机理为基础,以性状优良的品种为素材进行设计,针对目标性状进行精准选择和改良,提高育种的精准程度。分子设计育种依赖于遗传学、分子生物学和基因组学等学科的发展。在当前阶段,虽有较多研究从分子水平上阐明或部分阐明了水稻产量相关[8-10]、品质相关[11-12]、耐逆性[13-16]、氮素高效[17-18]等复杂性状形成的遗传机理,但精准育种需要对这些遗传调控网络进行系统解析和组装,难度成指数级增加。因此,引入基因组学研究成果虽然能一定程度上提高了研究效率,但基因数量庞大、遗传累赘、无法准确预测基因互作和遗传背景的影响,对水稻种质资源创新利用工作提出了新的挑战。【本研究切入点】高通量SNP分型技术的发展,推动了作物遗传、种质鉴定、分子育种等方面的研究[19],然而,结合高通量SNP技术和传统育种技术,评价遗传背景和未知基因的影响,提高育种效率的研究仍鲜见报道。稻米的蒸煮与食味品质是一个综合性状,是由众多淀粉合成与调控相关基因共同控制的复杂网络,又是优质稻米重要和较难确定的性状指标[20]。尽管对控制稻米蒸煮与食味品质性状的基因功能研究得比较透彻,但是目前仍然不能有效利用现有的基因资源实现精准和高效创制新的优质食味稻米种质。同时,稻米品质的形成还受到气候、土壤等环境因素的影响[21],因此,创制优质稻品种还需要对其进行精准鉴定评价,并研究其优质食味品质形成的遗传因素与环境控制因素,才能适应现代稻米产业化生产、加工的需求。【拟解决的关键问题】本研究利用湖北省现有的高档优质水稻资源鄂中5号、鉴真2号[22-24]创制新的种质ZY56,探讨其作为优质稻的生产适应性,提出将高通量SNP分型技术与传统育种技术相结合,以提高育种效率的新育种方法,使育种由经验选择转变为针对目标性状进行精准选择和改良,提高育种效率,为优质食味稻米新种质的鉴定和创制方法提供参考。
1 材料与方法
1.1 供试材料
鄂中5号(鄂审稻2004010)[22-24]和鉴真2号(鄂审稻2001004)[24]由国家农作物种质资源库湖北分库提供,均为中籼迟熟常规品种,感温性强,感光性弱,基本营养生长期长,稻米品质优良。
1.2 材料创制方法
2011年夏季在湖北,以鄂中5号为母本、鉴真2号为父本进行杂交,F1与鄂中5号回交得BC1F1,然后连续自交,至2014年冬季,获得相对稳定的株系4W1-056。
1.3 基因型检测方法
采用武汉基诺赛克科技有限公司提供的捕获测序SNP分型技术进行基因型检测,首先分别提取4W1-056株系中的66个中选单株的DNA,设计目标位点引物,通过PCR捕获目标DNA片段,然后依据Illumina文库构建流程完成Paired-end(PE)测序文库的构建,并利用Illumina Hiseq测序仪进行测序,最后进行目标位点的基因型分析。
水稻8K SNP芯片检测由中玉金标记(北京)生物技术有限公司完成。
1.4 统计方法
采用p-distance方法[25]分析遗传距离,采用MEGA7软件[26]完成进化分析。采用EXCEL完成方差分析。
1.5 田间试验及农艺性状记载
2017—2018年在湖北武汉、荆州、武穴、枣阳以及海南陵水等地种植ZY56,调查播种期、幼穗分化期、始穗期等的农艺性状,结合当地温度等气象数据(http://www.tianqihoubao.com/lishi/),综合分析其感光性、感温性和基本营养生长性,成熟后,每小区各取10株考种并用于分析稻米品质。
2018年5月18日、5月30日、6月12日在湖北武穴分期播种ZY56和鄂中5号,收获稻谷进行稻米品质分析。
1.6 稻米品质及快速黏度仪(Rapid Visco Analyzer,RVA)谱特征值分析
参照优质稻谷GB/T17891-2017的品质分析方法,由农业农村部食品质量监督检验测试中心(武汉)分析ZY56和鄂中5号的稻米品质。由农业农村部稻米及制品质量监督检验测试中心(杭州)分析黏度RVA谱特征值。
2 结果
2.1 亲本特性及遗传差异分析
通过分析亲本鄂中5号和鉴真2号的主要性状(表1)。发现二者的农艺性状和稻米品质指标差异较大。鉴真2号单株成穗12.6个,比鄂中5号多2.5个,表明其分蘖能力强于鄂中5号;鉴真2号株高104.2 cm,但茎秆较纤细柔软,鄂中5号株高117.9 cm,茎秆粗壮,但茎基部韧性不足,易发生倒伏;鉴真2号单株产量为34.5 g,高于鄂中5号,表明鉴真2号具有更高的产量潜力。鄂中5号稻米长宽比为3.6,无垩白,外观品质优于鉴真2号。2个品种的播始历期均较长,有利于优良稻米品质的形成,生产实践表明,二者均具有稻米品质优良的特点,但鄂中5号稻米蒸煮后的蓬松度较鉴真2号差。结合遗传相似系数与农艺性状差异考虑,选用鄂中5号和鉴真2号为亲本杂交再回交创制资源,以期快速获得新的优质种质。
2.2 4W1-056选育过程及其特性
以鄂中5号为母本、鉴真2号为父本进行杂交,F1代与鄂中5号进行回交得BC1F1,后代通过连续自交到BC1F7。在BC1F2—BC1F4系谱选育过程中,较早开展了混合株系的稻米品质分析检测,以筛选稻米的食味品质和米饭的膨松度。2014年冬季在海南BC1F7获得相对稳定的株系4W1-056,于2014—2016年在多地试验种植4W1-056,并以鄂中5号为对照品种,调查农艺性状,结果显示,4W1-056株型松散适度,剑叶挺直,生长量大,分蘖力强,茎秆粗壮,抗倒伏能力强,群体结构合理,后期转色好,农艺性状相对一致、稻米品质优良。
2.3 ZY56的选择过程
在常规方法无法继续筛选的情况下,收获全株系4W1-056的66个单株。2016年利用高通量SNP分型技术对收获的66个单株及鄂中5号、鉴真2号进行基因型分析,分析了908个SNP位点,涉及68个DNA片段,最终在310个位点找到差异,以每个位点的碱基差异数为单位,进行遗传距离和进化分析(图1)。
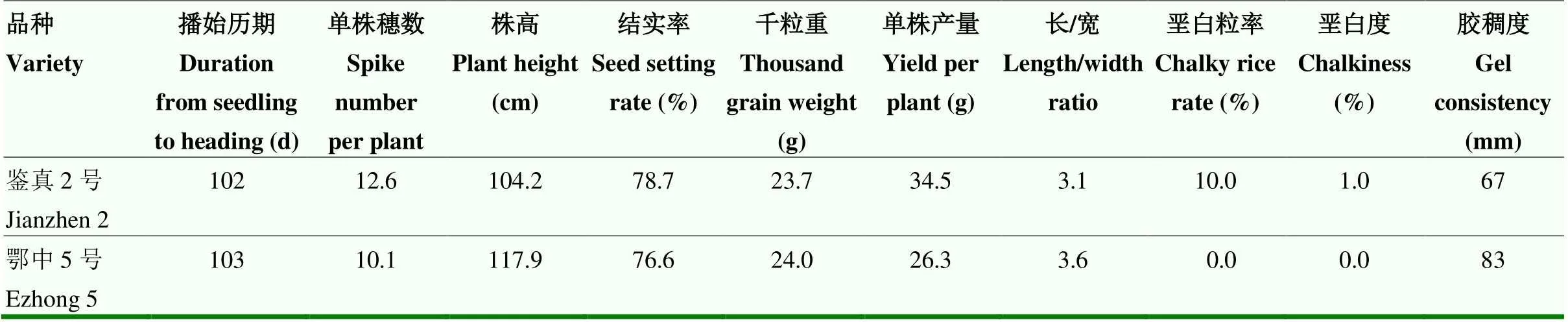
表1 鉴真2号与鄂中5号的主要性状表现
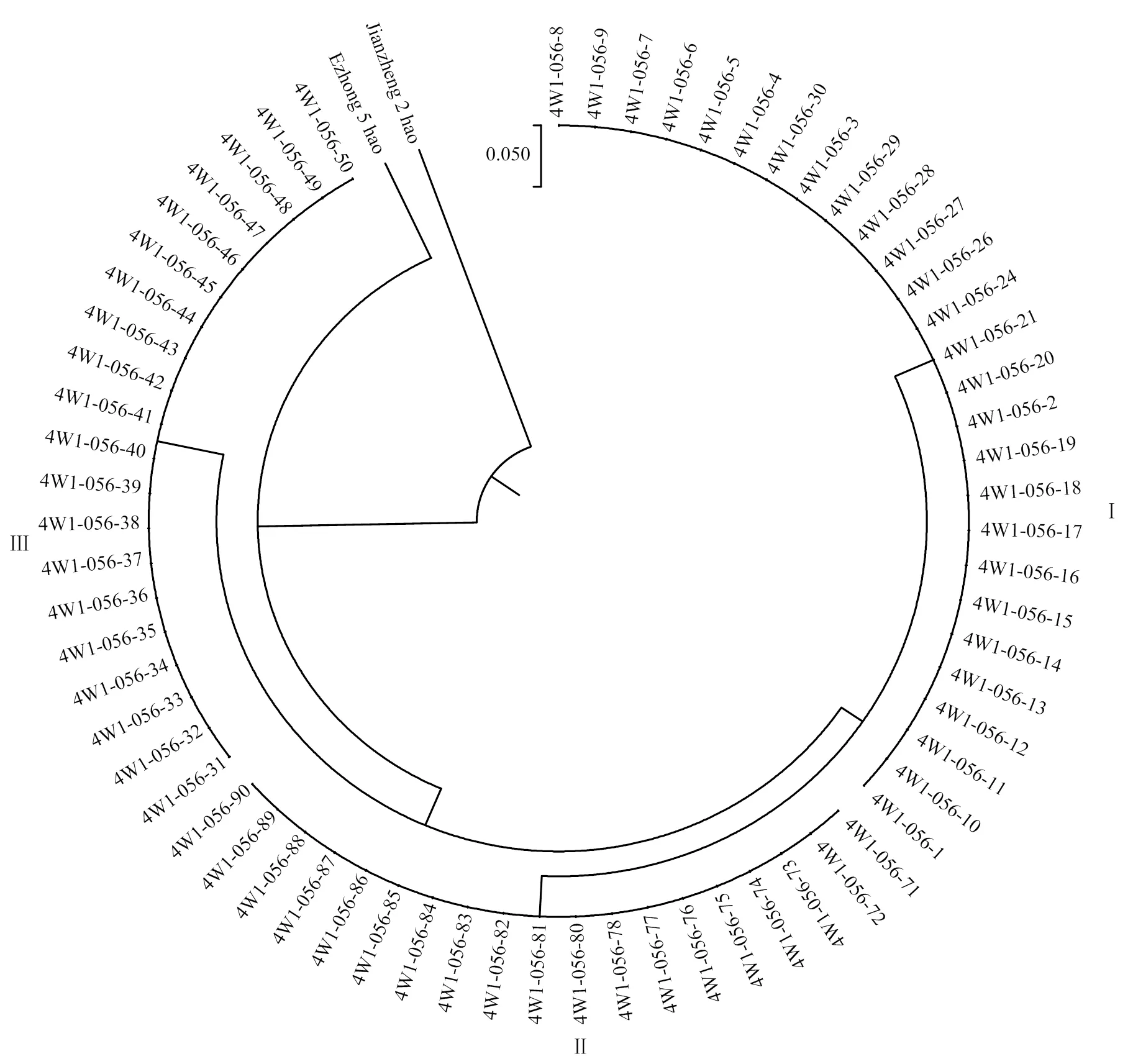
图1 4W1-056株系与鄂中5号、鉴真2号的遗传进化关系
鉴真2号与鄂中5号、4W1-056的遗传进化距离为0.6838,表明鉴真2号与鄂中5号及4W1-056的遗传背景差异较大;而4W1-056的66个单株与鄂中5号的遗传进化距离为0.2580,明显小于其与鉴真2号的遗传进化距离,表明4W1-056与鄂中5号之间的差异小于其与鉴真2号之间的差异,这与4W1-056的来源(鄂中5号/鉴真2号//鄂中5号)一致。而在4W1-056的66个单株之间,依据单株间的遗传分化,仍然可以分为3个类群(类群Ⅰ、类群Ⅱ和类群Ⅲ)。3个类群之间的碱基替代率为0.1398,类群Ⅰ和类群Ⅱ之间的碱基替代率为0.0662,而在各类群内的碱基替代率为0,表明同一类群内单株的遗传没有差异。由此可见,4W1-056各单株之间,在通过农艺性状测定等常规方法难以再细分的情况下,却可以利用分子生物学手段继续分出不同的类群,提供了继续选择的可能。各类群间的差异可以通过遗传分化(碱基替代率)来区分,而在各类群内,碱基替代率为0,可视为群体稳定。
将每个类群视为稳定群体,分别进行混合收种,并加代繁殖。群体自交至BC1F10,再次进行农艺性状、稻米品质的筛选(表2),发现类群Ⅱ具有生育期稍短、稻米外观品质更优等特征,因此,选择类群Ⅱ作为新的品系,命名为ZY56,于2020年获得植物新品种权证书(品种权号:CNA20180295.0)。类群Ⅰ和类群Ⅲ作为备用品系。
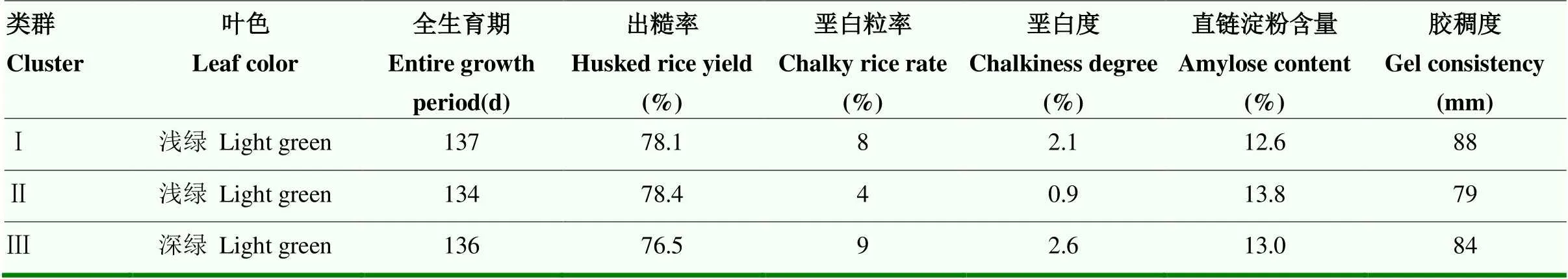
表2 4W1-056各类群之间部分性状的差异
2017年5月9日,在湖北武汉播种ZY56,全生育期为133.7 d,比对照鄂中5号早5 d,株高124.2 cm,有效穗数为280.5万/hm2,每穗总粒数为147.8粒,结实率为87.72%,千粒重25.1 g,产量稳定且比对照增产3.25%,稻瘟病综合指数为3.2,中抗稻瘟病。经过SNP分型选择和类群细分,ZY56抽穗集中,穗层整齐,农艺性状更稳定。
2.4 ZY56与亲本的SNP差异
采用水稻8K SNP芯片检测ZY56及2个亲本(鉴真2号和鄂中5号),结果(图2)表明,ZY56的染色体片段Chr.1:24536282—32696222、Chr.1:34520371—34964519、Chr.2:4803030—5632572、Chr.4:599979—6914238、Chr.8:26891056—28213513和Chr.11:17914093—18746042来源于亲本鉴真2号,其他均来源于亲本鄂中5号。统计表明,ZY56有14.13%的染色体片段来源于鉴真2号,85.87%的染色体片段来源于鄂中5号。8K SNP芯片检测结果与ZY56的系谱来源一致,验证了ZY56与亲本鉴真2号和鄂中5号的农艺性状既存在一定的差异,也有较多的相似性(如稻米品质、株高、生育期等)。
2.5 ZY56的温光反应特性
2017—2018年分别在湖北武汉、荆州、武穴、枣阳以及海南陵水等地播种ZY56,记录其生长过程中的积温、光照长度以考察ZY56温光反应特征(表3)。ZY56完成基本营养生长所需要的最低有效积温为760.5℃,转换为生殖生长的临界光照时长需短于14 h 13 min,完成幼穗分化需要有效积温高于711.5℃。有效积温能反映水稻生育期间对热量的要求,只要地理位置和其他外界条件变化不明显,通常同一品种对有效积温的要求是稳定的。由于ZY56由营养生长转换为生殖生长的临界光照时长短于14 h 13 min的要求会先于有效积温需求的条件,可以推测ZY56的生长适宜控制条件应当优先考虑有效积温是否满足。ZY56完成营养生长向生殖生长的转换,需有效积温大于760.5℃的条件。
2.6 ZY56的稻米品质特性
将2018年分期播种的ZY56和鄂中5号收获的种子送农业农村部食品质量监督检验测试中心分析稻米品质(表4)。随着播种期的推迟,ZY56表现出胶稠度下降,直链淀粉含量升高,垩白粒率和垩白度下降,外观品质变好;而鄂中5号的胶稠度和直链淀粉含量变化无规律,垩白粒率和垩白度逐渐升高,外观品质变差。5月18日播种的鄂中5号各项指标达到国标一级,垩白粒率和垩白度等指标优于ZY56;而6月12日播种的ZY56米质各项指标,尤其是整精米率、垩白粒率和垩白度、胶稠度等重要指标都优于鄂中5号。
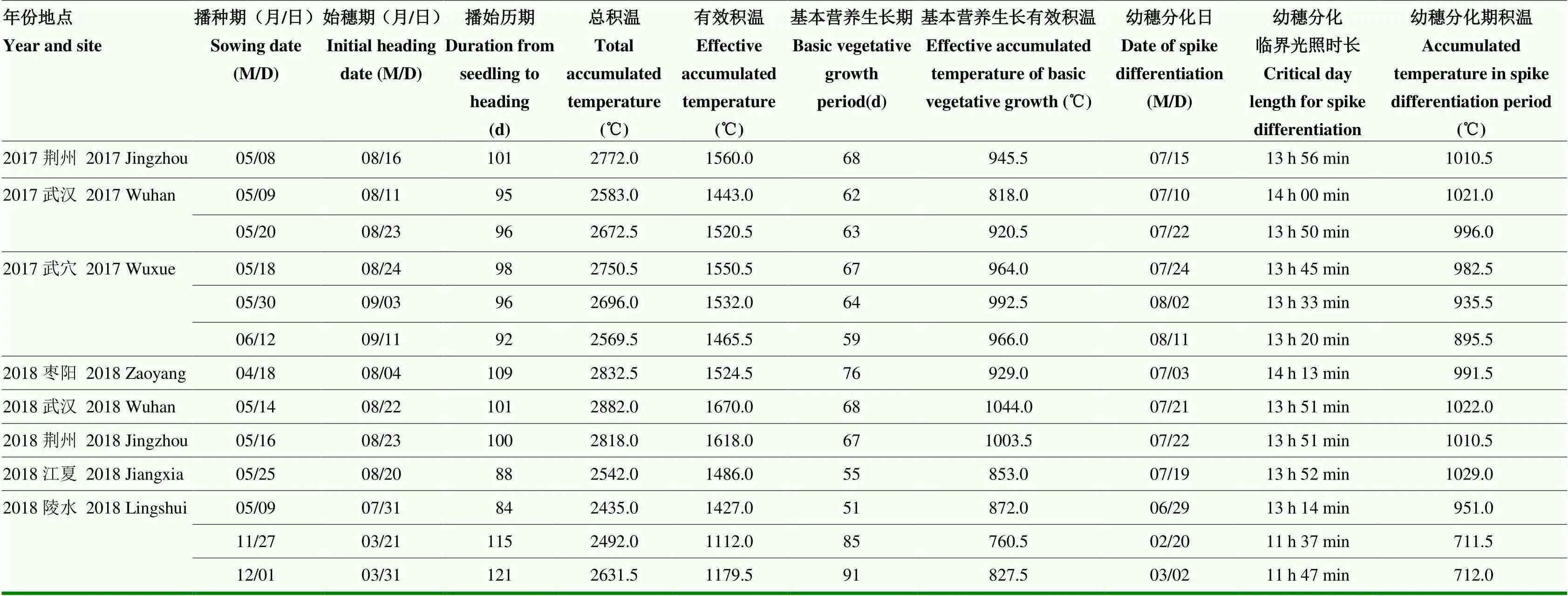
表3 ZY56在多个生长地点的温光反应
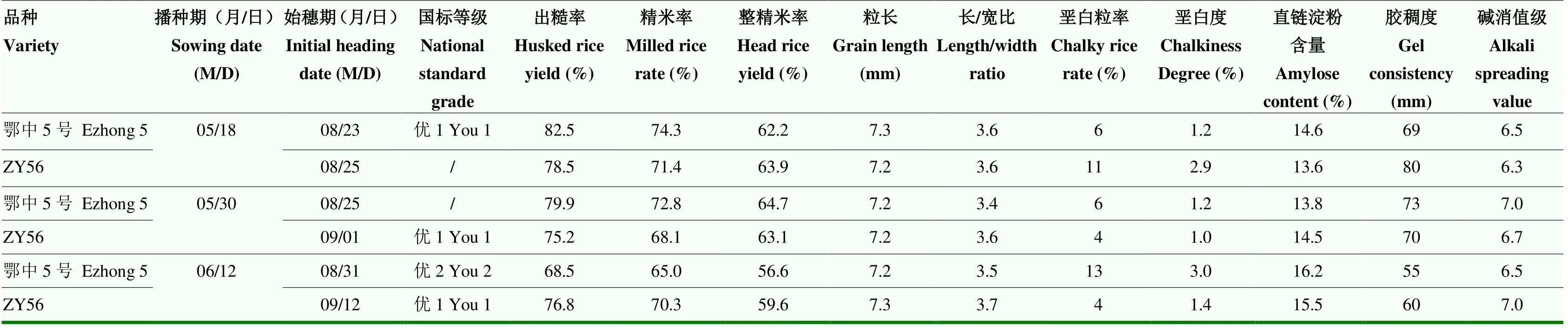
表4 不同播种期ZY56与鄂中5号的稻米品质分析结果(GB/T 17891-2017)
表4中播种始穗日期亦显示,分3期播种,播种期前后跨度25 d,光照长度变化明显,鄂中5号的播始历期分别为97、86和80 d,前后相差17 d;而ZY56的播始历期分别为99、94和92 d,前后相差7 d,说明ZY56对光照长度的反应明显弱于鄂中5号,但生长平稳,有利于稻米品质的形成。同时说明鄂中5号不宜迟播,短光照条件加速其生长发育,不利于稻米品质的发育。综上所述,ZY56宜迟于5月30日后播种,品质更佳。
经农业农村部稻米及制品质量监督检验测试中心分析不同播种期收获的ZY56和鄂中5号种子的黏度RVA谱的崩解值、峰值、终值、谷值、回冷值等5个特征值(表5)。结果显示,在3个播期间,二者的峰值(=0.013<0.05)差异显著;崩解值(=0<0.01)差异极显著;谷值(=0.149>0.05)、终值(=0.091>0.05)、回冷值(=0.815>0.05)差异不显著。鄂中5号的回冷值表现为第二播种期最小,无明显的变化规律,其他特征值表现出随着播种期的延迟而增加的趋势。ZY56的各个特征值均表现出随着播种期的延迟而增加的趋势。
米饭的食味品质与黏度RVA谱的峰值、谷值、崩解值、终值呈正相关,与RVA谱的回冷值呈负相关,崩解值和峰值的贡献更大,5个值对米饭食味品质的贡献大小排序为:崩解值>峰值>终值>谷值>回冷值。因此,可以认为随着播种期的延迟 (6月12日之前)ZY56和鄂中5号的食味品质均变好。
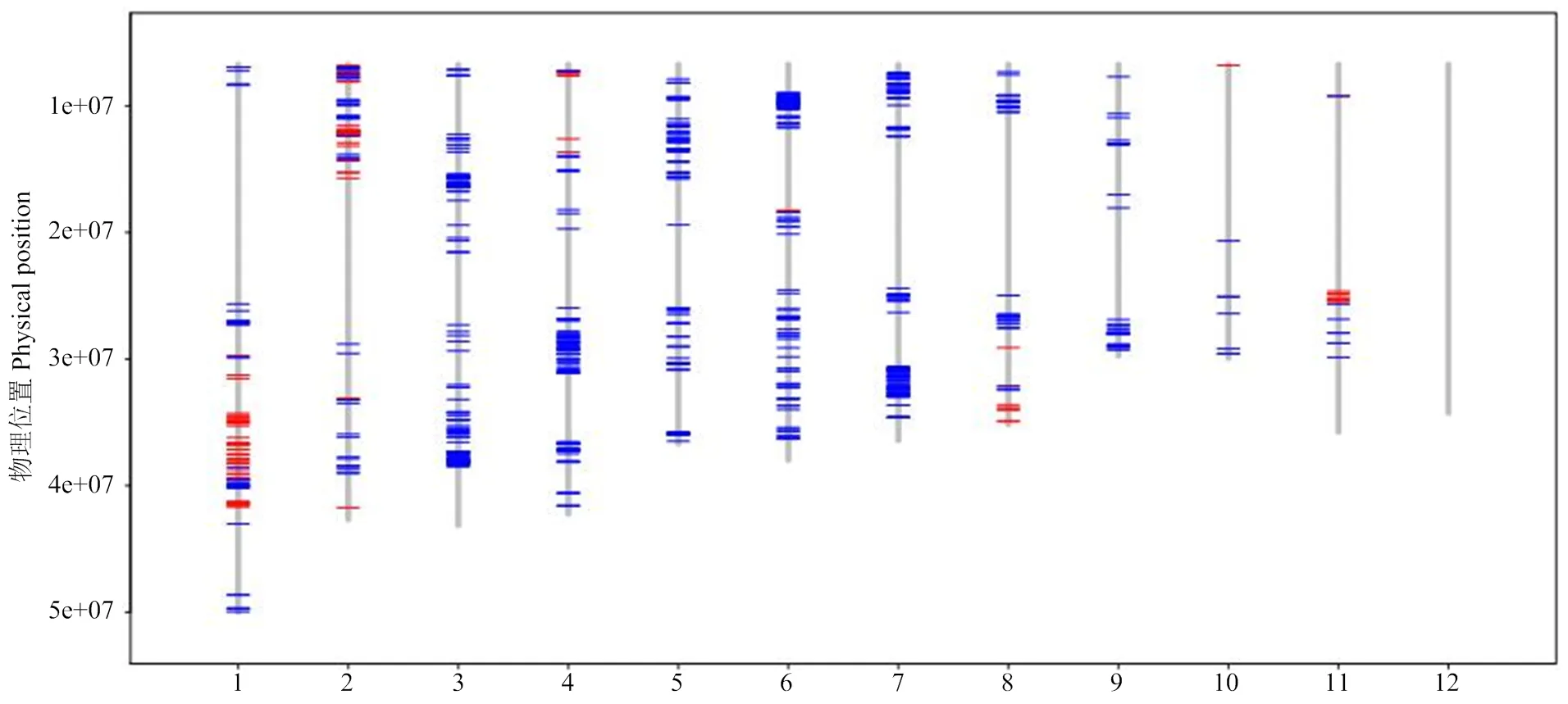
红色:来源于鉴真2号;蓝色:来源于鄂中5号 Red: the SNP sites come from Jianzhen 2; Blue: the SNP sites come from Ezhong 5
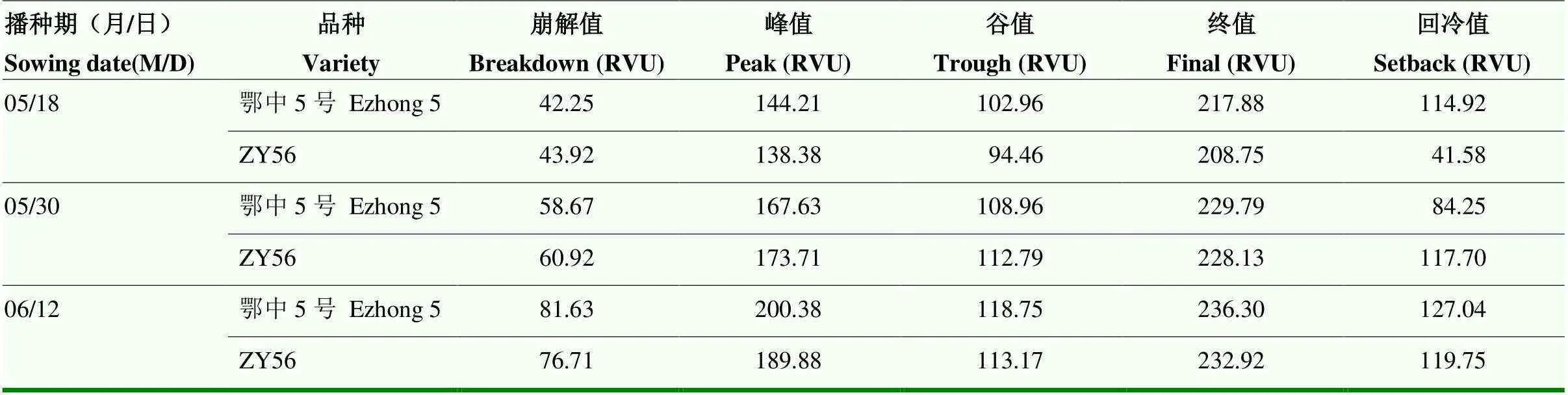
表5 不同播种期ZY56与鄂中5号的RVA谱特征值
为了探索食味品质与淀粉发育的关系,设置30、60和90 s等不同的加工精米时间,分析所得米粉的胶稠度和直链淀粉含量(表6)。经方差分析可知,同一播种期收获的稻谷不同加工精米时间,鄂中5号和ZY56的直链淀粉含量和胶稠度均表现为差异不显著,说明其在同一播期籽粒发育过程中,籽粒的内外层直链淀粉含量和胶稠度无明显的变化。不同的播期间,鄂中5号的直链淀粉含量和胶稠度差异显著;ZY56的直链淀粉含量差异显著,但胶稠度(=0.150>0.05)差异不显著,说明不同的抽穗灌浆期气候环境对其稻米品质的影响明显,而不同的品种表现出不同的变化规律,而ZY56的稻米品质具有更好的稳定性,这可能与其光温反应特性有关。
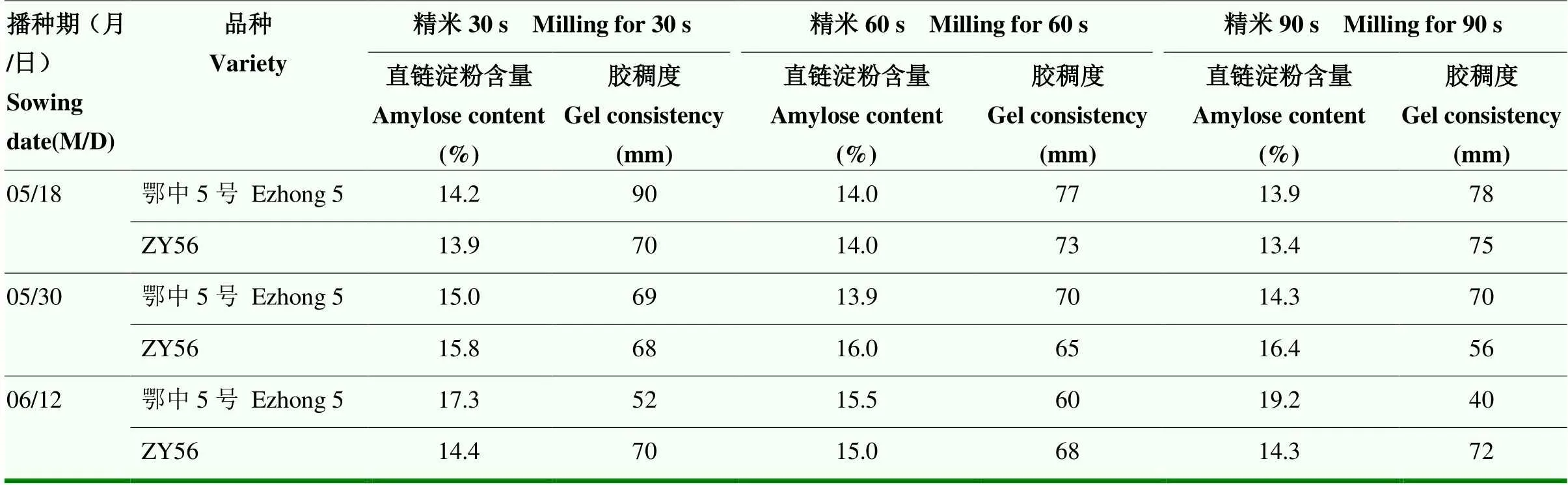
表6 不同加工精米时间ZY56与鄂中5号直链淀粉含量和胶稠度变化
3 讨论
3.1 优异水稻种质资源是优质稻育种的物质基础
鄂中5号、鉴真2号是湖北省审定的2个优质稻品种,2001—2018年累计推广面积80万hm2,是湖北的高档稻米品牌。但这两个品种都存在生育期偏长,耐寒性较差、不适宜轻简化栽培等缺点,已不适应当前优质稻产业高效发展的需求。本研究在保持鄂中5号、鉴真2号优质特性的基础上创制新的优质资源ZY56,具有生育期稳定、适宜直播和迟播等优点,在湖北各地自4月中旬至6月上旬均可播种,可轻简化栽培。与其亲本鄂中5号迟播品质不稳定的特性相比,ZY56适应性更广,具有更高的推广利用价值。
灌浆期气候环境因素对优质稻米资源的品质形成作用不容忽视。灌浆期是稻米品质形成的关键时期[21],温光因子影响品种的平均灌浆速率,因此,稻米品质的最终形成与环境因子关系密切。本研究为探索ZY56能获得优质高效的适宜栽培技术和生态环境,2017—2018年开展了多年多点种植试验,以推测其生育期间对光热资源的要求[27],同时评价不同的灌浆环境下稻米品质的变化[28-29],以确定其适宜的生产环境。因此,创制优质稻米资源应充分考虑其环境适应性,并制定配套的生产标准,以指导优质稻米生产。
3.2 传统育种方法与生物技术的结合加速了种质创新
种质资源的精准鉴定是其创新利用的基础。在ZY56的创制过程中,分析了选用的2个亲本的农艺性状,鄂中5号株高较高,鉴真2号株高较矮但茎秆较柔软,但2个品种均具有稻米品质优良的特点。根据邱东峰等[30]利用亲本的遗传相似系数结合农艺性状的差异选择亲本的方法,辜大川等[31]在27份水稻资源进行SSR分析结果显示,鉴真2号与鄂中5号的遗传相似系数为0.6822,小于平均遗传相似系数0.7536,显示它们的亲缘关系较远;同时,前述进行的高通量SNP分型分析亦表明,鉴真2号与鄂中5号遗传进化距离较远,遗传背景差异较大,这与利用SSR分子标记评价的结果吻合。在亲本差异较大的情况下宜采用杂交后回交的方法可快速稳定目标性状。因此,在ZY56的创制过程中采用杂交后再回交一次,快速获得相对稳定的后代群体,并从BC1F2代开始连续评价其稻米外观品质、蒸煮和食味品质,确保了品质性状选择的有效性。亲本选择中将表型鉴定和遗传背景评价相结合,提高了育种的成功率。
在新种质选育过程中,利用常规方法往往无法准确鉴定评价其稳定性、一致性,如果不能进行有效分型选择,所选品系很难快速稳定,甚至会丢失部分优良品系。因此应在选育过程中引入分子评价的方法进行遗传背景的筛选,以提高选择的效率和多样性。长期以来,对稻种资源的遗传多样性分群分析的研究[32-36]较多,但如何从分析的水稻资源中利用遗传距离的相关数据信息去创新材料或培育新品种,这些研究中并未涉及。本研究选择亲本以及新种质选育过程中利用的新方法,降低了选择的工作量,提高了选择效率。
稻米的品质性状由多基因控制,且各性状间彼此交叉,其遗传调控非常复杂,尽管很多基因的功能已比较清楚,但却不能有效地指导育种实践[20]。对于此类多基因控制的复杂性状,以及更多未知基因控制的其他性状,目前,在育种实践中并未见有较好的解决方法,本研究提出的通过高能量SNP分型对遗传背景进行综合评价的方法,比较有效地解决了这类问题。
4 结论
种质创制高代选择中,常规方法获得的相对稳定株系,利用高通量SNP分型技术进行遗传背景或未知基因的一致性筛选,可最终确定符合目标要求的株系。该方法克服传统系谱法在高世代选择中针对农艺性状难以继续选择的困难,具有避免同类单株重复选择和不同类单株漏选的问题,降低了高世代选择工作量,提高了选择效率,实用性强,具有推广的价值。
[1] 黎舒佳, 高谨, 李家洋, 王永红. 独脚金内酯调控水稻分蘖的研究进展. 植物学报, 2015, 50(5): 539-548.
LI S J, GAO J, LI J Y, WANG Y H. Advances in studies on the regulation of rice tiller by strigolactone. The Plant Journal, 2015, 50(5): 539-548. (in Chinese)
[2] 黎裕, 李英慧, 杨庆文, 张锦鹏, 张金梅, 邱丽娟, 王天宇. 基于基因组学的作物种质资源研究: 现状与展望. 中国农业科学, 2015, 48(17): 3333-3353.
LI Y, LI Y H, YANG Q W, ZHANG J P, ZHANG J M, QIU L J, WANG T Y. Genomics-based crop germplasm research: Advances and perspectives. Scientia Agricultura Sinica, 2015, 48(17): 3333-3353. (in Chinese)
[3] 胡兴明, 钱前. 现阶段中国水稻种质创新的研究策略和应用思考. 植物遗传资源学报, 2004, 5(2): 193-196.
HU X M, QIAN Q. Current research strategies and ways of utilization of new rice germplasm in China. Journal of Plant Genetic Resources, 2004, 5(2): 193-196. (in Chinese)
[4] 余泓, 王冰, 陈明江, 刘贵富, 李家洋. 水稻分子设计育种发展与展望. 生命科学, 2018, 30(10): 1032-1037.
YU H, WANG B, CHEN M J, LIU G F, LI J Y. Research advance and perspective of rice breeding by molecular design. Chinese Bulletin of Life Sciences, 2018, 30(10): 1032-1037. (in Chinese)
[5] Zuo J, Li J. Molecular dissection of complex agronomic traits of rice: a team effort by Chinese scientists in recent years. National Science Review,2014, 1: 253-276.
[6] 符德保, 李燕, 肖景华, 张启发, 吴昌银. 中国水稻基因组学研究历史及现状. 生命科学, 2016, 28: 1113-1121.
FU D B, LI Y, XIAO J H, ZHANG Q F, WU C Y. The history and current status of rice genomics research in China. Chinese Bulletin of Life Sciences, 2016, 28: 1113-1121. (in Chinese)
[7] Li Y, Xiao J H, Chen L L, HUANG X H, CHENG Z K, HAN B, ZHANG Q F, WU C Y. Rice functional genomics research: past decade and future. Molecular Plant, 2018, 11: 359-380.
[8] Wang A H, Hou Q Q, Si L Z, HUANG X H, LUO J H, LU D F, ZHU J J, SHANGGUAN Y Y, MIAO J S, XIE Y F, WANG Y C, ZHAO Q, FENG Q, ZHOU C C, LI Y, FAN D L, LU Y Q, TIAN Q L, WANG Z X, HAN B. The PLATZ transcription factor GL6 affects grain length and number in rice. Plant Physiology, 2019, 180: 2077-2090.
[9] Xue W Y, Xing Y Z, Weng X Y, ZHAO Y, TANG W J, WANG L, ZHOU H J, YU S B, XU C G, LI X H, ZHANG Q F. Natural variation inis an important regulator of heading date and yield potential in rice. Nature Genetics, 2008, 40: 761-767.
[10] Wang J, Zhou L, Shi H, MAWSHENG C, YU H, YI H, HE M, YIN J J, ZHU X B, LI Y, LI W T, LIU J L, WANG J C, CHEN X Q, QING H, WANG Y P, LIU G F, WANG W M, LI P, WU X J, ZHU L H, ZHOU J M, PAMELA C R, LI S Q, LI J Y, CHEN X W. A single transcription factor promotes both yield and immunity in rice. Science, 2018, 361: 1026-1028.
[11] Tian Z x, Qian Q, Liu Q Q, YAN M X, LIU X F, YAN C J, LIU G F, GAO Z Y, TANG S Z, ZENG D L, WANG Y H, YU J M, GU M H, LI J Y. Allelic diversities in rice starch biosynthesis lead to a diverse array of rice eating and cooking qualities. Proceedings of the National Academy of Sciences of the United States of America, 2009, 106: 21760-21765.
[12] Zhao D S, Li Q F, Zhang C Q, ZHANG C Q, ZHANG C, YANG Q Q, PAN L X, REN X Y, LU J, GU M H, LIU Q Q.acts as a transcriptional activator to regulate rice grain shape and appearance quality. Nature Communications, 2018, 9: 1240.
[13] Huang X Y, Chao D Y, Gao J P, ZHU M Z, SHI M, LIN H X. A previously unknown zinc finger protein, DST, regulates drought and salt tolerance in rice via stomatal aperture control.Genes & Development, 2009, 23: 1805-1817.
[14] Li X M, Chao D Y, Wu Y, HUANG X H, CHEN K, CUI L G, SU L, YE W W, CHEN H, CHEN H C, DONG N Q, GUO T, SHI M, FENG Q, ZHANG P, HAN B, SHAN J X, GAO J P, LIN H X. Natural alleles of a proteasome α2 subunit gene contribute to thermotolerance and adaptation of African rice. Nature Genetics, 2015, 47: 827-833.
[15] Liu Y Y, Cao Y L, Zhang Q L, LI X H, WANG S P. A cytosolic triosephosphate isomerase is a key component in XA3/XA26- mediated resistance. Plant Physiology, 2018, 178: 923-935.
[16] Hu L, Wu Y, Wu D, RAO W W, GUO J P, MA Y H, WANG Z Z, SHANGGUAN X X, WANG H Y, XU C X, HUANG J, SHI S J, CHEN R Z, DU B, ZHU L L, HE G C. The coiled-coil and nucleotide binding domains of BROWN PLANTHOPPER RESISTANCE14 function in signaling and resistance against planthopper in rice. The Plant Cell, 2017, 29: 3157-3185.
[17] Hu B, Wang W, Ou S j, TANG J Y, LI H, CHE R H, ZHANG Z H, CHAI X Y, WANG H R, WANG Y Q, LIANG C Z, LIU L C, PIAO Z Z, DENG Q Y, DENG K, XU C, LIANG Y, ZHANG L H, LI L G, CHU C C. Variation incontributes to nitrate-use divergence between rice subspecies. Nature Genetics, 2015, 47: 834-838.
[18] Wang Q, Nian J Q, Xie X Z, YU H, ZHANG J, BAI J T, DONG G J, HU J, BAI B, CHEN L C, XIE Q J, FENG J, YANG X L, PENG J L, CHEN F, QIAN Q, LI J Y, ZUO J R. Genetic variations inmediate grain yield by modulating nitrogen utilization in rice. Nature Communications, 2018, 9: 735.
[19] 张先文, 贺治洲, 江南, 邓华凤, 李继明. 高通量基因型分型技术及其在水稻中的应用. 生物技术通报, 2017, 33(12): 67-73.
ZHAng X w, HE Z z, JIANG N, DENG H f, LI J m. High-throughput genotyping techniques and their applications in rice. Biotechnology Bulletin, 2017, 33(12): 67-73. (in Chinese)
[20] 张昌泉, 赵冬生, 李钱峰, 顾铭洪, 刘巧泉. 稻米品质性状基因的克隆与功能研究进展. 中国农业科学, 2016, 49(22): 4267-4283.
Zhang C Q, Zhao D S, Li Q F, Gu M H, Liu Q Q. Progresses in research on cloning and functional analysis of key genes involving in rice grain quality. Scientia Agricultura Sinica, 2016, 49(22): 4267-4283. (in Chinese)
[21] 王惠贞, 吴瑞芬, 李丹. 稻米品质形成和调控机理概述. 中国稻米, 2016, 22(1): 10-13.
Wang H Z, Wu R F, Li D. Review on rice quality formation and its regulation mechanism. China Rice, 2016, 22(1): 10-13. (in Chinese)
[22] 李宏凯, 徐得泽, 陈亿毅. 优质中籼稻新品种鄂中5号的选育. 湖北农业科学, 2005(1): 31-32.
Li H K, Xu D Z, Chen Y Y. Breeding of a new middle indica rice line Ezhong 5 with high quality. Hubei Agricultural Sciences, 2005(1): 31-32. (in Chinese)
[23] 郑明. 高档优质中籼稻鄂中5号在孝感的种植表现. 中国稻米, 2015, 21(6): 105-106.
Zheng M. Planting performance of fine quality indica rice Ezhong 5 in Xiaogan City. China Rice, 2015, 21(6): 105-106. (in Chinese)
[24] 张再君, 杨金松. 中国水稻品种志(湖北卷). 北京: 中国农业出版社, 2018: 76-79.
Zhang Z j, Yang J s. China Rice Varieties (Hubei Juan). Beijing: China Agriculture Press, 2018: 76-79. (in Chinese)
[25] Nei M, Kumar S. Molecular Evolution and Phylogenetics. New York: Oxford University Press, 2000.
[26] Kumar S, Stecher G, Tamura K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Molecular Biology and Evolution, 2016, 33: 1870-1874.
[27] 薛大伟, 方茂庭, 钱前. 有效积温在水稻生产中的应用. 中国稻米, 2004(4): 47-48.
Xue D W, Fang M T, Qian Q. Application of effective accumulated temperature in rice production. China Rice, 2004(4): 47-48. (in Chinese)
[28] 舒庆尧, 吴殿星, 夏英武, 高明尉, Anna McClung. 稻米淀粉RVA谱特征与食用品质的关系. 中国农业科学, 1998, 31(3): 25-29.
Shu Q Y, Wu D X, Xia Y W, Gao M W, Anna M C. Relationship between RVA profile character and eating quality inL.. Scientia Agricultura Sinica, 1998, 31(3): 25-29. (in Chinese)
[29] 隋炯明, 李欣, 严松, 严长杰, 张蓉, 汤述翥, 陆驹飞, 陈宗祥, 顾铭洪. 稻米淀粉RVA谱特征与品质性状相关性研究. 中国农业科学, 2005, 38(4): 657-663.
Sui J M, Li X, Yan S, Yan C J, Zhang R, Tang S Z, Lu J F, Chen Z X, Gu M H. Studies on the rice RVA profile characteristics and its correlation with the quality. Scientia Agricultura Sinica, 2005, 38(4): 657-663. (in Chinese)
[30] 邱东峰, 张再君, 辜大川, 殷明珠, 焦春海, 杨金松. 水稻R7606、229A系列优异资源的创制. 植物遗传资源学报, 2013, 14(5): 918-924.
Qiu D f, Zhang Z j, Gu D c, Yin M z, Jiao C h, Yang J s. Breeding of elite rice lines R7606 and 229A. Journal of Plant Genetic Resources, 2013, 14(5): 918-924. (in Chinese)
[31] 辜大川, 邱东峰, 殷明珠, 焦春海, 杨金松, 张再君, 刘春萍. 水稻优异资源分子身份证的构建. 湖北农业科学, 2012, 51(24): 5581-5586.
Gu D c, Qiu D f, Yin M z, Jiao C h, Yang J s, Zhang Z j, Liu C p. Establishment of molecular identity card of elite rice resources. Hubei Agricultural Sciences, 2012, 51(24): 5581-5586. (in Chinese)
[32] 李丹婷, 农保选, 夏秀忠, 刘开强, 杨庆文, 张宗文, 高国庆. 东南亚稻种资源收集与鉴定评价. 植物遗传资源学报, 2012, 13(4): 622-625.
LI D t, Nong B x, XIA X z, LIU K q, YANG Q W, ZHANG Z W, GAO G Q. Collection and evaluation of rice germplasm from Southeast Asia. Journal of Plant Genetic Resources, 2012, 13(4): 622-625. (in Chinese)
[33] 束爱萍, 刘增兵, 余丽琴, 黎毛毛, 陈大洲. 水稻SSR标记的遗传多样性研究进展. 植物遗传资源学报, 2013, 14(5): 918-924.
SHU A p, LIU Z b, YU L q, LI M m, CHEN D z. Research progress on genetic diversity in rice based on SSR marker analysis. Journal of Plant Genetic Resources, 2013, 14(5): 918-924. (in Chinese)
[34] 刘传光, 张桂权. 用SSR标记分析1949-2005年华南地区常规籼稻主栽品种遗传多样性及变化趋势. 作物学报, 2010, 36(11): 1843-1852.
LIU C g, ZHANG G q. SSR analysis of genetic diversity and the temporal trends of major commercial inbredrice cultivars in south China in 1949-2005. Acta Agronomica Sinica, 2010, 36(11): 1843-1852. (in Chinese)
[35] Zeng Y W, Zhang H L, Li Z C, SHEN S Q, SUN J L, WANG M X, LIAO D Q, LIU X, WANG X K, XIAO F H, WEN G S. Evolution of genetic diversity of rice landraces (L.) in Yunnan, China. Breeding Science, 2007, 57: 91-99.
[36] 陈峰, 朱其松, 徐建第, 孙公臣,柳发财,朱文银,张洪瑞,高洁,袁守江. 山东地方水稻品种的农艺性状与品质性状的多样性分析. 植物遗传资源学报, 2012, 13(3): 393-397.
CHEN F, ZHU Q s, XU J d, SUN G c, LIU F c, ZHU W y, ZHANG H r, GAO J, YUAN S j. Diversity analysis of agronomic and quality characters of rice landraces in Shandong. Journal of Plant Genetic Resources, 2012, 13(3): 393-397. (in Chinese)
Breeding and Evaluation of Elite Rice Line ZY56
QIU DongFeng1, GE PingJuan1, Liu Gang1, Yang JinSong1, Chen JianGuo2, ZHANG ZaiJun1
1Food Crop Institute, Hubei Academy of Agricultural Sciences/Hubei Key Laboratory of Food Crop Germplasm and Genetic Improvement, Wuhan 430064;2School of Life Sciences, Hubei University, Wuhan 430062
【】In order to avoid the influences of unknown genes, genetic background and other uncontrollable factors in breeding practice, experiments were conducted to create new high-quality rice germplasm using an improved method, which integrated high-throughput SNP genotyping with traditional breeding to improve breeding efficiency.【】Two varieties of rice (Jianzhen 2 and Ezhong 5) were used as parents to make a cross (Ezhong 5/Jianzhen 2), and Ezhong 5 was used as recurrent parent to obtain backcross generations, whose progenies were selected by pedigree method. Rice qualities were evaluated in generations from BC1F2to BC1F4. An elite line, 4W1-056, was screened in the generation of BC1F7. DNA segments from Jianzhen 2, Ezhong 5 and 66 plants of 4W1-056 were PCR-amplified and sequenced using capture sequencing technologies, and 908 SNP sites were analyzed. Phylogenetic analyses were performed based on genotypes of SNP sites using p-distance method in software MEGA7. Novel lines were selected and evaluated based on agronomic traits and rice qualities in combination with the results from phylogenetic analyses.【】An elite line 4W1-056 was obtained by means of crossing, backcrossing and selfing, which has desirable plant type, strong tillering ability, good lodging resistance for stocky stems, excellent appearance quality and preliminarily stable in agronomic traits. Sixty-six plants selected from 4W1-056 were clustered into three groups based on the results from phylogenetic analyses.The base substitution rate among cluster Ⅰ, Ⅱ and cluster Ⅲ was 0.1398, and that between cluster I and Ⅱ was 0.0662. The base substitution rate within any of these clusters was 0, which suggested that there were no genetic differences among individual plants within the same cluster. Cluster II was designated ZY56 as a new line based on its agronomic traits and rice qualities, and its appearance quality is better than that of 4W1-056, with a chalkiness score of 0.9, which is closer to the appearance quality performance of Ezhong 5. ZY56 and its original parents were detected using 8K RICE SNP chips, the results showed that 14.13% of ZY56’s chromosome segments were from Jianzhen 2 and 85.87% were from Ezhong 5, Chip detection further showed that most of the genes in ZY56 originated from Ezhong 5, and verified the selection results after crossing, backcrossing and selfing. The minimum effective accumulated temperature needed for basic vegetative growth in ZY56 was 760.5℃, the critical light length for reproductive growth was 14 h 13 min, and the effective accumulated temperature needed for panicle differentiation was above 711.5℃. The results of quality analyses from experiments at different sowing dates showed that ZY56 responded to light length significantly weaker than Ezhong 5, its photosensitivity was weak and growth was stable, which was conducive to the development of rice qualities, and the rice qualities of ZY56 were more stable than Ezhong 5. 【】We proposed a new method for the selection in higher generations of a germplasm development program, in which high-throughput SNP genotyping technology was used to screen plants for genetic consistency, and eventually to find out the strains that conform to breeding targets. This method overcame the difficulty in the selection for agronomic traits by means of traditional pedigree in advanced generations, and circumvented the problems that similar types of strains were repeatedly selected and different types were missed. The method saved efforts of selection in higher generations, while increased the efficiency of selection, so it had value of popularization.
rice; germplasm improvement; resources evaluation

10.3864/j.issn.0578-1752.2021.06.001
2020-08-13;
2020-11-05
国家重点研发计划(2016YFD0100101-05)、国家作物种质资源库(湖北分库)(NICGR2020-31)、物种资源保护费(1120162130135252037)
邱东峰,Tel:18672779158;E-mail:qdflcp@163.com。通信作者张再君,Tel:13871426107;E-mail:zjzhang0459@aliyun.com。通信作者陈建国,E-mail:jgchen@hubu.edu.cn
(责任编辑 李莉)
猜你喜欢
美食(2022年5期)2022-05-07
早期教育(家庭教育)(2021年11期)2021-12-17
少儿科学周刊·儿童版(2021年21期)2021-12-11
绿色科技(2019年14期)2019-11-19
江苏农业科学(2019年5期)2019-09-02
民间故事选刊·下(2018年5期)2018-05-18
文苑·感悟(2018年4期)2018-04-09
江苏农业科学(2017年17期)2017-11-15
小说评论(2014年3期)2014-11-14
小小说月刊·下半月(2013年12期)2013-05-14
