
枫香刺小蠹雌雄成虫的形态差异、繁殖特性及其坑道系统形成过程
2021-04-01 10:59鞠瑞亭
复旦学报(自然科学版) 2021年1期
赵 洁,高 磊,鞠瑞亭
(1.复旦大学 生命科学学院 生物多样性和生态工程教育部重点实验室, 上海 200438; 2.上海市园林科学规划研究院 上海城市困难立地绿化工程技术研究中心, 上海 200232)
小蠹是鞘翅目(Coleoptera)、象甲科(Curculionidae)、小蠹亚科(Scolytinae)昆虫的统称,是世界公认的对树木最具危害性的昆虫类群之一,具有繁殖能力强、发生范围广、危害隐蔽、易暴发成灾等特点[1-2].虽然大多数小蠹以攻击垂死或衰弱的树木为主,但仍然有许多种类会攻击健康树木,发生严重时会导致树木大规模死亡[3-5].随着国际贸易的快速发展,木材贸易和外来植物的引种愈加频繁,这一过程极易造成具潜在危险性的外来小蠹传入新区,形成生物入侵问题,给土著生态系统和农林生产造成巨大危害[6],例如,20世纪末入侵到我国的红脂大小蠹(Dendroctonusvalens)在山西、河北等多个省份暴发,造成了至少700万株松树死亡,直接经济损失超过6.84亿元[7-8].
枫香刺小蠹(Acanthotomicussuncei)是近年来在我国东部城市化地区发现的一种新兴害虫[7-8].该虫隶属齿小蠹族(Ipini)刺小蠹属(Acanthotomicus),是枫香属(Liquidambar)树木上的主要蛀干性害虫,尤其对新引种植物北美枫香(Liquidambarstyraciflua)的为害最为严重[11].北美枫香是2000年前后作为观赏树、行道树和防护林树种引入我国的外来乔木,目前已在华东、华中等地区广泛栽植[12-13].然而,近年来,北美枫香遭受枫香刺小蠹的严重为害,被危害的植株通常在半年之内枯萎,已导致上海多个苗圃地中90%以上的北美枫香死亡[11].此外,根据经济损失模型预测,若该虫传入到北美地区,将造成大约1.52亿美元的经济损失[14].
目前,国内外学者虽然已对枫香刺小蠹的生活史动态、发生为害情况和经济损失评估做了一部分研究[9-11],但有关成虫的性别快速识别、繁殖习性及坑道形成过程等信息仍很缺乏.为此,本文对枫香刺小蠹成虫的雌雄形态差异进行了补充描述,同时通过室内饲养,进一步明确了其繁殖习性,并记录了饲养过程中成虫坑道系统的形态特征及形成过程,其目的是为更加全面地掌握该虫的生物学特性提供参考资料.
1 材料与方法
1.1 昆虫采集及饲养
在上海浦东南汇新城镇某北美枫香苗圃地,选取未受枫香刺小蠹危害健康的北美枫香树干以及已受该虫危害的树干(胸径均为8~10cm),锯成若干个15cm长的木段,在切口两端涂上石蜡后用PVC膜封住,以减少水分蒸发.在枫香刺小蠹成虫羽化高峰期,将健康木段与带虫木段放置在一起,每日检查是否有小蠹侵入健康木段,并对新出现的侵入孔进行标记;然后将有新侵入孔的木段单独放入透明塑料盒(21cm×12cm×11cm)中饲养.在塑料盒盖和侧面,预先打若干孔洞并用滤网(200目)封孔,保证盒内外温湿度一致.所有塑料盒被放置于恒温培养箱(三洋MIR 350H)中培养,培养环境为: 温度(30±1)℃、相对湿度(RH)60%~70%、光照周期14L∶10D.
1.2 雌雄成虫形态差异
在枫香刺小蠹新种描述及各虫态关键形态特征文献报道的基础上[10,12],对其雌雄成虫的形态差异进行补充描述;并随机选取成虫标本93头(雄∶雌=46∶47),分别测量雌雄成虫的体长和前胸背板的最宽宽度,比较其形态大小的差异性.
1.3 繁殖特性
待新一代成虫完全羽化后,统计树皮上每窝小蠹的羽化孔数量,以此作为最终羽化成虫的数量.利用木工凿、镊子等工具移除受害木段(n=15)的树皮,结合树皮内侧和木质部保留的坑道形状,观察记录每窝小蠹的母坑道数、产卵量、子坑道数和蛹室数等参数,根据母坑道数量确定每窝母代雌成虫的数量,产卵量以每条母坑道上的卵痕数来指示.
各参数计算方法为: 孵化率=(子坑道数/卵痕数)×100%;化蛹率=(蛹室数/子坑道数)×100%;羽化率=(羽化孔数/蛹室数)×100%.
1.4 坑道发展过程观察
在枫香刺小蠹侵入树木后,每3天随机解剖10个侵染坑道,观察并记录坑道发展情况,取其中最为相似的坑道形状,作为坑道发展的典型阶段模式,绘制不同母坑道类型和子坑道后期的形态示意图.随机选取母坑道和子坑道各20条,分别测量其长度,并计算其平均值.
1.5 数据分析
对所有数据进行正态和方差齐性检验后,采用t测验比较雌雄成虫体长、前胸背板的最宽宽度的差异.采用单因素方差分析(One-way ANOVA)检验坑道内营不同性比的成虫的单雌产卵量、孵化率和化蛹率等繁殖指标的差异,若处理间存在显著差异,结合Duncan’s多重比较法进行事后检验.所有数据处理均在SPSS 23.00软件上进行.
2 结 果
2.1 雌雄成虫形态差异
枫香刺小蠹雄虫鞘翅翅盘边缘着生4对齿,分别位于第2、4、6、8刻点沟与翅盘相连处隆起的部位,4对齿较明显,在第7刻点沟在近翅盘边缘着生1对小瘤突,其外形与齿相似,但小于前4对齿(图1(a)).雌成虫的4对齿、1对瘤突着生与排列方式和雄虫一致,但4对齿的长度均小于雄虫,且第2对齿的着生部位更靠近翅盘中部(图1(b)).

图1 枫香刺小蠹雄雌成虫鞘翅翅盘形态特征
93头枫香刺小蠹成虫(雄∶雌=46∶47)体长和前胸背板的最宽宽度的测量结果显示,雄成虫体长范围为1.93~2.69mm(平均值=(2.37±0.03)mm),前胸背板的最宽宽度范围为0.66~0.86mm(平均值=(0.78±0.01)mm);雌成虫体长范围为1.96~2.67mm(平均值=(2.36±0.03)mm),前胸背板的最宽宽度范围为0.68~0.83mm(平均值=(0.77±0.01)mm).统计结果发现,雌雄成虫体长和前胸背板的最宽宽度无显著差异(t=0.228,df=91,P>0.05).
2.2 繁殖特性
枫香刺小蠹主坑道内成虫性别组合主要有1雄1雌、1雄2雌、1雄3雌、1雄4雌等4种形式.坑道内单雌产卵量为13~71粒不等(平均值=(35±3.75)粒).不同性比情况下单雌产卵量、孵化率、化蛹率和羽化率如表1所示,其中单雌产卵量(F=14.850,df=3,P<0.01)、卵孵化率在具不同性比的坑道处理间有显著差异(F=16.824,df=3,P<0.01),每窝雌雄比1∶1和2∶1的坑道内单雌繁殖力,显著高于雌雄比较高(3∶1和4∶1)的坑道内成虫的繁殖力.此外,具不同性比的坑道处理间化蛹率(F=0.245,df=3,P>0.05)和孵化率(F=0.157,df=3,P>0.05)无显著差异.

表1 不同性比坑道内枫香刺小蠹单雌产卵量、孵化率、化蛹率和羽化率
2.3 坑道系统的发展过程
(1)侵入孔和交配室
雄成虫首先在枫香树干上钻蛀一个直径约为1mm的圆形侵入孔(图2(a)),并在树皮下形成交配室(图2(b)),再吸引1至数头雌虫进入交配(图2(c)).

图2 枫香刺小蠹侵入初期活动阶段
(2)母坑道形成及产卵
完成交配后的雌虫沿交配室周围不同方向挖掘母坑道(随雌虫数不同有所差异),母坑道主要横向伸展,形状多样(图3),有时会形成近似环形带.母坑道的宽范围为0.9~1.0mm、长范围为2.5~7.0cm(平均值=(4.2±0.3)cm).卵产在母坑道两侧的卵室中,每个卵室内产卵1粒,同侧相邻两粒卵的间隔约0.9mm.
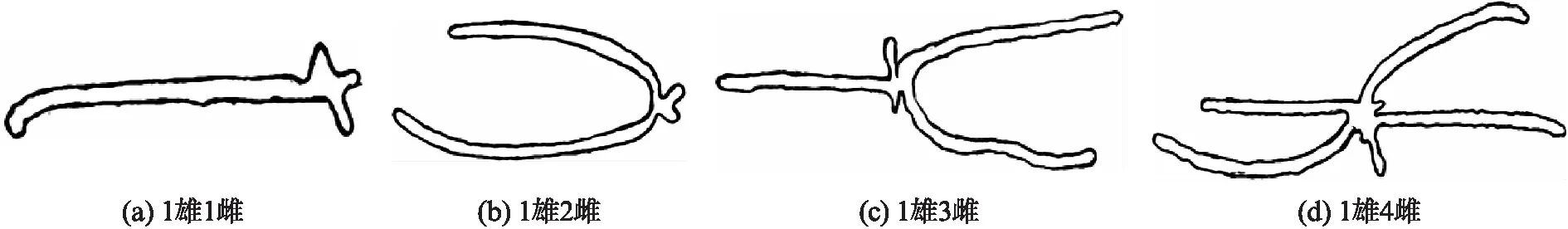
图3 具不同性比的母坑道形状类型
(3)子坑道形成及筑室化蛹
幼虫孵化后开始在树干的韧皮部与木质部之间取食并发展由细到粗的子坑道.初始子坑道方向与母坑道近垂直,随着幼虫成长,子坑道伸展方向不定,但相互间交叉较少(图4).子坑道在幼虫发育至高龄期后停止,长度范围为2.3~8.0cm(平均值=(4.9±0.3)cm).老熟幼虫化蛹前通常会在子坑道尽头挖一大小约4.8mm×1.8mm的椭圆形蛹室,并在其中化蛹.

图4 枫香刺小蠹子坑道形成末期
3 讨 论
准确识别昆虫的雌雄成虫的形态并了解繁殖特性是开展昆虫生物学研究和害虫防治的前提.一般而言,昆虫的雌雄成虫个体之间,除了在第一性征(如雄性输卵管,雌性产卵器等)具有明显差异外,在第二性征(如大小、颜色、触角等)上也会有不同的特征,尤其在昆虫中,雌雄二型现象普遍存在,但不同种类性征的表型差异不尽相同[15-19].本研究发现,枫香刺小蠹雌雄成虫的体长和前胸背板的最宽宽度无显著差异,这与大多数树皮小蠹的情况一致[20].在此基础上,本研究提供了基于外部形态特征区别枫香刺小蠹雌雄成虫的方法.该方法可以在不解剖外生殖器的基础上,快速准确地区分雌雄成虫,从而为调查工作中分别统计寄主树木上雌雄成虫的发生量提供了方便.
一雄多雌是树皮小蠹的重要交配繁殖模式,其雌虫会在各自形成的母坑道中产卵,但是关于每窝中母坑道数与雌成虫数对应关系的报道极少;作者在前期的实验中观察到,枫香刺小蠹的母坑道数和雌成虫数存在显著的对应关系,因此本研究将每窝中的母坑道数作为雌成虫数.本研究结果显示,枫香刺小蠹每窝雌雄性比在1∶1和2∶1的情况下,其单雌产卵量和卵的孵化率显著高于性比在3∶1和4∶1的情况,这一现象在果蝇、实蝇等其它种类昆虫的研究中亦有发现过[21-22].据推测,这可能与雄虫和多头雌虫交配后精子数量和质量下降有关,特别是对小蠹类昆虫来说,所产生的精子量有限,在与多头雌虫交配后,“精子不足”或“精子质量下降”的情况可能会发生.据此,我们认为,这种“一夫多妻”型的“婚配模式”,对枫香刺小蠹种群的持久发展可能产生不利影响.
树皮小蠹主要在韧皮部和木质部间蛀食危害,可留下具有种类特异性的为害痕迹——坑道系统,这是树皮小蠹为害的一个重要特征[23-28].一般来说,小蠹的种类鉴定主要依据形态特征、生化特征、细胞学特征和分子特征等参数,而这些参数的获得往往都需要借助解剖显微镜、特殊实验场所和设备等才能完成,在野外调查中难以快速应用[29-33].以往的研究表明,小蠹的坑道系统因虫而异,除少数种类“借用”其他虫种坑道繁殖外,很多都具有独特的坑道系统,据此特征在野外针对部分寄主植物和地点鉴定准确率可达70%左右,因此也是识别其种类的重要依据[34-35].枫香刺小蠹的坑道形成过程和其他树皮小蠹类似,但也有些许不同,这些坑道特点结合寄主树种等信息,可作为野外初步鉴定的重要依据.然而,值得注意的是,小蠹种类繁多,坑道特征亦千变万化,很多小蠹种类的坑道系统尚未报道,因此,枫香刺小蠹的坑道形态特征与物种本身是否具有独特的对应关系,该问题还需要进一步通过与其他小蠹坑道的对照研究.
枫香刺小蠹属于树皮小蠹,这类小蠹主要通过蛀食树木的疏导组织,对寄主造成严重为害甚至死亡[36].作者在前期野外调查研究中发现,枫香刺小蠹攻击致死的植株,最小胸径约3cm,小于该胸径的枫香树苗通常并不会遭受小蠹攻击,这可能是由于较小胸径植株韧皮部的营养量难以维持枫香刺小蠹正常生长的缘故.同时,作者也发现有胸径大于20cm的北美枫香被为害致死的现象,这一现象应引起重视,因为北美枫香引种到中国约20年,随着植株的生长,枫香刺小蠹对植株高大的北美枫香的潜在威胁并未降低.再者,随着北美枫香植株生长变高,实际工作中对枫香刺小蠹为害的监测和防治难度也会增大,因此,当前急需开发出一种有效监测和防控枫香刺小蠹的方法,控制该虫的发生蔓延.
猜你喜欢
食品安全导刊(2022年24期)2022-11-18
花卉(2022年8期)2022-05-09
今日农业(2021年6期)2021-11-27
环境与生活(2021年5期)2021-06-24
农家科技(2020年5期)2020-06-15
儿童故事画报·自然探秘(2019年10期)2019-01-14
现代农业研究(2017年10期)2018-01-02
当代贵州(2016年23期)2016-07-21
Coco薇(2015年12期)2015-12-10
现代畜牧科技(2015年12期)2015-10-21
