
有机物料对植烟土壤氮素矿化及微生物性质的影响*
2021-04-08 03:38李孝刚彭曙光靳志丽刘勇军周志高王兴祥
土壤学报 2021年1期
李孝刚,彭曙光,靳志丽,周 羽,刘勇军,周志高,王兴祥†
(1. 南方现代林业协同创新中心,南京林业大学生物与环境学院,南京 210037;2. 湖南省烟草科学研究所,长沙 410004;3. 中国科学院南京土壤研究所,南京 210008;4. 湖南省烟草公司永州市公司,湖南永州 425000)
土壤是优质烟系统工程的基础,是影响烤烟品质的首要环境因素。土壤有机质影响着土壤的其他理化性质以及微生物活性,是反映土壤肥力和地力的重要指标[1-2]。对于一般农作物而言,土壤有机质含量越高,土壤肥力性状越好,作物越容易获得高产稳产。但对于烤烟而言,由于其特殊的需氮规律,土壤有机质含量过高或过低对烤烟生长与品质均不利[3]。土壤有机氮有效化的主要途径是有机质的矿化和分解过程,矿化形成的无机氮是烟株氮营养的主要来源。其中,烤烟在整个生育期均表现出对硝态氮偏向吸收的特性,是烤烟氮素吸收的最主要形式,而铵态氮肥效相比硝态氮表现出一定的滞后性,且吸收积累量相对较少。因此,如何协调土壤的供氮特征,使之与烤烟生长需氮要求相匹配,有效改善植烟区优质烟叶的产率,成为烟草行业所面临的巨大挑战。
土壤氮素的供应与烤烟需氮规律的协调性是决定烤烟品质的关键因素[4]。一般认为土壤氮矿化量与土壤有机质含量呈显著正相关关系,土壤有机质含量过高并不利于烟叶品质的形成。原因在于,烟株生长后期土壤中依然能矿化出较多的无机氮,烟株贪青晚熟,烟叶不易正常落黄,吃味辛辣,可用性差[3]。然而,有机物料组成影响土壤有机氮矿化速率,其中碳氮比(C/N)是调控土壤有机氮矿化的重要因子[5-6]。目前,施用有机粪肥、饼肥、秸秆还田和种植绿肥等改善植烟土壤结构和养分供应的技术措施,在不同植烟区提升烟叶产质的生产实践效果往往不一,这可能与不同植烟区土壤性状存在显著差异有关[7]。土壤微生物是驱动土壤有机物质和养分转化的主要动力,外源有机物料的投入改变了土壤微生物生长所需能量物质的数量和质量,进而影响土壤微生物群落结构及活性[8],相应地会导致土壤养分供应尤其是氮素矿化特征的差异[9]。
湖南烤烟种植主要分布于湘南和湘西北两个生态区域,分别以烟-稻轮作及旱烟为主。本研究采集上述烟区典型植烟土壤,选择稻草、油菜、黑麦草及菜籽饼等C/N 不同的有机物料类型,开展室内培育试验,以揭示如下科学问题:(1)有机物料施入下高、低有机质土壤无机氮矿化特征和微生物功能活性响应差异,(2)不同C/N 有机物料施入对土壤无机氮矿化和微生物功能活性的影响,(3)有机物料施入下土壤氮素矿化的微生物机制。研究结果将为湖南烟区制定科学合理的有机物料施用制度、提高烤烟产质提供科学依据。
1 材料与方法
1.1 供试土壤及有机物料
分别在湖南省湘南植烟区和湘西植烟区选择典型烟田,湘南植烟土壤采自永州市蓝山县土市镇植烟水稻土(YZ,25°31′29.3″N,112°13′42″E),湘西植烟土壤采自湘西自治州花垣县道二乡植烟旱地(XX,28°31′25″N,109°27′28″E)。于2016 年5 月,按照多点取样法在选定的标准烟田采集0~20 cm土壤,带回实验室用于培育实验研究。将土壤中的石块以及植物残体(如根系)剔除后,风干过筛(10目)待用。取一部分研磨后过100 目筛孔,用于土壤理化性质测定(表1)。
供试有机物料选取成熟油菜秸秆(YC)、稻草(DC)、成熟黑麦草(HMC)和菜籽饼(CZB),为当地烟区常用的几种有机物料。各有机物料烘干、粉碎、碾磨,过40 目筛待用,其基本性质见表2。
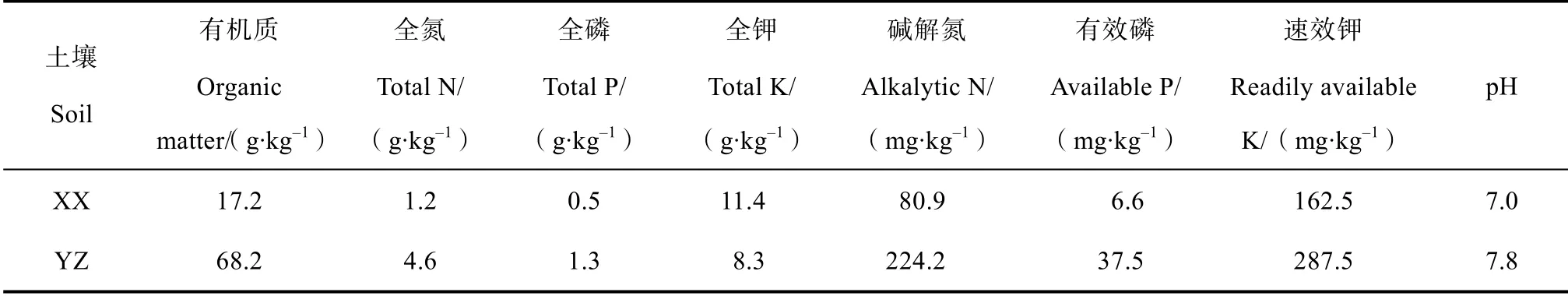
表1 供试土壤基本理化性质Table 1 Physico-chemical properties of the soils in the study

表2 各有机物料碳氮比Table 2 Carbon to nitrogen ratios of the organic materials tested
1.2 试验设计
试验设置稻草、油菜、菜籽饼、黑麦草以及对照(不添加物料,CK)5 个处理,每个处理3 次重复。各有机物料添加量均按纯 N 100 mg·kg-1土壤进行折算。称取50 g 风干土,不同处理中添加的有机物料与土壤充分混匀后,置于150 mL 三角瓶培养,用蒸馏水调节含水量至 60%田间最大持水量,于25 ℃恒温培养60 d,每隔3 d 采用恒重法补充蒸发的水分。培养过程中共设置180 个培养装置(三角瓶),用于在培养的第7、14、21、31、60 天开展破坏性采样,测定土壤铵态氮与硝态氮含量。在第14、31 天采样中,取部分样品用于测定土壤脲酶、蔗糖酶,并对31 d 采集的土壤样品分析土壤微生物群落功能多样性。
1.3 分析方法
有机物料总碳用重铬酸钾容量法测定,全氮用凯氏法测定。土壤样品的用 2 mol·L-1的KCl 浸提,靛酚蓝比色法测定;用1 mol·L-1的KCl 浸提,220 nm/275 nm 双波长紫外分光光度法[10]测定。土壤脲酶用靛酚蓝比色法测定,转化酶用3,5-二硝基水杨酸比色法[11]测定。
采用BIOLOG 生态板(Eco microplate,Matr ix Technologies Corporation,美国)分析法测定不同处理土壤微生物群落功能的多样性。主要步骤[12]为:称取相当于5 g 风干土壤的新鲜土壤样品加入盛有45 mL 灭菌的生理盐水(0.85%)三角瓶中,室温振荡30 min 后静置。吸取1 mL 土壤悬浮液至9 mL 无菌生理盐水中,混匀,用排枪将10-3稀释液均匀加至96 孔生态板,25℃恒温避光培养,0、24、48、72、96、120、144、168、192和 216 h 后在590 nm、750 nm 波长下测定吸光度值(OD)。
1.4 指标计算
不同有机物料处理下各时段土壤表观硝化速率=(各时段土壤N含量—初始土壤N含量)/培养天数。
对于BIOLOG 数据,培养72h的微生物生长代谢处于旺盛时期,因此选取培养时间72h的试验结果进行分析,利用平均颜色变化率(AWCD)来反映土壤微生物活性。计算公式为:AWCD=∑(C590-C750)/31,式中,C590、C750 分别为590、750 nm 的吸光值,31 为BIOLOG 生态板上碳源数量。
香农-威纳(Shannon-Wiener)多样性指数(H):式中,Pi为第i孔的相对吸光值与整个平板相对吸光值总和的比率。
1.5 数据处理方法
利用 Excel 2011 软件进行数据统计,用Origin 8.6 软件作图,采用SPSS 17.0 软件对各指标进行方差分析,用多重比较法(LSD)进行差异显著性检验,显著水平设为0.05。使用CANOCO 4. 5. 1 软件(Microcomputer Power,Ithaca,美国)对微生物功能多样性与主要碳源利用类型进行冗余分析,采用 499 次的蒙特卡罗排列检验(MonteCarlo permutation test,499 permutations,reduced model)进行显著性检验。
2 结 果
2.1 不同有机物料添加下植烟土壤无机氮矿化量的变化
对于高有机质植烟土壤,在整个培养过程中,添加油菜秸秆、菜籽饼和黑麦草处理下土壤硝态氮均呈现逐渐增加趋势(图1)。在培养前21 天,与对照相比,油菜秸秆、稻草和黑麦草处理均显著降低了高有机质土壤硝态氮含量,其中稻草处理降低了267.9%~675.9%;随着培养时期延续,油菜秸秆和黑麦草处理土壤硝态氮含量与对照差异逐渐不明显,而在整个培养阶段稻草处理土壤硝态氮含量均显著低于对照。但是,在整个培养时期菜籽饼处理增加了土壤硝态氮含量。而对低有机质土壤,土壤硝态氮呈现先上升后下降的趋势,与对照相比,不同有机物料添加均显著增加了土壤硝态氮含量。
在整个培养过程中,几种有机物料处理下高有机质土壤铵态氮含量呈逐渐下降趋势,而低有机质土壤呈现先上升后下降趋势。在培养第14 天,稻草和菜籽饼处理显著增加了高有机质土壤铵态氮含量,而其他不同时期各有机物料无显著性影响;但对于低有机质土壤,油菜秸秆和稻草秸秆添加显著降低了土壤铵态氮含量,而菜籽饼处理显著增加了土壤铵态氮含量,增幅达2.7 倍~3.3 倍。
2.2 不同有机物料添加下植烟土壤硝化速率及净矿化速率动态
对于高有机质土壤,除稻草处理外,其他各有机物料处理和对照的表观硝化速率在整个培养时期中均呈现先增加后下降趋势(图2a));而对于低有机质土壤,添加有机物料处理硝化速率呈现先增加后迅速下降的趋势,但对照土壤在整个培养过程中无明显变化(图2b))。与对照相比,添加C/N 较高的有机物料(稻草、油菜秸秆、黑麦草)显著降低了高有机质土壤表观硝化速率,而C/N 较低的菜籽饼处理显著增加了土壤表观硝化速率(图2a))。然而对于低有机质土壤,各有机物料添加均能显著增加土壤表观硝化速率(图2b))。
在整个培养时期中,油菜秸秆、菜籽饼和黑麦草处理土壤的净矿化速率呈先增加后下降的趋势,而在培养的前31 d 稻草处理下土壤净矿化速率呈负值,之后逐渐增加为正值(图2c))。与对照相比,油菜、稻草和黑麦草处理显著降低了高有机质土壤净矿化速率,而菜籽饼处理提高了土壤净矿化速率(图2d))。
2.3 不同有机物料添加下土壤酶活性变化
添加不同有机物料显著提升了高有机质土壤脲酶和转化酶活性(图3a),图3c))。稻草、油菜秸秆、菜籽饼和黑麦草处理下土壤脲酶活性分别较对照增加了100.1%、88.4%、32.5%和40.6%(图3a)),土壤转化酶活性分别增加了253.0%、57.2%、134.6%和127.5%(图3c))。在低有机质土壤,添加油菜秸秆和稻草可显著提升土壤脲酶活性,而菜籽饼处理显著降低了土壤脲酶活性(图3b))。相对于高有机质土壤,添加有机物料对低有机质土壤转化酶影响更明显,稻草、油菜秸秆和黑麦草处理下土壤转化酶活性分别为对照的27 倍、18 倍和34 倍(图3d))。四种有机物料类型之间,添加稻草处理的土壤脲酶和转化酶活性最高,添加菜籽饼处理最低。
2.4 不同有机物料添加下土壤微生物功能多样性变化
不同有机物料添加显著提升了植烟土壤微生物碳源代谢活性。在高有机质植烟土壤中,添加稻草、油菜秸秆和黑麦草处理土壤微生物活性分别为对照的2.8 倍、3.2 倍和3.6 倍(图4a)),低有机质植烟土壤下分别为对照的20.6 倍、12.9 倍和7.1 倍(图4b))。同样,稻草、油菜秸秆和黑麦草处理下土壤微生物功能多样性显著高于对照土壤,而菜籽饼添加对土壤微生物功能多样性无显著影响(图4c),图4d))。
通过对土壤微生物功能活性与主要利用碳源类型进行冗余分析,探讨不同有机物料对土壤微生物功能多样性起主要作用的因子。结果显示各有机物料处理下土壤微生物功能群落发生明显变异,其中,油菜秸秆和黑麦草处理较好地聚合在一起,而菜籽饼处理与其他有机物料处理出现明显分化。对于高有机质植烟土壤,利用氨基酸类、胺类等碳源的微生物种群的增多是造成油菜秸秆和黑麦草处理下微生物功能群落变异的主要因素,而利用酸类、糖类的微生物种群与稻草处理下微生物功能群落变异最为相关(图5a))。对于低有机质植烟土壤,利用糖类和脂类的微生物种群的增多是造成油菜秸秆处理下微生物功能群落变异的主要因素,而利用醇类和酸类的微生物种群是造成稻草处理下微生物功能群落变异的主要因素(图5b))。
3 讨 论
3.1 有机物料对土壤氮素矿化的影响
有机物料的特性决定了其施入土壤后的矿化行为[13-14]。高有机质含量土壤中添加高C/N 的物料,如稻草、油菜秸秆和黑麦草对土壤无机氮有固持作用,且这种效应与有机物料的C/N 比呈正相关性,表明土壤中不同有机物料处理间有机氮矿化过程与矿质氮的微生物同化过程的强弱取决于有机物料的C/N[15-16]。本研究中,稻草C/N 明显高于其他有机物料,而该处理下土壤呈现出负净矿化速率(图2),其原因在于丰富的碳源诱导了土壤微生物大量增殖,导致矿质氮的生物固持作用大于有机氮的矿化作用[17-18];而低C/N 的菜籽饼添加显著促进了土壤的净矿化速率,引起培养过程中土壤无机氮浓度显著高于对照。对于低有机质土壤,添加高C/N 的稻草、油菜秸秆和黑麦草虽然显著降低了土壤净矿化速率(图2),但提升了土壤硝态氮浓度(图1),这与有机物料添加下土壤硝化作用显著高于对照有关,进而导致了土壤铵态氮浓度显著降低[19-21]。同时,土壤氮的矿化是一个复杂的过程,受温度、水分、土壤pH 等多种因素的影响,因此,在等氮条件下不同有机物料添加量的差异引起土壤有机质、pH 和物理性质等其他理化性质的变化,这是否对土壤氮素矿化产生影响需要进一步研究[16,22]。
本研究发现,在高有机质土壤上添加高C/N 的稻草可引起土壤净矿化速率显著降低(图2)。然而,若烤烟种植在有机质含量过低的土壤,烟叶小而薄,烤后烟叶化学成分不协调,吃味平淡。本研究中在低有机质植烟土壤中添加菜籽饼可显著提升土壤的净矿化速率(图2),引起土壤中铵态氮积累和硝态氮浓度增加(图1)。由此可见,C/N 不同的有机物料添加下引起土壤完全不同的氮矿化特征:高有机质土壤中通过添加高C/N 的稻草可明显增加土壤矿质氮的固持,而低有机质土壤增施低C/N 的菜籽饼肥可有效补充土壤本身较低的矿化能力,进而提高土壤矿质氮含量。这在烟草农业生产中具有重要的现实指导意义。
值得注意的是,有机物料添加后土壤硝态氮和铵态氮浓度在高、低有机质土壤中表现差异较大,其中,在高有机质土壤中硝态氮浓度显著高于铵态氮,而在低有机质土壤中表现为铵态氮浓度显著高于硝态氮(图1)。这一差异说明高有机质土壤可能含有更高的硝化细菌数量,进而提高了土壤硝化效果[23]。也有研究发现长期施用有机肥的土壤中硝化细菌的群落组成发生了显著变化,硝化能力较强的种属如Nitrosospiracluster 3 在高有机质土壤中明显增多[24]。因此,低有机质土壤硝化能力较弱是导致铵态氮在土壤积累的重要原因。
3.2 有机物料对土壤微生物性质的影响
有机物料可以为土壤微生物增加额外营养源和能源,进而调控土壤微生物群落的结构和代谢功能[22]。然而,鉴于土壤微生物的响应差异,不同物料(碳源)可能诱导土壤中喜好该类碳源的微生物生长,从而对微生物群落代谢特征产生影响[25-26]。本研究表明,不管是高有机质还是低有机质土壤,添加有机物料后土壤微生物活性和功能多样性均显著上升(图4)。大量外源有机物料的加入增加了土壤微生物生长所需要的有效性碳源数量和种类,这种在能源供应方面的巨大扰动,极大地刺激了土壤异养微生物的活动,这也解释了有机物料添加下引起培养前期转化酶活性显著提升而后期无明显影响的结果(图3)。与此同时,对土壤微生物酶活性动态分析发现,高C/N 的有机物料添加显著提高了土壤脲酶的活性,而低C/N 的菜籽饼添加对土壤脲酶无显著影响(图3),这与不同有机物料的添加对土壤微生物功能活性影响的结果一致。Zhou 等[27]研究发现,添加外源物对低有机质土壤微生物群落多样性的影响大于高有机质土壤。可见,除所施用的有机物料种类的影响外,本底土壤有机质等也是影响微生物群落功能变异的重要因素。进一步对土壤微生物功能群落的冗余分析结果表明,不同类型有机物料诱导了土壤不同微生物功能群落的变异,其中,高有机质土壤中添加稻草诱导了利用醇类和酯类碳源的微生物类群增加,而油菜秸秆和黑麦草处理主要诱导了利用胺类碳源微生物类群增加(图5)。但是,无论是高有机质还是低有机质土壤,菜籽饼处理下土壤微生物群落对各种碳源利用能力并未明显增强(图5),这也解释了菜籽饼添加下高有机质土壤矿质氮含量无显著变化,而低有机质土壤较高的铵态氮浓度可能来自物料本身的分解,而不是来自土壤有机氮的矿化[15]。
4 结 论
有机物料对土壤氮矿化的影响因土壤有机质含量不同而异。施入高C/N 的有机物料(如稻草)可有效控制高有机质植烟土壤的矿化速率,显著降低土壤净矿化氮水平;而在低有机质植烟土壤施用高C/N 的有机物料将会加重土壤矿质氮素缺乏,施用菜籽饼肥可显著提升低有机质土壤中铵态氮水平,但这可能主要来自有机物料自身矿化。高C/N 的有机物料施入显著促进了土壤微生物功能多样性,引起土壤酶活性增高;但不同有机物料诱导了对碳源利用类型差异的微生物种群增多,造成土壤微生物功能群落发生变异。由此可见,在烟草生产实际中,应当根据植烟土壤性质的差异合理安排有机物料种类施入,充分发挥不同有机物料的效应,实现植烟土壤的可持续利用。
致谢感谢南京师范大学程谊教授在实验设计中的帮助和指导!
猜你喜欢
CHINA TODAY(2022年4期)2022-11-22
奥秘(创新大赛)(2022年8期)2022-09-14
水土保持学报(2022年3期)2022-05-26
科技创新与应用(2022年6期)2022-03-24
散文诗世界(2022年2期)2022-03-10
中国生态农业学报(中英文)(2022年1期)2022-01-15
中国果业信息(2021年10期)2021-12-07
花卉(2021年17期)2021-09-15
小猕猴学习画刊(2021年8期)2021-08-27
中国饲料(2021年8期)2021-05-17
